| |
RESULTS
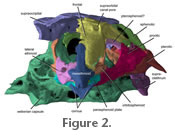 The specimen of Sarcoglanis simplex analyzed for this pilot study was only 17 mm standard length. The head skeleton is 5.5 mm long and 5 mm wide across the widest part of the skull (from left to right opercle) giving the head skeleton an almost spherical overall shape. The delicate jaw, suspensory, branchial, and pectoral girdle elements are moveably (kinetically) articulated to a pear-shaped neurocranium (Figure 2). The specimen of Sarcoglanis simplex analyzed for this pilot study was only 17 mm standard length. The head skeleton is 5.5 mm long and 5 mm wide across the widest part of the skull (from left to right opercle) giving the head skeleton an almost spherical overall shape. The delicate jaw, suspensory, branchial, and pectoral girdle elements are moveably (kinetically) articulated to a pear-shaped neurocranium (Figure 2).
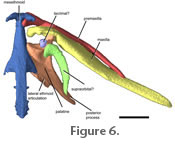
Neurocranium. The mesethmoid is a long bone located along the midline of the snout (Figure 2,
Figure 3,
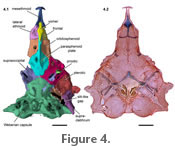
Figure 4,
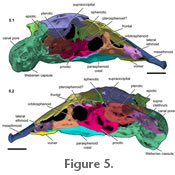
Figure 5, and
Figure 6).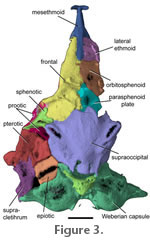 It is slender anteriorly, and its posterior third widens at its joint with the lateral ethmoids. The cornua of the mesethmoid are curved anteroposteriorly creating a v-shaped notch on the midline (Figure
2). Lateral ethmoids are situated posterolaterally ventrally to the mesethmoid, ventral to the frontal-mesethmoid joint and anterior to the orbitosphenoid (Figure 2). The lateral ethmoids are laterally expanded and cuplike. Lateral ethmoids are opened dorsally but covered by the anterior process of the frontal. The lateral ethmoids are much wider than the frontal process of the mesethmoid (Figure 3 and
Figure 4). Posterior to the lateral ethmoids is the orbitosphenoid that contributes to the floor and anterior sidewalls of the neurocranium. The floor of the orbitosphenoid is relatively thin (Figure 3 and
Figure 4). There is a sharp notch posteriorly on the orbitosphenoid where the parasphenoid plate attaches. The neck of the parasphenoid extends anteroventrally to the orbitosphenoid and contacts the posteriormost portion of the lateral ethmoids and median vomer (Figure 4). It is slender anteriorly, and its posterior third widens at its joint with the lateral ethmoids. The cornua of the mesethmoid are curved anteroposteriorly creating a v-shaped notch on the midline (Figure
2). Lateral ethmoids are situated posterolaterally ventrally to the mesethmoid, ventral to the frontal-mesethmoid joint and anterior to the orbitosphenoid (Figure 2). The lateral ethmoids are laterally expanded and cuplike. Lateral ethmoids are opened dorsally but covered by the anterior process of the frontal. The lateral ethmoids are much wider than the frontal process of the mesethmoid (Figure 3 and
Figure 4). Posterior to the lateral ethmoids is the orbitosphenoid that contributes to the floor and anterior sidewalls of the neurocranium. The floor of the orbitosphenoid is relatively thin (Figure 3 and
Figure 4). There is a sharp notch posteriorly on the orbitosphenoid where the parasphenoid plate attaches. The neck of the parasphenoid extends anteroventrally to the orbitosphenoid and contacts the posteriormost portion of the lateral ethmoids and median vomer (Figure 4).
 Along the parasphenoid neck, is a well-developed longitudinal crest that starts on the parasphenoid plate and terminates behind the vomer. The head of the kite-shaped vomer is ventral to the ethmoid cartilage and a deep trench in the mesethmoid. The vomer in Sarcoglanis is long but relatively narrower than the vomer in Stauroglanis (de Pinna 1989: figure 5). Along the parasphenoid neck, is a well-developed longitudinal crest that starts on the parasphenoid plate and terminates behind the vomer. The head of the kite-shaped vomer is ventral to the ethmoid cartilage and a deep trench in the mesethmoid. The vomer in Sarcoglanis is long but relatively narrower than the vomer in Stauroglanis (de Pinna 1989: figure 5).
