|
|
|
DISCUSSIONAnalysis of the Putative TrackmakersThe morphology and size range of the footprints and trackways analyzed herein strongly suggests that they were produced by a rodent or rodent-like mammal of medium to large size. Among all known South American taxa with tetradactyl manus and pes, the extinct rodent-like notoungulates (i.e., hegetotheriids and mesotheriids) can be ruled out. Hegetotheriids as Hegetotherium, Pachyrukhos and Hemihegetotherium have digit III longer than digits II and IV, and digit V is greatly reduced (Sinclair 1909). In addition, Pachyrukhos has arrow-like hoofed ungual phalanges that do not correspond with the long, heavy, and pointed claws observed in the Vinchina specimens. And, mesotheriids as Trachytherus and Mesotherium have a pentadactyl manus with a reduced digit V and the remaining four digits subequal in size (Shockey et al. 2007). The ungual phalanges are moderately flattened with the distal ends fissured (Shockey et al. 2007).
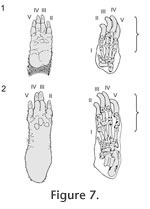
The only living species of Dinomyidae, Dinomys branickii, have lost external signs of the first digit in the forefoot and hind foot, being functionally tetradactyl (Figure 7) (Peters 1873;
Grand and Eisenberg 1982). The remaining skeletal elements of the pollex and hallux are found within a well-developed pad (Grand and Eisenberg 1982). Moreover, in Dinomys the third and fourth digits are almost equal in length and
longer than the second and fifth (Figure 7) (Grand and Eisenberg 1982), as in Tracheria troyana (Figure 3). A similar pattern is present in the Miocene extinct dinomyids of La Venta, Colombia as Olenopsis aequatorialis (sensu
Fields 1957, p. 337). Olenopsis, with longer and more robust digits than Dinomys, also would have functional tetradactyl manus and pes.
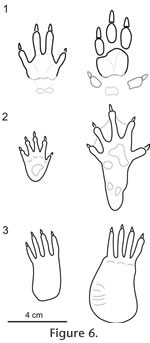
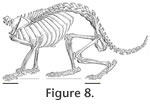
Grand and Eisenberg (1982) suggested that Dinomys has a long and narrow foot with a slightly mobile preaxial pad related with its predominately cursorial habit; opposite to the arboreal Coendou that has a short and broad foot with a more flexible, opposable pad for powerful grasping. The trackways of Tracheria troyana represent a digitigrade cursorial gait to occasionally plantigrade, as occurs in Dinomys (see Peters 1873, p. 233, pl. 3). The footprints represent the impression on the substrate of the digits and the metapodial-phalangeal junctions (Figure 7). That portion of the feet represents in Dinomys the half of the total length of the forefoot and a third of the total length of the hind foot (Figure 8). Based on the autopodial measurements provided by Peters (1873, pp. 233-234) (manus 80 mm long and 30 mm wide, pes 125 mm long and 32 mm wide), Dinomys would produce forefoot and hind foot footprints of similar length (approximately 4 cm long), although the hind foot is 36% longer. The same reasoning could be used to explain the sub-equal size (see Table 1) of the Vinchina forefoot and hind foot impressions.
