

 Pleistocene record of mammals and pollen from Mexico (Las Tazas, Valsequillo, Puebla) and their paleoenvironmental interpretation
Pleistocene record of mammals and pollen from Mexico (Las Tazas, Valsequillo, Puebla) and their paleoenvironmental interpretation
Article number: 27.1.a15
https://doi.org/10.26879/1285
Copyright Paleontological Society, February 2024
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 26 March 2023. Acceptance: 23 January 2024.
ABSTRACT
The Valsequillo paleontological area in Puebla has been widely explored and several fossil localities are studied with the presence of megafaunal and micromammal remains. It has been suggested the possible presence of humans by indirect evidence from Pleistocene outcrops. Despite being a site of great paleontological interest, the geology of the area is complex and poorly defined, mainly at a local scale. We present the new Late Pleistocene locality Las Tazas, Valsequillo, Puebla, Mexico, which is located in Barranca Caulapan, bearing megafaunal remains and fossil pollen material. Twenty-two megafaunal elements and 32 fossil pollen samples were recovered from the same stratigraphic layer. The collected fossil material was deposited in the Paleontological Collection of the Benemérita Universidad Autónoma de Puebla. The fossil mammal remains represent the orders Cingulata, Perissodactyla, Artiodactyla, and Proboscidea; the families Pampatheriidae, Equidae, Bovidae, Camelidae, and Elephantidae; the genera Pampatherium, Equus, Bison, Camelops, and Mammuthus; and the species Pampatherium mexicanum, Equus conversidens, Camelops hesternus, and Mammuthus columbi. The fossil pollen of the genera Pinus and Quercus is identified; as well as pollen related to grasses (e.g., Poaceae, Asteraceae, and Amaranthaceae). The presence of Bison allows us to infer a Late Pleistocene-Early Holocene age for the fossil association. Las Tazas is the fossil locality with the greatest paleodiversity in the state of Puebla. Palynological analyses indicate a paleoenvironment of grasslands with Pinus and Quercus open forests and the presence of water bodies.
Azarael Tomas-Mosso. Benemérita Universidad Autónoma de Puebla, Facultad de Ciencias Biológicas. Ciudad Universitaria, Puebla, Puebla, México. azaraeltomas@hotmail.com
Carlos Castañeda-Posadas. Benemérita Universidad Autónoma de Puebla, Laboratorio de Paleobiología, Ecocampus-BUAP, San Pedro Zacachimalpa, Puebla, Mexico. (Corresponding author) carlos.castaneda@correo.buap.mx
J. Alberto Cruz. La Brea Tar Pits and Museum, Los Angeles, USA. Centro de Investigación Quinametzin, Coordinación Nacional de Arqueología, Instituto Nacional de Antropología e Historia, Mexico City, Mexico. cruzsilvajac@yahoo.com.mx
Ivan Alarcón-Durán. Instituto Nacional de Antropología e Historia, Sección de Paleontología, Centro INAH Puebla, Av. Ejércitos de Oriente S/N, Centro Cívico Cinco de Mayo Los Fuertes, Cd. Puebla, Puebla, Mexico. ialarcond@yahoo.com.mx
Keywords: Quaternary; megafauna; paleoecology; vegetation change; Puebla-Tlaxcala Basin
Final citation: Tomas-Mosso, Azarael, Castañeda-Posadas, Carlos, Cruz, J. Alberto, and Alarcón-Durán, Ivan. 2024. Pleistocene record of mammals and pollen from Mexico (Las Tazas, Valsequillo, Puebla) and their paleoenvironmental interpretation. Palaeontologia Electronica, 27(1):a15.
https://doi.org/10.26879/1285
palaeo-electronica.org/content/2024/5141-fossils-from-las-tazas-valsequillo-puebla-mexico
Copyright: February 2024 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
The biological diversity in Mexico is mainly explained by the great climatic variation that prevails in the region, due to both its latitude and longitude location and its orography, which generate temperate and tropical conditions that converge in the central part of the country (Challenger and Soberón, 2008; Ferrari et al., 2012). The central part of Mexico shows a complex vegetation pattern resulting from its ecological and geological history (Mastretta-Yanes et al., 2015).
In Mexico, changes in the floristic and faunal composition during the Pleistocene are documented at high elevations (Trans-Mexican Volcanic Belt [TMVB], Sierra Madre Oriental, and Sierra Madre Occidental); they were generated by (cold) glacial and (warm) interglacial episodes at higher elevations (Arroyo-Cabrales et al., 2005). During these glacial and interglacial periods the dispersion of vertebrate fauna occurred (Ceballos et al., 2010; Ferrusquía-Villafranca et al., 2010), resulting in an extensive record of large mammals from studies carried out from the 1950s to the present day (Armenta, 1959; Pérez-Crespo et al., 2009; Ceballos et al., 2010; Ferrusquía-Villafranca et al., 2010; Castañeda Posadas, 2015; Castañeda-Posadas et al. 2022).
The state of Puebla is located in the TMVB and near Puebla city. The Valsequillo Basin has the highest number of Pleistocene mammals recorded in the state (Arroyo-Cabrales et al., 2005; Ferrusquía-Villafranca et al., 2010; Castañeda-Posadas, 2015; Herrera Flores, 2017). The Valsequillo area is an interesting locality because there is a record of the presence human at the end of the Pleistocene (Pichardo 1997; VanLandingham 2004; Huddart et al., 2008; González et al., 2006, 2015).
Despite the extensive megafaunal evidence in Mexico, the paleobotanical record is scarce. Metcalfe et al. (2016) reported the presence of Pleistocene pollen and diatoms from Valsequillo suggesting that the region was covered by lake or a water body surrounded by pine forest (Straka and Ohngemach, 1989; VanLandingham, 2006). Other paleoenvironmental reconstructions have been based on stable isotope analyses in megafaunal remains and continental snails (VanLandingham, 2006; Stevens et al., 2012; Pérez-Crespo et al., 2014; Metcalfe et al., 2016) suggesting the presence of grasslands and open forests. However, as is the case with paleontological studies of the Pleistocene of Mexico, fossil studies in vertebrates are independent of paleobotanical micropaleontological studies. Therefore, this study presents the record of the Pleistocene mammals from Las Tazas, Valsequillo Basin, Puebla, linking the information between the megafauna and fossil pollen record, and combining paleontological efforts to substantially complement the paleoenvironmental interpretation.
GEOLOGICAL SETTING
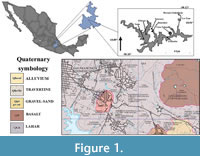 The locality Las Tazas, Junta Auxiliar de Santo Tomás Chautla, Puebla, is located in “18°57´06.3” N, “98°08´08.9” W, at 2076 m a.s.l. (Figure 1), southern Puebla City, the capital of the state.
The locality Las Tazas, Junta Auxiliar de Santo Tomás Chautla, Puebla, is located in “18°57´06.3” N, “98°08´08.9” W, at 2076 m a.s.l. (Figure 1), southern Puebla City, the capital of the state.