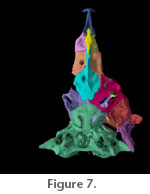
Paired frontals are located anterodorsally (Figure
2 and Figure 3;
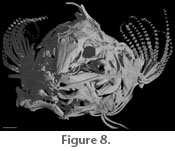
Figure 7). A wide supraorbital canal is present along the margin of posteriormost third of each frontal, and a canal pore is present at the anterior extremity of the supraorbital canal on the frontal. The supraorbital laterosensory canal continues along the margin of the sphenotic
 (Figure
3; Figure 8). The left and right supraorbital canals meet medially as in Microcambeva ribeirae (Costa et al. 2004, figure 3). The anterior mesethmoid process of the frontal is narrow and extends approximately half the length of the mesethmoid. The posteriormost third of the frontals widens abruptly behind the orbit and suture to one another. There is no anterior cranial fontanelle between the frontals, though a remnant fontanelle may be present in the form of a small circle between the frontal suture (Figure 1.2). The posterior margin of the frontals contacts the anterior margin of the supraoccipital that also lacks a midline fontanelle. Posterolaterally, the frontal contacts the sphenotic part of the compound sphenotic-prootic bone. (Figure
3; Figure 8). The left and right supraorbital canals meet medially as in Microcambeva ribeirae (Costa et al. 2004, figure 3). The anterior mesethmoid process of the frontal is narrow and extends approximately half the length of the mesethmoid. The posteriormost third of the frontals widens abruptly behind the orbit and suture to one another. There is no anterior cranial fontanelle between the frontals, though a remnant fontanelle may be present in the form of a small circle between the frontal suture (Figure 1.2). The posterior margin of the frontals contacts the anterior margin of the supraoccipital that also lacks a midline fontanelle. Posterolaterally, the frontal contacts the sphenotic part of the compound sphenotic-prootic bone.
The supraoccipital is broad across its joint with the frontals and the sphenotics (Figure
2 and Figure 3).
 A short shelf lies ventral to the posterior end of the frontal. Two anterolaterally diverging crests on the supraoccipital are continuous with the coronal crest of the sphenotic. Absent from the neurocranium of S. simplex are any anterolateral openings (de Pinna 1989). Areas of weak ossification, however, are present above the supraoccipital crests (Figure
3). Ventrally, there are elongate, slit-like gaps formed by the supracleithrum and Weberian complex (Figure 4). A short shelf lies ventral to the posterior end of the frontal. Two anterolaterally diverging crests on the supraoccipital are continuous with the coronal crest of the sphenotic. Absent from the neurocranium of S. simplex are any anterolateral openings (de Pinna 1989). Areas of weak ossification, however, are present above the supraoccipital crests (Figure
3). Ventrally, there are elongate, slit-like gaps formed by the supracleithrum and Weberian complex (Figure 4).
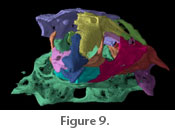
The supraorbital canal continues from the frontal through the sphenotic part of the sphenotic + prootic complex and connects to the postotic canal in the pterotic (Figure
9).
De Pinna (1989) recognized three bones in this complex: the sphenotic, prootic, and pterosphenoid. The postotic canal continues through to the supracleithrum. The sphenotic bone is small dorsally. Ventrolaterally, the sphenotic contacts the prootic. The prootic extends ventrally contacting the entire dorsolateral margin of the parasphenoid plate and the anteriormost margin of the basioccipital-Weberian complex (Figure 4 and
). The prootic is weakly ossified ventrally. Posterior to the prootic + sphenotic complex is the pterotic, which contributes to the lateral walls of the neurocranium. The pterotic contacts the sphenotic anterodorsally, the prootic anteriorly, the supraoccipital dorsally, the epiotic posterodorsally, and the supracleithrum posteriorly. Anterior to the supracleithrum is a short, thick process that extends anterodorsally off the lateral wall of the pterotic. We were unable to differentiate a separate pterosphenoid. There is, however, a small bridge of bone extending anterodorsally from the prootic to the orbitosphenoid contributing to a foramen for the passage of the optic, trigeminal, and facial nerves (Figure 5.1-2: rendered light pink). This bridge of bone may be the remnant of the pterosphenoid. Posterior to the pterotic compound is the supracleithrum. The supracleithrum is long and poorly ossified laterally. There is a small canal pore at the posterior limit of the supracleithrum adjacent to the lateral projection off the Weberian capsule (Figure 5). The prootic is weakly ossified ventrally. Posterior to the prootic + sphenotic complex is the pterotic, which contributes to the lateral walls of the neurocranium. The pterotic contacts the sphenotic anterodorsally, the prootic anteriorly, the supraoccipital dorsally, the epiotic posterodorsally, and the supracleithrum posteriorly. Anterior to the supracleithrum is a short, thick process that extends anterodorsally off the lateral wall of the pterotic. We were unable to differentiate a separate pterosphenoid. There is, however, a small bridge of bone extending anterodorsally from the prootic to the orbitosphenoid contributing to a foramen for the passage of the optic, trigeminal, and facial nerves (Figure 5.1-2: rendered light pink). This bridge of bone may be the remnant of the pterosphenoid. Posterior to the pterotic compound is the supracleithrum. The supracleithrum is long and poorly ossified laterally. There is a small canal pore at the posterior limit of the supracleithrum adjacent to the lateral projection off the Weberian capsule (Figure 5).