Carrano and Wilson (2001) established that when analyzing footprint producers, it is preferable to establish osteological synapomorphies that could be identified in the fossil footprints. Unfortunately, until now, phylogenetic studies of caviomorph rodents (including dinomyids) do not include characters of the distal autopodium in their analyses (e.g., Adkins et al. 2001; Huchon and Douzery 2001; Spotorno et al. 2004; Horovitz et al. 2006; Nasif 2010). In that context, we analyzed putative trackmakers based on anatomical features of the postcranial skeleton that are transcribed to the footprint morphology and trackway pattern. Until new osteological evidence is provided by the finding of new specimens of caviomorph rodents with distal autopodium preserved, the most parsimonious hypothesis is that the footprints of Vinchina were produced by dinomyids or a close relative with a similar appendicular skeleton. The Dinomyid Bone-Teeth Record in South America
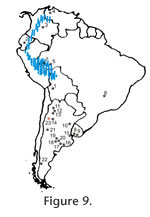
The oldest record of the family corresponds to Branisamys luribayensis Hoffstetter and Lavocat 1970, from the Late Oligocene of Bolivia (Hoffstetter and Lavocat 1970; Patterson and Wood 1982; Mones 1986; Vucetich 1986; Nasif 2010). During the Neogene, a few species are represented in various countries of South America though there is a wide record of dinomyids in Argentina. In Ecuador, dimonyids are preserved in sediments of possible Miocene age at the Nabon area (Fields 1957; Patterson and Wood 1982). Also, they are preserved in La Venta (Middle Miocene), Colombia (Fields 1957; Walton 1997); the Urumaco Formation, (Laventanense South American Land Mammal Age), Venezuela (Linares 2004); Rio Acre (Late Miocene) and Minas Gerais (Pliocene-Pleistocene), Brazil (Paula Couto 1951; Frailey 1986); and the Camacho (Late Miocene) and Raigón formations (Pliocene-Pleistocene), Uruguay (Mones and Castiglioni 1979; Rinderknecht and Blanco 2008). In Argentina there is a wide record of dinomyids, and they are known since the Middle Miocene by isolated teeth from the Laguna Blanca and Río Frías formations of Patagonia (Ameghino 1904). The highest diversity occurs during the Late Miocene and Pliocene in Central North Argentina. In the northeastern area they occur particularly in deposits known as the "conglomerado osífero" (McKenna and Bell 1997; Nasif et al. 2004; Candela 2005; Candela and Nasif 2006), in northwestern Argentina are recorded at the Chiquimil and Andalhuala formations (Herbst et al. 2000; Nasif 2006, 2010). And, in central Argentina they are recorded at the Salicas Formation, La Rioja province; the Ullum Formation, San Juan province and the Epecuén, Arroyo Chasicó, and other formations from Buenos Aires province (e.g., Bondesio et al. 1980; Contreras 1985; Candela 2005; Tauber 2005; Nasif 2010) (Figure 9).The occurrence of Tacheria troyana in Late Oligocene deposits of the Vinchina Formation could represent the oldest record of Dinomyidae in Argentina, suggesting the extension of its temporal and geographical distribution. It also would indicate that dinomyids were widely distributed in South America since the Late Oligocene. The Record of Caviomorph Rodent Footprints in South AmericaThe large footprints of Vinchina Tracheria troyana are not comparable to any other footprint recorded in South America. Indeed, the record of South American Hystricognathi rodent footprints is scarce, and it is mostly restricted to caviioid (i.e., hydrochoerids and caviids) footprints, recorded in Argentina. Angulo and Casamiquela (1982) proposed a new ichnotaxon Porcellusignum consulcator for digitigrade footprints with tridactyl and occasionally tetradactyl impressions recorded in the Río Negro Formation, at Balneario El Cóndor, Río Negro province, Argentina. The material was assigned with doubt to Hydrochoeridae, unfortunately the photographs of the footprints are not clarifying. Besides the material recorded by Angulo and Casamiquela (1982), Aramayo (2007) described additional specimens assigned to cf. Porcellusignum isp. preserved in the Río Negro Formation (Late Miocene-Early Pliocene), between the Balneario El Cóndor and La Lobería, Río Negro province, Argentina. The footprints were interpreted as produced by hydrochoerids and the material consists of tetradactyl manus and tridactyl pes impressions. In addition, heteropod footprints (tetradactyl manus and tridactyl pes) assigned to dolichotine caviids were described by Aramayo and M. de Bianco (2009) from Pleistocene deposits of Pehuen-Có, Buenos Aires province, Argentina. More recently, Krapovickas et al. (2009b) described heteropod footprints (tetradactyl manus and tridactyl pes) preserved in Miocene deposits of the Toro Negro Formation at Vinchina, La Rioja province, Argentina. The footprints were suggested to be produced by two separate groups: hegetotheriid notoungulates and caviomorph rodents (most likely dolichotine caviids). All the previously mentioned footprints, with tetradactyl manus and tridactyl pes, differ strongly from Tracheria troyana, not only on its digit arrangement but also in the general morphology and larger size of the footprint. Furthermore, some mammal footprints, described and illustrated by Leonardi (1994), were assigned with doubt to rodents and are recorded in the Neogene of Argentina, all of them are markedly dissimilar to Tracheria troyana. One of the records is preserved at Salar del Rincón, Salta province, possibly in the Trinchera Formation. Another is preserved at Catua in the boundary between Salta and Jujuy provinces. Finally, small galloping mammal footprints are recorded at the Quebrada del Jarillal Formation at Sierra de Mogna, San Juan province, Argentina. |
|