The oldest rocks in the area include an early Cretaceous limestone beneath sediments of the Balsas Group, mainly Eocene-Oligocene red conglomerates (Pichardo, 1997). The Pleistocene strata are composed of basaltic tuff and lava spills (named Zacachimalpa or Caulapan, lakes bed or Ixcalco lava by González et al., 2006). However, most of these sediments consist of volcanic deposits with sandy-carbonate clay and carbonated silt deposited in large basins to a lesser extent (Castañeda-Posadas, 2015). The upper strata are composed of igneous rocks (intermediate and acid tuffs, andesites, basalts, and volcaniclastics) and sedimentary rocks (limestones, sandstones-conglomerates, conglomerates, and siltstones-sandstones) of Pleistocene age (Jiménez-Hidalgo et al., 2015), where several mammal and human fossils have been found (González et al., 2006).
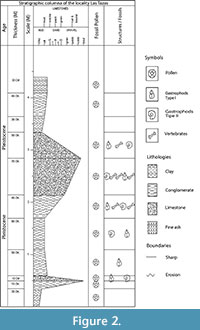 Las Tazas site is located on the western slope of Barranca Caulapan, where lacustrine, fluvial, and volcanic ash deposits known as Valsequillo gravels. The estimated age from Barranca Caulapan is between 38.9 and 7,151 14C years before present (González et al., 2006; Díaz-Sibaja et al., 2020). The stratigraphic column of Las Tazas includes, from the base to the top, a brown silt with clay, silts with conglomerate lens, brown silt with fine sands, conglomerate with presence of fossil mammals, green silts and ashes, and finally a black paleosol (Figure 2).
Las Tazas site is located on the western slope of Barranca Caulapan, where lacustrine, fluvial, and volcanic ash deposits known as Valsequillo gravels. The estimated age from Barranca Caulapan is between 38.9 and 7,151 14C years before present (González et al., 2006; Díaz-Sibaja et al., 2020). The stratigraphic column of Las Tazas includes, from the base to the top, a brown silt with clay, silts with conglomerate lens, brown silt with fine sands, conglomerate with presence of fossil mammals, green silts and ashes, and finally a black paleosol (Figure 2).
MATERIAL AND METHODS
A systematic excavation was conducted for the extraction of fossils (Viñas-Vallverdú et al., 2015), which were packed and transported in field jackets for their protection. The pollen samples from the sediments of the fossil-bearing strata and each sample was placed in hermetic plastic bags, which were individually labeled for their transportation. The stratigraphic column of Las Tazas was made in SedLog 3.1 (Zervas et al., 2009). The entire process of cleaning, stabilization, and reconstruction of specimens and the preparation of palynological material were done at the Paleontology Laboratory of the Benemérita Universidad Autónoma de Puebla (BUAP). We did the extraction of palynomorph following the procedure proposed by Martinez-Hernández, et al. (1980).
 A total of 22 megafaunal fossil elements were identified corresponding to four orders, five families, five genera, and six species (Table 1, Figure 3), as well as pollen records of the genera Pinus, and Quercus, and herbaceous (e.g., Poaceae, Asteraceae, and Amaranthaceae). Both megafaunal and pollen specimens were deposited in the Paleontology Laboratory rof BUAP (BUAPAL). Megafaunal fossils were identified using specialized literature (e.g., Skinner and Kaisen 1947; Schmid, 1972; Von den Driesch, 1976; Sisson and Grossman, 1978; Gillette and Ray, 1981; Dixon, 1993; Adams and Crabtree, 2001; Lopez Bejar et al., 2008; Campos-Camacho, 2012; Góis Lima, 2013) and compared to remains housed at BUAPAL and the Paleontological Collection of the Centro INAH-Puebla (CRINAHP). Palynomorphs were identified using specialized literature (Chavez and Rzedowski, 1993; Perveen, 1999; Van Wichelen et al., 1999; Flores-Cruz et al., 2006).
A total of 22 megafaunal fossil elements were identified corresponding to four orders, five families, five genera, and six species (Table 1, Figure 3), as well as pollen records of the genera Pinus, and Quercus, and herbaceous (e.g., Poaceae, Asteraceae, and Amaranthaceae). Both megafaunal and pollen specimens were deposited in the Paleontology Laboratory rof BUAP (BUAPAL). Megafaunal fossils were identified using specialized literature (e.g., Skinner and Kaisen 1947; Schmid, 1972; Von den Driesch, 1976; Sisson and Grossman, 1978; Gillette and Ray, 1981; Dixon, 1993; Adams and Crabtree, 2001; Lopez Bejar et al., 2008; Campos-Camacho, 2012; Góis Lima, 2013) and compared to remains housed at BUAPAL and the Paleontological Collection of the Centro INAH-Puebla (CRINAHP). Palynomorphs were identified using specialized literature (Chavez and Rzedowski, 1993; Perveen, 1999; Van Wichelen et al., 1999; Flores-Cruz et al., 2006).
SYSTEMATIC PALEONTOLOGY
Superorder XENARTHRA Cope, 1889
Order CINGULATA Illiger, 1811
Family PAMPATHERIIDAE Paula Couto, 1954
Genus PAMPATHERIUM Gervais and Ameghino, 1880
Pampatherium mexicanum Edmund, 1996
Referred material. One movable osteoderm (dermal bony plate) (BUAPAL 768; Figure 3A), movable osteoderm fragment (BUAPAL 767; Figure 3B), and two fixed scapular shields (BUAPAL 769 and 779; Figure 3C-D).
Description. The complete movable osteoderm (BUAPAL 768) has a rectangular shape with three sections (Montellano-Ballesteros, 1991). The lower part (45 mm) has a series of shallow foramina in the lower margin. The middle part (30 mm) is rugose and has a 35 mm long curvature at the attachment near the upper part; this latter part is completely smooth (Montellano-Ballesteros, 1991; Aguilar and Laurito, 2009; Góis Lima, 2013).
The movable osteoderm fragment (BUAPAL 768) only has the upper part (25 mm), with a rectangular shape, reticulations on the anterior articular surface, and a series of dots on the sculpted surface, mainly on the contour of the surface. The fixed osteoderms (BUAPAL 769 and 779) from the scapular region have a poorly marked ornamentation composed of multiple small pores, its marginal elevation is flattened or diffuse, and the longitudinal depressions are superficial.
Comparison and remarks. The family Pampatheriidae is represented in Mexico by two genera, Pampatherium and Holmesina. This family is characterized by diagnostic osteoderms that allow the distinction between the two genera (Edmund, 1996; Scillato-Yané et al., 2005). Pampatherium mexicanum is the only recognized species of this genus in North America (Mead et al., 2007). Movable osteoderms in the genus Pampatherium are poorly ornamented, the marginal band may be absent, particularly in the Mexican population, or may consist of a narrow band of follicular pores on the posterior part and the lateral margins; the marginal band is not significantly depressed under the submarginal band and in the central part is broad, low, and without protrusions or a raised keel (Edmund, 1996), which are characteristics observed in the fossil material from Las Tazas.
Order PERISSODACTYLA Owen, 1848
Family EQUIDAE Gray, 1821
Genus EQUUS Linnaeus, 1758
Equus conversidens Owen, 1869
Referred material. Left M2 (BUAPAL 773; Figure 3E), right M3 (BUAPAL 774; Figure 3F).
Description. The measurement of left M2 and right M3 are: length (Al) = 24.6 mm, transversal width (Tw) = 23.81 mm and Al =24.33 mm, Tw =19.31 mm. Both molars present the enamel well, and its occlusal configuration can be observed, where the lingual edge of the protocone is moderately concave, the enamel folding on the outer edge of the fossettes is simple and deep, the caballing fold is simple, faint in upper left M2 and strongly developed in upper right M3.
