RESULTS AND DISCUSSION
 We performed CWT and SA analysis on fish-scale remains in 66 sample horizons in core TUL99B-03. Ten species of fish were documented, with Pacific
Herring and Northern Anchovy being the most abundant. We required 3-10 cm of core per sample to obtain sufficient fish-scale material. For this sample size, the effective Nyquist sampling interval
(Davis
1986) obtainable for CWT and SA was 17 cm, giving a ~60 year temporal resolution
(Figure 8). Thus, only cycles with periods greater than 60 years could be considered reliable. As indicated by
Figure 10 and Figure 11 70- to 110-year cycle, corresponding to the well-documented Gleissberg Cycle (Garcia and Mouradian 1998), is a predominant influence on both
Northern Anchovy and Pacific Herring populations throughout the sampled intervals. Cycles attributable to the Suess cycle are also present in the fish-scale record
(Figure 10, Figure 11). The 200-year Suess Cycle
(Sonnett and Finney
1990) has been linked to the Maunder Minimum of the Little Ice Age, and corresponds to other Maunder-like minima with the same 200-year period in Mid-Holocene sediments from North America
(Dean et al.
1996) and elsewhere (Dean
2000). The ca. 3,400 BP phase/frequency shift (Figure
8) is also evident in both the herring and anchovy data
(Figure 11). Palynological climate proxy records from sites around British Columbia record widespread and regionally variable climate change about that time, which correspond to the development of neoglacial conditions in the Cascadia region
(Pellatt et al.
2001). Conditions on the west coast of Vancouver Island changed from warmer and dryer than at present to cooler and wetter than at present. A 120- to180-year cycle, in the northern anchovy data, between 1,800 and 3,000 yBP, appears to have been a temporary signal, while the 680-year cycle could be the anomalous result of three nearby samples with very high anchovy scale populations.
We performed CWT and SA analysis on fish-scale remains in 66 sample horizons in core TUL99B-03. Ten species of fish were documented, with Pacific
Herring and Northern Anchovy being the most abundant. We required 3-10 cm of core per sample to obtain sufficient fish-scale material. For this sample size, the effective Nyquist sampling interval
(Davis
1986) obtainable for CWT and SA was 17 cm, giving a ~60 year temporal resolution
(Figure 8). Thus, only cycles with periods greater than 60 years could be considered reliable. As indicated by
Figure 10 and Figure 11 70- to 110-year cycle, corresponding to the well-documented Gleissberg Cycle (Garcia and Mouradian 1998), is a predominant influence on both
Northern Anchovy and Pacific Herring populations throughout the sampled intervals. Cycles attributable to the Suess cycle are also present in the fish-scale record
(Figure 10, Figure 11). The 200-year Suess Cycle
(Sonnett and Finney
1990) has been linked to the Maunder Minimum of the Little Ice Age, and corresponds to other Maunder-like minima with the same 200-year period in Mid-Holocene sediments from North America
(Dean et al.
1996) and elsewhere (Dean
2000). The ca. 3,400 BP phase/frequency shift (Figure
8) is also evident in both the herring and anchovy data
(Figure 11). Palynological climate proxy records from sites around British Columbia record widespread and regionally variable climate change about that time, which correspond to the development of neoglacial conditions in the Cascadia region
(Pellatt et al.
2001). Conditions on the west coast of Vancouver Island changed from warmer and dryer than at present to cooler and wetter than at present. A 120- to180-year cycle, in the northern anchovy data, between 1,800 and 3,000 yBP, appears to have been a temporary signal, while the 680-year cycle could be the anomalous result of three nearby samples with very high anchovy scale populations.
 Analysis of the data using SA shows that there is also a positive cross-correlation at 0.0125 cpy (80-year period) between herring remains and the brighter diatom rich layers that were deposited under cold upwelling conditions at the Gleissberg Cycle frequency
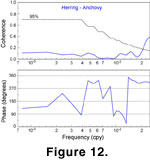
(Figure 11). Additional SA analysis (Figure
12) indicates that herring and anchovy have neither a positive nor negative correlation with each other at Gleissberg cycle frequencies. Squared coherency analysis also indicates that peaks in cross-amplitude spectrum and negative (-3 radians) correlation between herring and anchovy at the Gleissberg cycle band (70- to 90-year wavelength,
Figure 12) is not significant at the 95% confidence level calculated after
Emery and Thomson
(2001). There may be a correlation between herring and anchovy populations at higher frequencies (e.g., at time-scales of years to interdecadal scales;
Chavez et al.
2003) but detection was not possible at the sampling resolution available here.