Comparison and remarks. The structures observed in the fossils BUAPAL 774 and 773 as observed in E. conversidens; (Reynoso-Rosales and Montellano-Ballesteros, 1994; Sánchez Salinas et al., 2016; Priego-Vargas et al., 2017). The values presented in this work are within the variation of E. conversidens and E. excelsus (Table 2), this species are smaller in size than E. mexicanus but are larger than E. cedralencsis and E. francisi. However, there are qualitative differences that help to separate both species, such as the lion edge of the protocone, which is more concave in E. excelsus than in E. conversidens; the folding of the enamel on the external edge of the pliprotoloph and plihypostyle pits are absent in E. excelsus and present in E. conversidens. The variation in size and occlusal characteristics of molars in equids has allowed the identification of at least five species associated in three groups of equids for the Mexican territory (Reynoso-Rosales and Montellano-Ballesteros, 1994; Sánchez-Salinas et al., 2016; Priego-Vargas et al., 2017).
Equus sp.
Referred material. Left mandible with three complete deciduous premolars dp2, dp3, and dp4 (BUAPAL 780; Figure 3I-J), right p3 (BUAPAL 771; Figure 3G), and left m2 (BUAPAL 772; Figure 3H).
Description. The left hemimandible has three deciduous premolars that are slightly worn. The length of the dp2 to dp4 is approximately 100 mm. The molariform p3 and m2 have an Al = 24.0 mm, Tw = 13.04 mm, Al = 22.4 mm, and Tw = 14.47 mm, respectively.
Comparison and remarks. The values of the dp2 to dp4 is within the variation of length of p2-p4 of adults E. mexicanus p2-p4 length = (86.99-109.95 mm), instead the other species of Pleistocene adult horses the maximum value of p2-p4 length is 92.6 mm (Priego-Vargas et al., 2017) (Table 3). Jiménez-Hidalgo et al. (2019) describe a jaw of a juvenile E. mexicanus with deciduous premolars with a length of the dp2 to dp4 =120 mm from Chiapas, Mexico. The juvenile specimen from Chiapas is larger than BUAPAL 780 and because the teeth of juvenile individuals do not present diagnostic characters to separate between species of horses, we determine the fossil material to genus level.
The molariforms are smaller than E. mexicanus, and its measurements are within the ranges of E conversidens and E. cedralensis. The molariforms exhibit advanced deterioration on the occlusal surface, but the prefosette and postfosette are visible. In the right p3 the u-shaped linguaflex, possibly pointed metastyle, entoconid possibly square and almost absent equine fold. The left m2 presents a v-shaped linguaflex, apparently round entoconid and # aballing fold. Several qualitative occlusal characters do not fall into the description reported for small horses in the literature (Sánchez-Salinas et al., 2016; Priego-Vargas et al., 2017), therefore we determine the fossil material to genus level.
Order ARTIODACTYLA Owen, 1848
Family BOVIDAE Gray, 1821
Genus BISON Hamilton Smith, 1827
Bison sp.
 Referred material. Cranium fragment with right horn core and lacking facial bones (BUAPAL 755, incomplete right mandible with a fragment of p3 and complete m1 to m3 (BUAPAL 756), complete molar M2 (BUAPAL 781), proximal scapula fragment (BUAPAL 761), a fragment of the proximal epiphysis of the right ulna (BUAPAL 754), an almost complete left metacarpus (BUAPAL 764), right talus (BUAPAL 763), fragment of cervical vertebra (BUAPAL 762), and two lumbar vertebrae (BUAPAL 758 and 759); Figure 4.
Referred material. Cranium fragment with right horn core and lacking facial bones (BUAPAL 755, incomplete right mandible with a fragment of p3 and complete m1 to m3 (BUAPAL 756), complete molar M2 (BUAPAL 781), proximal scapula fragment (BUAPAL 761), a fragment of the proximal epiphysis of the right ulna (BUAPAL 754), an almost complete left metacarpus (BUAPAL 764), right talus (BUAPAL 763), fragment of cervical vertebra (BUAPAL 762), and two lumbar vertebrae (BUAPAL 758 and 759); Figure 4.
Description. The cranium fragment (BUAPAL 755) retains part of the left and right frontal from the orbital fossa to the occipital condyle. The right horn core has a length of 200 mm, an internal length of 230 mm, an external length of 295 mm from tip to base, a circumference of 290 mm, a diameter of 100 mm, and an estimated total horn core length from tip to tip (TLTT) of 700 mm.
Comparison and remarks. These horn measurements and the horn cores laterally directed are within the variation of B. antiquus (Table 4) (Lucas, 1899; Skinner and Kaisen, 1947; Von den Driesch, 1976; McDonald, 1981; Díaz-Sibaja et al., 2020). B. occidentalis despite presenting horn nuclei that moved backwards (Díaz -Sibaja et al., 2020) is considered synonymous of B. antiquus based on ancient DNA studies that demonstrated that both are the same anagenetic lineage (Shapiro et al., 2004; Wilson et al., 2008).
The M2 (BUAPAL 781) has a length = 33.41 mm and a width = 22.06 mm. The occlusal surface is badly damaged, it presents a well-developed entostyle, the fossettes have the characteristic U shape with some folds, and the wear of the molar indicates it belonged to an adult. The incomplete right mandible presents a fragment of pm3 and complete m1, m2, and m3 (BUAPAL 756); the molars are hypsodont with well-developed entostyle, and the shape of the molars are sharp in lateral view with light wear. All of these traits are characteristics of genus Bison (Allen, 1876; Chandler, 1916; Hillson, 2005).
A fragmented right scapula (BUAPAL 761) presents the glenoid fossa with a diameter = 88 mm, the internal border of the scapula has a length = 165 mm, and the total length of the scapula is 430 mm. It is observed that the area immediately dorsal to the glenoid and bordered by the mid-neck ridge and posterior margin is markedly thinned and flattened, which is characteristic of the genus Bison (Von den Driesch, 1976; McCuaig-Balkwill and Cumbaa, 1992; France, 2008; Raymond and Prothero, 2011). The scapula belongs to an adult individual according to the measurements reported by Von den Driesch (1976), McCuaig-Balkwill and Cumbaa (1992), and France (2008).
The ulna is fragmented (BUAPAL 754), it presents a fracture in the diaphysis, the proximal region is preserved with a flattened and well-defined olecranon, with the major cavity in semi-lunate shape. The curvature of the olecranon has a depth of 14 mm. All these features and the ulna not fused with the radius indicate that the specimen was a juvenile (Ritz and Wing, 2008).
Left metacarpus (BUAPAL 764) is wider and slightly shorter, characteristic observed in Bison (McDonald, 1981; Skinner and Kaisen, 1947) and its measurements (Table 5) are close to those of B. antiquus (McDonald, 1981; McCuaig-Balkwill and Cumbaa, 1992). The articular condyle of the left metacarpal IV (BUAPAL 764) lacks distal epiphysis and it is close to B. antiquus (McDonald, 1981; Baskin, et al., 2022). However, we do not have diagnostic characteristics to assign it to specific level.
The right calcaneus (BUAPAL 763) shows the lateral articular surface of the distal trochlea covering the distal surface, the tubercle is at the same line with the dorsal portion, features of the genus Bison (McCuaig-Balkwill and Cumbaa, 1992).
The sixth cervical vertebra (BUAPAL 762) has well-defined caudal and cranial regions, the rostral condyle is convex with a rounded margin, the medullary canal is present; the articular process is wide with a rounded margin, the cranial articular process is flat and incomplete, the transverse processes are flat, large, and present a triangular shape, the articular processes are damaged, and the angle between the lateral branches of the transverse process is wider and deeper, similar to features present in Bison (McCuaig-Balkwill and Cumbaa, 1992).
The lumbar vertebrae (BUAPAL 758, 759) are almost complete. They exhibit a compression of the vertebral body in the cranial and caudal regions. The cranial region has a height = 52 mm, which is the same as the width of the facies = 52 mm. In the caudal region, the vertebrae present a width in the facies = 59 mm and the length of the vertebrae = 73 mm; the dorsal margin of the anterior articular process is at the same level than the upper part of the articular facet, characteristics observed in Bison (McCuaig-Balkwill and Cumbaa, 1992). The fossil material was determined to belong to an adult individual because of the similarity with the measurements reported by Von den Driesch (1976) and Skinner and Kaisen (1947).
All postcranial remains are assigned to Bison sp. because they exhibit the morphological characteristics of this genus (McDonald, 1981) but is very difficult separate species into this genus with postcranial material. Ancient DNA analyzes with 405 remains of Bison aged between 60,000 and recent, demonstrated broad genetic diversity in populations during the Late Pleistocene that are mainly subdivided into four major clades, showing that all Bison sequences are closely related and are not indicative of multiple and independent species (Wilson et al., 2008). Due to the similarity of the isolated horn nuclei and postcranial elements of B. antiquus and B. occidentalis, identification of Late Pleistocene and early Holocene Bison should be made with caution (Wilson et al., 2008).
Family CAMELIDAE Gray, 1821
Genus CAMELOPS Leidy, 1854
Camelops cf. C. hesternus Leidy, 1873
Referred material. Left calcaneus (BUAPAL 757), Figure 5.
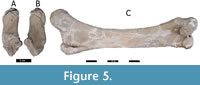 Description. The left calcaneus (BUAPAL 757) is robust, with a length = 158 mm, body width = 68 mm, interior curvature = 35 mm, and belongs to an adult individual since the epiphyses are well fused without a trace of the sutures (Reitz and Wing 2008).
Description. The left calcaneus (BUAPAL 757) is robust, with a length = 158 mm, body width = 68 mm, interior curvature = 35 mm, and belongs to an adult individual since the epiphyses are well fused without a trace of the sutures (Reitz and Wing 2008).
Comparison and remarks. Unlike other artiodactyls of similar size, such as the genus Bison, the cuboid articular facet is wider, the distal astragalar surface is divided into a medially oriented distal part and an anteriorly oriented proximal part, the articular surface of the fibula is convex proximally and concave distally, and the proximally positioned groove is shallower than in Bison (Guérin and Faure, 1999; Zazula et al., 2016). The family Camelidae in Valsequillo during the Late Pleistocene is represented by Camelops, Hemiauchenia, and Paleolama. We can exclude an assignment of BUAPAL 757 to Hemiauchenia because this genus is smaller (Table 6). Camelops and Paleolama are of similar size, but Camelops is more robust which is more consistent with the morphology of BUAPAL 757 (Labarca and López, 2006; Zazula et al., 2016) (Table 6).
Order PROBOSCIDEA Illiger, 1811
Family ELEPHANTIDAE Gray, 1821
Genus MAMMUTHUS Brookes, 1828
Mammuthus columbi Falconer, 1858
Referred material. Left femur (BUAPAL 775), Figure 5.
Description. The femur presents the greater and lesser trochanters well defined, the internal and external femoral condyles are also present, as well as the fossette in lesser trochanter region. It has a long body with a length =1300 mm, with well-defined proximal and distal regions with a width = 230 mm and 280 mm, respectively, and a slender diaphysis.
Comparison and remarks. Therefore, the characters mentioned above correspond to the genus Mammuthus, since the other two proboscidean families (Mammutidae and Gomphotheriidae) have expanded distal condyles medial and laterally and the robustness of the femur is less than in mastodon but is greater than gomphotheres, particularly on the shaft and medial condyle, and the femur of these families is smaller in size (Tapia-Ramírez et al., 2013). M. columbi is the only valid mammoth species of the Late Pleistocene in Mexico (Lister and Sher, 2015). M. columbi is the largest proboscidean in Mexico, it has a wide distribution in North America with more than 270 localities in central Mexico (Agenbroad, 2005; Arroyo-Cabrales et al., 2010).
PALYNOLOGICAL ANALYSIS
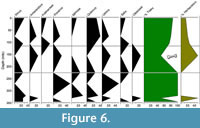 Ten samples of sediment were taken from the different layers in the study site (Figure 2), recovering a total of 38 palynomorphs. The dominant groups identified belong to genera Pinus (with 48%) and Quercus (with 52 %). However, the genera Amaranthus, Fraxinus, Mimosa, Ulmus, and Salix also appear, as well as representatives of the family Olaceae and Asteraceae (Figure 6).
Ten samples of sediment were taken from the different layers in the study site (Figure 2), recovering a total of 38 palynomorphs. The dominant groups identified belong to genera Pinus (with 48%) and Quercus (with 52 %). However, the genera Amaranthus, Fraxinus, Mimosa, Ulmus, and Salix also appear, as well as representatives of the family Olaceae and Asteraceae (Figure 6).
The pollen diagram shows variation along the stratigraphic column in the dominance of floristic elements and in the arboreal and herbaceous elements (Figure 6). The basal layers of the sequence show a greater presence of the genera Pinus and Quercus, suggesting the presence of other tree groups and a temperate forest. Subsequently, in the sequence from 115 to 230 cm deep, where the bone-bearing stratum is found, the arboreal groups are getting scarcer, and the herbaceous groups and plants associated with water bodies dominate. In the stratum above 115 cm, the dominance of a closed forest reappears, and the portion of creeping plants, and plants associated with water bodies decreases.
The results suggest the variation In the conditions and floristic associations, in the basal is a temperate forest (of Pinus and Quercus), but later it becomes more grassland (Amarantaceae and Asteraceae) and presence of water bodies as suggested by the presence of the genera Fraxinus, Ulmus, and Mimosa.
DISCUSSION
Interdisciplinary studies of the fossil record are fundamental to analyze the possible changes in the flora and fauna throughout geological time, the response of biotic communities to climate change, and extinction events (O’Keefe et al., 2023). Specific information about the extinctions, diversification processes, and paleodistribution during the Pleistocene can be obtained from Mexico (Ferrusquía-Villafranca et al., 2017) due that ⁓800 localities with presence of fossil mammals have been recorded across the Mexican territory (Ferrusquía-Villafranca et al., 2010). Valsequillo Basin is a fossiliferous area with the most representative Late Pleistocene fauna of the state of Puebla (Jiménez-Hidalgo et al., 2015), and various studies have been conducted in this place (González et al., 2006). The continuous discovery of fossil material in this locality allows a better understanding of both plants and animals and the different structures of the Pleistocene communities, as well as contributing information that can be compared with paleobiological patterns present in other regions (Jiménez-Hidalgo et al., 2015).