Both species are negatively correlated with a 1,300-year cycle that we associate with the Bond cycle.
Analysis of the data using SA shows that there is also a positive cross-correlation at 0.0125 cpy (80-year period) between herring remains and the brighter diatom rich layers that were deposited under cold upwelling conditions at the Gleissberg Cycle frequency
(Figure 11). Additional SA analysis (Figure
12) indicates that herring and anchovy have neither a positive nor negative correlation with each other at Gleissberg cycle frequencies. Squared coherency analysis also indicates that peaks in cross-amplitude spectrum and negative (-3 radians) correlation between herring and anchovy at the Gleissberg cycle band (70- to 90-year wavelength,
Figure 12) is not significant at the 95% confidence level calculated after
Emery and Thomson
(2001). There may be a correlation between herring and anchovy populations at higher frequencies (e.g., at time-scales of years to interdecadal scales;
Chavez et al.
2003) but detection was not possible at the sampling resolution available here.
Both species are negatively correlated with a 1,300-year cycle that we associate with the Bond cycle.
 The herring data is also positively correlated to the gray values for cycle periods of 210 and 300 years, whereas the anchovy are negatively correlated to gray values at 340-year periods. Our findings indicate particularly close relationships between the Gleissberg solar cycle and the cyclicity of both fish species examined here, particularly Pacific
Herring populations, and by inference ocean productivity. This relationship holds through most of the 1,800-4,700 years BP record even through a major climate shift at 3,400 years BP that disrupted most other cycles
(Figure 11). Our findings compare favorably with the 100-year rate of anchovy fluctuation in Santa Barbara Basin between A.D. 270-1970
(Baumgartner et al.
1992) that falls within the variability of the Gleissberg Cycle (Garcia and Mouradian
1998). Because the anchovy populations of the North American CUD are linked, there is an indication that these pelagic fish species may have been fluctuating according to the Gleissberg Cycle through at least the last 4,700 years.
The herring data is also positively correlated to the gray values for cycle periods of 210 and 300 years, whereas the anchovy are negatively correlated to gray values at 340-year periods. Our findings indicate particularly close relationships between the Gleissberg solar cycle and the cyclicity of both fish species examined here, particularly Pacific
Herring populations, and by inference ocean productivity. This relationship holds through most of the 1,800-4,700 years BP record even through a major climate shift at 3,400 years BP that disrupted most other cycles
(Figure 11). Our findings compare favorably with the 100-year rate of anchovy fluctuation in Santa Barbara Basin between A.D. 270-1970
(Baumgartner et al.
1992) that falls within the variability of the Gleissberg Cycle (Garcia and Mouradian
1998). Because the anchovy populations of the North American CUD are linked, there is an indication that these pelagic fish species may have been fluctuating according to the Gleissberg Cycle through at least the last 4,700 years.
Correlation between solar activity and climate change has been suggested at a variety of different scales. Satellite-based measurements do show that there is a clear correlation between solar irradiance and the 11-year Schwabe cycle. Although the irradiance amplitude through the Schwabe cycle is only ~ 0.1%
(Beer et al.
2000) this energy variability may be sufficient to produce changes in the surface climate as a result of atmospheric dynamic responses to changes in temperature and stratospheric ozone
(Schindell et al. 1999;
Haigh
2001) or perhaps due to changes in cloudiness caused by variations in the intensity of galactic cosmic rays in the atmosphere
(Carslaw et al. 2002;
Shaviv and Veizer
2003). There is also growing evidence that solar forcing, at longer time-scales, may be an important driving forces of the climate system
(Soon et al. 2000; Beer et al. 2000) and dependent ecosystems at several temporal scales through the late Holocene
(Haigh
2001; Bond et al. 2001;
Schindell et al.
2001).
These examples of possible mechanisms for solar-driven climate change, and the teleconnection mechanisms proposed to propagate them globally, provide independent support for our hypothesis that solar forcing at the Gleissberg frequency seems to be an important control on the cycling of Pacific
Herring and Northern Anchovy. However, solar forcing need not be the only mechanism involved. The observation that the solar irradiance driven Gleissberg cycle is unaffected by a major climate shift that disrupted several other cycles is in itself suggestive that there are at least two independent climate forcing mechanisms at play.
Our correlation analysis of the fish data indicates that the ecosystem in Effingham Inlet is varying according to the climatic changes recorded in the sediments, regardless of the ultimate source of the climate forcing. Population fluctuations such as those indicated in our data are inevitable as these ecosystem shifts affect the carrying capacity of the environment
(Francis et al. 1999).