The locality Las Tazas in the Valsequillo area is characterized by its distinct diversity of Pleistocene mammals, including artiodactyls (Bison sp. and Camelops hesternus), proboscideans (Mammuthus), equids (Equus conversidens, Equus sp.), and pampatheriids (Pampatherium mexicanus), which correspond to a Rancholabrean association, consistent with previous reports (Ceballos et al., 2010; Ferrusquía-Villafranca et al., 2010, 2017). In addition, the inclusion of the plant species emphasizes the heterogeneity of the vegetation present in the surroundings of this locality. In the megafauna-bearing stratum, the identified pollen allows inferring a vegetation composed of a semi-open mixed forest composed mainly of pines and oaks (Lozano-García et al., 2014) and the genus Salix indicates the presence of water bodies in the region (García-Ortega and Belmonte Soler, 2008). Open-type vegetation with a water body nearest to site in low areas and more closed plant communities in elevated areas, were available for herbivores with mixed diets, thus showing heterogeneity in vegetation and species derived fauna associated with food diversity (Louys et al., 2011). For example, the presence of grasslands, Amaranthus, and Asteraceae is associated with the faunal record of bisons, mammoths, and horses; on the other hand, shrub areas with presence of Pinus, Fraxinus, Mimosa, Quercus, Ulmus, and Salix can be associated with camels (Green et al., 2005; Pérez-Crespo et al., 2009; Stevens et al., 2012).
There are few studies documenting the plant paleodiversity of the state of Puebla, and it was until the end of the 1980s when Straka and Ohngemach (1989) conducted a palynological analysis in the basin of the Mexican highland, considered the first palynological record for Puebla, reporting the presence of Alnus, Carpinus, Carya, Junglans, Ulmus, Fraxinus, Liquidambar, and Fagus, which is confirmed with that recorded by Stevens et al. (2012). The latter authors mention that, at approximately 20 000 YBP, the climate of Valsequillo was colder and may have been drier compared to present-day conditions, and thus the species would have had to adapt to the changing climatic conditions. Metcalfe et al. (2016) report the presence of pines and woody shrubs, as well as shallow lakes, according to the pollen found, which allows making an association with the present pine-oak vegetation where the megafauna inhabited. This association of forest patches and increased water availability is supported by records of angiosperm wood such as Cordia, Dalbergia, and Prioria (Sainz-Reséndiz, 2011).
Therefore, the study of palynological material allows to infer patterns about the population dynamics of the vegetation present in the locality Las Tazas in an effective way as along as presence data are related to similar localities (Ferrusquía-Villafranca et al., 2010). The importance of this locality with respect to the association of the flora and fauna present, compared to the localities in central Mexico, lies in the diversity of species with the same specific ecological requirements, which suggests that the organisms belonged to the same community (Herrera Flores, 2017).
Many studies referring to the area of Valsequillo show the great diversity of species that have been identified over more than half a century (Arroyo-Cabrales et al., 2005; Armenta, 1959; Cruz-Muñoz, 2009; Ferrusquia-Villafranca et al., 2010; Castañeda- Posadas, 2015; Bravo-Cuevas et al., 2016; Herrera Flores, 2017), but they were initially carried out without a systematic collection of information and are mainly based on material collected in the 1960s that lacks stratigraphic data. In contrast, in the present study, we were able to obtain as much information as possible on the stratigraphy and both plant and animal diversity of the Late Pleistocene in a specific area of Valsequillo, Puebla.
CONCLUSIONS
During the Late Pleistocene, the megafauna dominated the landscape of the locality Las Tazas and was part of the Pleistocene ecosystems of Valsequillo, with a great diversity of organisms.
The Pleistocene landscape of Las Tazas was dominated by herbivores associated with the vegetation structure, which was composed of trees (Pinus and Quercus) and an herbaceous stratum (Oleaceae, Mimosa, Asteraceae [Compositae]).
The landscape inferred for the locality Las Tazas a tree assemblage composed of a temperate-humid forest with open areas and bodies of water. The megafauna that inhabited the landscape of Las Tazas was characteristic of Pleistocene ecosystems dominated by large herbivores, grazers and browsers.
ACKNOWLEDGEMENTS
The authors thank the Vice-Rectory for Research and Postgraduate Studies VIEP-BUAP, for the support for the development of their research. Also to the Consejo de Paleontología of the INAH, for the endorsement of the Project official No. 401.1S.3-2020/131 and 401.1S.3-2022/265 to CCP-IAD-JACS.
REFERENCES
Adams, B. and Crabtree, P. 2011. Comparative Osteology a Laboratory and Field Guide of Common North American Animals. Academic Press, Kidlington, Oxford, UK.
http://booksite.academicpress.com/9780123884374
Agenbroad, L. 2005. North American proboscideans: Mammoths: The state of knowledge, 2003. Quaternary International, 126–128:73–92.
https://doi.org/10.1016/j.quaint.2004.04.016
Aguilar, D.H. and Laurito, C.A. 2009. El armadillo gigante (Mammalia, Xenarthra, Pampatheriidae) del río Tomayate, Blancano tardío-Irvingtoniano temprano, El Salvador, América Central. Revista Geológica de América Central, 41:25–36.
https://doi.org/10.15517/rgac.v0i41.4176
Allen, J.A. 1876. The American bison. Memoirs of the Geological Survey of Kentucky, 1:1–248.
Armenta, J. 1959. Hallazgo de un artefacto asociado con mamut, en el Valle de Puebla. Instituto Poblano de Antropología e Historia e Instituto Nacional de Antropología e Historia, México.
Arroyo-Cabrales, J., Polaco O.J., and Johnson, E. 2005. La mastofauna del Cuaternario tardío en México. Bases de datos SNIB-CONABIO, proyecto G012. Instituto Nacional de Antropología e Historia, México.
https://www.snib.mx/iptconabio/resource?r=SNIB-G012
Arroyo-Cabrales, J., Polaco, O.J., Johnson, E., and Ferrusquia-Villafranca, I. 2010. A perspective on mammalian biodiversity and zoogeography in the Late Pleistocene of México. Quaternary International, 212:187–197.
https://doi.org/10.1016/j.quaint.2009.05.012
Baskin, J.A., Walls, R., and Thomas, R.G. 2022. Bison antiquus and B. latifrons from The Latest Rancholabrean (Latest Pleistocene), Nueces River Valley, South Texas, p. 213–224. In Morgan G.S. (ed.), Late Cenozoic Vertebrate Paleontology: Tribute to Arthur H. Harris. New Mexico Museum of Natural History and Science, Albuquerque, New Mexico, USA.
Bravo-Cuevas, V.M., Arroyo-Cabrales, J., and Priego-Vargas, J. 2016. The record of camelids (Artiodactyla, Camelidae) from the Valsequillo Basin, Late Pleistocene of Puebla state, central Mexico: taxonomy, diet, and geographic distribution. Revista Brasileira de Paleontología, 19:243–258.
https://doi.org/10.4072/rbp.2016.2.08
Brookes, J. 1828. A Catalogue of the Anatomical and Zoological Museum of Joshua Brookes, Esq. F.R.S. F.L.S. & c. Duplicate.
Campos-Camacho, M.A. 2012. Los équidos del Rancholabreano (Pleistoceno tardío) de la Fauna Local Viko vijin, Mixteca Alta Oaxaqueña, sur de México. Tesis de Licenciatura. Centro Universitario de Ciencias Biológicas y Agropecuarias. Universidad de Guadalajara, México.
Castañeda-Posadas, C. 2015. El registro paleobiológico del estado de Puebla. Benemérita Universidad Autónoma de Puebla, Puebla, México.
Castañeda-Posadas, C., Pérez-Lara, D.K., and Cruz, J.A. 2022. El registro paleontológico del estado de Tlaxcala. Benemérita Universidad Autónoma de Puebla, Puebla, México.
Ceballos, G., Arroyo-Cabrales, J., and Ponce, E. 2010. Effects of Pleistocene environmental changes on the distribution and community structure of the mammalian fauna of Mexico. Quaternary Research, 73:464–473.
https://doi.org/10.1016/j.yqres.2010.02.006
Challenger, A. and Soberón, J. 2008. Los ecosistemas terrestres, p. 87–108. In Soberon, J., Halffter, G., and Llorente-Bouquets, J. (eds.), Capital natural de México. Vol. I: Conocimiento actual de la biodiversidad. CONABIO, Ciudad de Mexico, México.
Chandler, A.C. 1916. A study of the skull and dentition of Bison antiquus Leidy, with special reference to material from the Pacific Coast. Bulletin of the Department of Geology, 9:121–135.
Chávez, R.P. and Rzedowski, J. 1993. Estudio palinológico de las floras fósiles del Mioceno Inferior y principios del Mioceno de la región de Pichucalco, Chiapas, México. Acta Botánica Mexicana, 24:1–96.
Cope, E.D. 1889. The Edentata of North America. The American Naturalist, 23:657–664.
Cruz-Muñoz, V., Arroyo-Cabrales, J., and Graham, R.W. 2009. Rodents and lagomorphs (Mammalia) from the Late-Pleistocene deposits at Valsequillo, Puebla, México. Current Research in the Pleistocene, 26:147–149.
Díaz-Sibaja, R., Jiménez-Hidalgo E., Ponce-Saavedra J., and García-Zepeda, M.L. 2020. A combined mesowear analysis of Mexican Bison antiquus shows a generalist diet with geographical variation, Journal of Paleontology, 92:1130–1139.
https://doi.org/10.1017/jpa.2018.19
Dixon, P.M. 1993. The radiological appearance of mandibular cheek teeth in ponies of different ages. Equine Veterinary Education, 5:317–323.
https://doi.org/10.1111/j.2042-3292.1993.tb01061.x
Edmund, A.F. 1996. A review of the Pleistocene giant armadillos (Mammalia, Xenarthra, Pampatheriidae). In Stewart, K.M. and Seymour, K.L. (eds.), Palaeontology and Palaeoenvironments of Late Cenozoic Mammals: Tributes to the Career of C.S. (Rufus) Churcher. University of Toronto Press, Toronto.
https://doi.org/10.3138/9781487574154-016
Falconer, H. 1858. On the species of mastodon and elephant occurring in the fossil state in England. Part II. Elephas. Quarterly Journal of the Geological Society, 14:81–84.
https://doi.org/10.1144/GSL.JGS.1858.014.01-02.11
Ferrari, L., Orosco-Esquivel, T., Manea, V., and Manea, M. 2012. The dynamic history of the Trans-Mexican Volcanic Belt and the Mexico subduction zone. Tecnophysics, 522–253:122–149.
https://doi.org/10.1016/j.tecto.2011.09.018
Ferrusquía-Villafranca, I., Arroyo-Cabrales, J., Martinez-Hernández, E., Gama-Castro, J., Ruiz-Gonzales, J., Polaco, O.J., and Johnson, E. 2010. Pleistocene mammals of Mexico: A critical review of regional chronofaunas, climate change response and biogeographic provinciality. Quaternary International, 217:53–104.
https://doi.org/10.1016/j.quaint.2009.11.036
Ferrusquía-Villafranca, I., Arroyo-Cabrales, J., Johnson, E., Ruiz-González, J., Martínez-Hernández, E., Gama-Castro, J., de Anda-Hurtado, P., and Polaco, O.J. 2017. Quaternary mammals, people, and climate change: A view from southern North America, p. 27–67. In Monks, G. (ed.), Climate Change and Human Responses: A Zooarchaeological Perspective. Springer, The Netherlands.
Flores-Cruz, M., Santana-Lira, H.D., Martínez-Bernal, A., and Fraile, M. 2006. Morfología del Polen de Mimosa Serie Quadrivales (Leminosae, Mimosoideae). Acta Botanica Mexicana, 77:1–13.
France, D.L. 2008. Human and Nonhuman Bone Identification: A Color Atlas. CRC Press, USA.
García-Ortega, P. and Belmonte Soler, J. 2008. Pinos (Pinus) Características botánicas y distribución. Unidad de Botánica, 2:139–142.
Gervais, H. and Ameghino, F. 1880. Los mamíferos fósiles de América del Sur. F. Savy-Igon Hnos, Paris–Buenos Aires.
Gillette, D.D. and Ray, C.E. 1981. Glyptodonts of North America. Smithsonian Institution Washington, D.C.
https://doi.org/10.5479/si.00810266.40.1
Góis Lima, F. 2013. Análisis morfológico y afinidades de los Pampatheriidae (Mammalia, Xenarthra). Unpublished PhD Thesis, Universidad Nacional de La Plata, La Plata, Argentina.
González, S., Huddart, D., and Bennett, M. 2006. Valsequillo Pleistocene archaeology and dating: Ongoing controversy in Central Mexico. World Archaeology, 38:611–627.
https://doi.org/10.1080/00438240600963155
González, S., Huddart, D., Israde-Alcántara, I., Domínguez-Vázquez, G., Bischoff, J., and Felstead, N. 2015. Paleoindian sites from the Basin of Mexico: Evidence from stratigraphy, tephrochronology and dating. Quaternary International, 363:4–19.
https://doi.org/10.1016/j.quaint.2014.03.015
Gray, J.E. 1821. On the natural arrangement of vertebrate animals. London Medical Repository and Review, 15:296–310.
Green, J.L., Semprebon, G.M., and Solounias, N. 2005. Reconstructing the paleodiet of Florida Mammut americanum via low-magnification stereomicroscopy. Palaeogeography, Palaeoclimatology, Palaeoecology, 223:34–48.
https://doi.org/10.1016/j.palaeo.2005.03.026
Guérin, C. and Faure, M. 1999. Palaeolama (Hemiauchenia) niedae nov. sp., nouveau Camelidae du Nordeste brésilien et sa place parmi les Lamini d'Amérique du Sud. Geobios, 32:629–659.
Hamilton-Smith, C. 1827. Order 7, Ruminantia, p. 296-376. In Cuvier, G. (ed.), A synopsis of the Class Mammalia, The Animal Kingdom. Vol. 5. Whittaker, London, UK.
Herrera Flores, J.A. 2017. Investigaciones paleontológicas en el Valle de Puebla durante el siglo XX. Ciencia Ergo Sum, 23:259–266.
https://doi.org/10.30878/ces.v24n3a8
Hillson, S. 2005. Teeth. Cambridge University Press, Cambridge, UK.
Huddart, D., Bennett, M.R., González, S., and Velay, X. 2008. Analysis and preservation of Pleistocene human and animal footprints: An example from Toluquilla, Valsequillo basin (Central Mexico). Ichnos, 15:232–245.
https://doi.org/10.1080/10420940802470599
Illiger, C. 1811. Prodromus Systematis Mammalium et Avium Additis Terminis Zoographicis Utriusque Classis. C. Salfeld, Berlin.
Jiménez-Hidalgo, E., Carbot-Chanona, G., and Castañeda-Posadas, C. 2015. El registro de mamíferos fósiles de Puebla, p. 173–186. In Castañeda-Posadas, C. (ed.), El Registro Paleobiológico de Estado de Puebla. Benemérita Universidad Autónoma de Puebla, Puebla, México.
Jiménez-Hidalgo, E., Carbot-Chanona, G., Guerrero-Arenas, R., Bravo-Cuevas, V.M., Holdridge, G.S., and Israde-Alcántara, I. 2019. Species diversity and paleoecology of Late Pleistocene horses from southern Mexico. Frontiers in Ecology and Evolution, 7:394.
https://doi.org/10.3389/fevo.2019.00394
Labarca, R.O. and López, P.G. 2006. Los mamíferos finipleistocénicos de la Formación Quebrada Quereo (IV Región-Chile): biogeografía, bioestratigrafía e inferencias paleoambientales. Mastozoología neotropical, 13:89–101.
Leidy, J. 1854. Description of a fossil apparently indicating an extinct species of the camel tribe. Proceedings of the Academy of Natural Sciences of Philadelphia, 7:172–173.
Leidy, J. 1858. Notice of remains of extinct Vertebrata from the valley of the Niobrara River, collected during the exploring expedition of 1857, in Nebraska. Proceedings of the Academy of Natural Sciences of Philadelphia, 10:20–29.
Leidy, J. 1873. Contributions to the extinct vertebrate fauna of the western territories. United States geological and geographical survey of the territories, 1:14–358.
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. 1: Regnum animale. Editio decima, reformata. Laurentii Salvii, Stockholm, Sweden.
Lister, A.M. and Sher, A.V. 2015. Evolution and dispersal of mammoths across the Northern Hemisphere. Science, 350:805–809.
https://doi.org/10.1126/science.aac5660
Lopez Bejar, M.A.L., López-Plana, C., Mayor-Aparicio, P., Navarro-Beltrán, M., Carretero-Romay, A., Nacher-García, V., and Ruberte-París, J. 2008. Atlas de osteología de los mamíferos domésticos. Universitat Autònoma de Barcelona, Spain.
http://veterinariavirtual.uab.es/anatomia/osteologia/Atlas_Virtual/primera.html
Louys, J., Meloro, C., Elton, S., Ditchfield, P., and Bishop, L.C. 2011. Mammal community structure correlates with arboreal heterogeneity in faunally and geographically diverse habitats: implications for community convergence. Global Ecology and Biogeography, 20:717–729.
https://doi.org/10.1111/j.1466-8238.2010.00643.x
Lozano-García S., Correa-Metrio A., and Luna L. 2014. Análisis de la lluvia de polen moderna de la cuenca de México: una herramienta para la interpretación del registro fósil. Boletín de la Sociedad Geológica Mexicana, 66:1–10.
https://doi.org/10.18268/bsgm2014v66n1a1
Lucas, F.A. 1899. The fossil bison of North America. Proceedings of the United States National Museum, 21:755–771.
Martínez-Hernández, E., Ludlow-Wieches, B., and Sánchez-López, M. 1980. Palinología y sus aplicaciones geológicas: cuenca carbonífera de Fuentes-Rio Escondido, Coahuila. Comisión Federal de Electricidad, Ciudad de México, Mexico.
Mastretta-Yanes, A., Moreno-Letelier, A., Piñero, A., Jorgense, T.H., and Emerson, B.C. 2015. Biodiversity in the Mexican highlands and the interaction of geology geography and climate within the Tras-Mexican Volcanic Belt. Journal of Biogeography 42:1586–1600.
https://doi.org/10.1111/jbi.12546
McCuaig-Balkwill, D. and Cumbaa, S.L.1992. A guide to the identification of postcranial bones in Bos taurus and Bison bison. Syllogeus No. 71. Canadian Museum of Nature, Ottawa.
McDonald, J.N. 1981. North American Bison. University of California Press, Berkeley, USA.
Mead, J.I., Swift, S.L., White, R.S., McDonald, H.G., and Baez, A. 2007. Late Pleistocene (Rancholabrean) Glyptodont and Pampathere (Xenarthra, Cingulata) from Sonora, Mexico. Revista Mexicana de Ciencias Geológicas, 24:439–449.
Metcalfe, S.E., Leng, M.J., Kirby, J.R., Huddart, D., Vane, C.H., and González, S. 2016. Early-Mid Pleistocene environments in the Valsequillo Basin, Central Mexico: a reassessment. Journal of Quaternary Science, 31:325–336.
https://doi.org/10.1002/jqs.2851
Montellano-Ballesteros, M. 1991. Panorama de las faunas pleistocénicas de vertebrados de México, en convención sobre la evolución geológica de México. Universidad Autónoma del Estado de Pachuca, México.
O’Keefe, F.R., Dunn, R.E., Weitzel, E.M., Waters, M.R., Martínez, L.N., Binder, W.J., Southon, J.R., Cohen, J.E., Meachen, J.A., DeSantis, L.R.G., Kirby, M.E., Ghezzo, E., Coltrain, J.B., Fuller, B.T., Farrell, A.B., Takeuchi, G.T., McDonald, G., Davis, E.B., and Lindsey, E.L. 2023. Pre-Younger Dryas megafaunal extirpation at Rancho La Brea linked to fire-driven state shift. Science, 381:eabo3594.
https://doi.org/10.1126/science.abo3594
Owen, R. 1848. Description of teeth and portions of jaws of two extinct Anthracotheroid quadrupeds (Hyopotamus vectianus and Hyopbovinus) discovered by the Marchioness of Hastings in the Eocene deposits on the N.W. coast of the Isle of Wight: with an attempt to develop Cuvier’s idea of the classification of pachyderms by the number of their toes. Quarterly Journal of the Geological Society of London, 4:103–141.
Owen, R. 1869. On fossil remains of Equines from Central and South America referable to Equus conversidens, Ow., Equus tau, Ow., and Equus arcidens, Ow. Philosophical Transactions of the Royal Society of London, 159:559–573.
Paula Couto, C. 1954. Sôbre um gliptodonte do Uruguay e um tatu fóssil do Brasil. Divisão de Geologia e Mineralogía, Notas preliminares e Estudios, 80:1–10.
Peerven, A. 1999. Contributions to the pollen morphology of the farnily Compositae. Turkish Journal of Biology, 23:523–535.
Pérez-Crespo, V.A., Sánchez-Chillón, B., Arroyo-Cabrales, J., Albertdi, M.T., Polaco, O.J., Santos-Moreno, A., and Cienfuegos-Alvarado, E. 2009. La dieta y el hábitat del Mamut y los caballos del Pleistoceno Tardío de El Cedral con base en isótopos estables (13C, 18O). Revista Mexicana de Ciencias Geológicas, 26:347–355.
Pérez-Crespo, V.A., Arroyo-Cabrales, J., Alva-Valdivia, L.M., Morales-Puente, P., Cienfuegos-Alvarado, E., Otero, F.J., and Ochoa-Castillo, P. 2014. La paleodieta de cinco especies de mamíferos herbívoros rancholabreanos de Valsequillo (Puebla, México). Revista Chilena de Antropología, 30:76–82.
Pichardo, M. 1997. Valsequillo biostratigraphy: new evidence for Pre-Clovis date. Anthropologischer Anzeiger, 55:233–246.
Priego-Vargas, J., Bravo-Cuevas, V.M., and Jiménez-Hidalgo, E. 2017. Revisión taxonómica de los équidos del Pleistoceno de México con base en la morfología dental. Revista Brasileira de Paleontología, 20:239–268.
https://doi.org/10.4072/rbp.2017.2.07
Raymond, K.R. and Prothero, D.R. 2011. Did climate changes affect size in Late Pleistocene Bison? New Mexico Museum of Natural History and Science Bulletin, 53:636–640.
Reitz, E.J. and Wing, E.S. 1999. Zooarchaeology. Cambridge University Press, Cambridge, UK.
Reynoso-Rosales, V.H and Montellano-Ballesteros, M. 1994. Revisión de los équidos de la fauna Cedazo del Pleistoceno de Aguascalientes, México. Revista Mexicana de Ciencias Geológicas, 11:87–105.
Sainz-Reséndiz, B.A. 2011. Descripción e identificación de maderas del Paleógeno de San Juan Atzingo, Puebla, México. Tesis de licenciatura. Facultad de Estudios Superiores, Iztacala. Universidad Nacional Autónoma de México, México.
Sánchez Salinas, M., Jiménez Hidalgo, E., and Castañeda Posadas, C. 2016. Mamíferos fósiles del Pleistoceno tardío (Rancholabreano) de San Mateo Huexoyucán, Tlaxcala, México. Boletín de la Sociedad Geológica Mexicana, 68:497–514.
Schmid, E. 1972. Atlas of Animal Bones: For Prehistorians, Archaeologists and Quaternary Geologists. Elsevier, Amsterdam, The Netherlands.
Scillato-Yané, G.J., Carlini, A.A., Tonni, E.P., and Noriega, J.I. 2005. Paleobiogeography of the late Pleistocene pampatheres of South America. Journal of South American Earth Sciences, 20:131–138.
https://doi.org/10.1016/j.jsames.2005.06.012
Servicio Geológico Mexicano. 2014. Carta Geológico Minero: San Francisco Totimehuacán E14-B53, Puebla. Servicio Geológico Mexicano, Pachuca Hidalgo, Mexico.
Shapiro, B., Drummond, A.J., Rambaut, A., Wilson, M.C., Matheus, P.E., Sher, A.V., Pybus, O.G., Gilbert, M.T.P., Barnes, I., Binladen, J., Willerslev, E., Hansen, A.J., Baryshnikov, G.F., Burns, J.A., Davydov, S., Driver, J.C., Froese, D.G., Harington, C.R., Keddie, G., Kosintsev, P., Kunz, M.L., Martin, L.D., Stephenson, R.O., Storer, J., Tedford, R., Zimov, S., and Cooper, A. 2004. Rise and Fall of the Beringian Steppe Bison. Science, 306:1561–1565.
https://doi.org/10.1126/science.1101074
Sisson, J.D. and Grossman, R.G. 1978. Anatomía de los animales domésticos (Vol. 1). Salvat Editores, Madrid, España.
Skinner, M.F. and Kaisen, O.C. 1947. The fossil Bison of Alaska and preliminary revision of the genus. Bulletin of the American Museum of Natural History, 89:123–256.
Stevens, R.E., Metcalfe, S.E., Leng, M.J., Lamb, A.L., Sloane, H.J., Naranjo, E., and González, S. 2012. Reconstruction of late Pleistocene climate in the Valsequillo Basin (Central Mexico) through isotopic analysis of terrestrial and freshwater snails. Palaeogeography, Palaeoclimatology, Palaeoecology, 319:16–27.
https://doi.org/10.1016/j.palaeo.2011.12.012
Straka, H. and Ohngemach, D. 1989. Late Quaternary vegetation history of the Mexican highland, p. 115–132. In Ehrendorfer, F. (ed.), Woody plants–evolution and distribution since the Tertiary. Springer, Vienna, Austria.
Tapia-Ramírez, G., Guzmán, A.F., and Polaco, O.J. 2013. Los gonfoterios (Proboscidea, Gomphotheriidae) de Colima, México. Boletín de la Sociedad Geológica Mexicana, 65:591–607.
VanLandingham, S.L. 2004. Corroboration of Sangamonian age of artifacts from the Valsequillo region, Puebla, Mexico by means of diatom biostratigraphy. Micropaleontology, 50:313–342.
https://doi.org/10.2113/50.4.313
VanLandingham, S.L. 2006. Diatom evidence for autochthonous artifact deposition in the Valsequillo region, Puebla, Mexico during the Sangamonian (sensu lato= 80,000 to ca. 220,000 yr BP and Illinoian (220,000 to 430,000 yr BP). Journal of Paleolimnology, 36:101–116.
https://doi.org/10.1007/s10933-006-0008-4
Van Wichelen, J., Camelbeke, K., Chaerle, P., Goetghebeur, P., and Huysmans, S. 1999. Comparison of different treatments for LM and SEM studies and systematic value of pollen grains in Cyperaceae. Grana, 38:50–58.
https://doi.org/10.1080/001731300750044708
Viñas-Vallverdú, R., Arroyo-Cabrales, J., Rivera-González, II, Rodríguez-Álvarez, X. P., Rubio-Mora, A., Eudave-Eusebio, I.N., and Ardelean, C.F. 2015. Recent archaeo-palaeontological findings from Barranca del Muerto site, Santiago Chazumba, Oaxaca, México. Quaternary International, 431:168–180.
https://doi.org/10.1016/j.quaint.2015.04.055
Von den Driesch, A. 1976. A guide to the measurement of animal bones from archaeological sites. Peabody Museum of Archaeology and Ethnology, Harvard University, Massachusetts, USA.
Wilson, M.C., Hills, L.V., and Shapiro, B. 2008. Late Pleistocene northward-dispersing Bison antiquus from the Bighill Creek Formation, Gallelli gravel pit, Alberta, Canada, and the fate of Bison occidentalis. Canadian Journal of Earth Sciences, 45:827–859.
https://doi.org/10.1139/E08-027
Zazula, G.D., MacPhee, R.D., Hall, E., and Hewitson, S. 2016. Osteological assessment of Pleistocene Camelops hesternus (Camelidae: Camelinae: Camelini) from Alaska and Yukon. American Museum Novitates, 2016:1–45.
https://doi.org/10.1016/S0016-6995(95)80217-7
Zervas, D., Nichols, G.J., Hall, R., Smyth, H.R., Lüthje, C., and Murtagh, F. 2009. SedLog: A shareware program for drawing graphic logs and log data manipulation. Computers & Geosciences, 35:2151–2159.
https://doi.org/10.1016/j.cageo.2009.02.009