 Jaws. The premaxilla is located lateral to the mesethmoid cornua. It is an exceptionally long, slender, and toothless bone approximately three-quarters the length of the maxilla. The mesethmoid process of each premaxilla is short and hooked around its cornu, articulating both in front of and behind the cornua (Figure 6). Anteroventral to the mesethmoid process of the premaxilla is a small shelf that projects out under the cornua of the mesethmoid. There are two small projections of equal size on the proximal half of the premaxilla that are separated by a c-shaped gap (not shown in
Figure 6, see
Figure 9). This gap articulates with the premaxillary process of the maxilla. Distal to the maxillary articulation, the premaxilla tapers to a point. The distalmost end of the premaxilla is pointed and flattened anteroposteriorly. Jaws. The premaxilla is located lateral to the mesethmoid cornua. It is an exceptionally long, slender, and toothless bone approximately three-quarters the length of the maxilla. The mesethmoid process of each premaxilla is short and hooked around its cornu, articulating both in front of and behind the cornua (Figure 6). Anteroventral to the mesethmoid process of the premaxilla is a small shelf that projects out under the cornua of the mesethmoid. There are two small projections of equal size on the proximal half of the premaxilla that are separated by a c-shaped gap (not shown in
Figure 6, see
Figure 9). This gap articulates with the premaxillary process of the maxilla. Distal to the maxillary articulation, the premaxilla tapers to a point. The distalmost end of the premaxilla is pointed and flattened anteroposteriorly.
The maxilla is greatly enlarged. Proximally, the maxilla is laterally compressed. A short square process articulates with the mesethmoid posterior to the mesethmoid process of the premaxilla.
 The main articulation of the maxilla is with the large anterior palatine cartilage that fills the space between most the space between these bones (not shown). The premaxillary process of the maxilla is triangular and contributes to the deepest portion of the maxilla. Distal to this process the maxilla tapers gradually and twists slightly. The main articulation of the maxilla is with the large anterior palatine cartilage that fills the space between most the space between these bones (not shown). The premaxillary process of the maxilla is triangular and contributes to the deepest portion of the maxilla. Distal to this process the maxilla tapers gradually and twists slightly.
The palatine is located posterior to the maxilla and lateral to the mesethmoid (Figure 6,). The palatine articulates with a slight anterolateral concavity of the lateral ethmoid (Figure 5.1 and
Figure 6). The body of the palatine is rectangular with a long process projecting from the dorsolateral corner. There is a small anterior ossification of the palatine present. The lateral margin of the palatine is concave and continues from the short maxillary process of the palatine to the long, pointed posterior process. The maxillary process articulates in a facet on the internal side of the maxilla. The posterior process is shorter than that in Microcambeva and thinner than that in Stauroglanis.
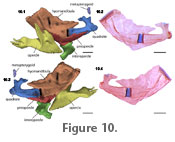 Suspensorium and operculars. There is a minute cylindrical metapterygoid located anterodorsal to the quadrate (Figure
10). The quadrate is relatively stout and is approximately the same length as the preopercles. There is a robust, round process that projects anteroventrally and articulates with the lower jaw, not treated in this analysis. Dorsal to this articular process is the hyomandibular process. The hyomandibular process is posteriorly recurved. It has two smaller processes that project from it. The anterior one articulates with the metapterygoid cartilage, and the posterior one comes close to but does not articulate with the hyomandibula. The hyomandibula is the largest bone in the suspensorium. It is roughly rectangular and flat. There is a short, wide process associated with the hyomandibular process of the quadrate. Across the body of the hyomandibula is a well-defined ridge (Figure
10.1-2). This ridge lacks a strong pointed process like that in Stauroglanis (de Pinna 1989). Suspensorium and operculars. There is a minute cylindrical metapterygoid located anterodorsal to the quadrate (Figure
10). The quadrate is relatively stout and is approximately the same length as the preopercles. There is a robust, round process that projects anteroventrally and articulates with the lower jaw, not treated in this analysis. Dorsal to this articular process is the hyomandibular process. The hyomandibular process is posteriorly recurved. It has two smaller processes that project from it. The anterior one articulates with the metapterygoid cartilage, and the posterior one comes close to but does not articulate with the hyomandibula. The hyomandibula is the largest bone in the suspensorium. It is roughly rectangular and flat. There is a short, wide process associated with the hyomandibular process of the quadrate. Across the body of the hyomandibula is a well-defined ridge (Figure
10.1-2). This ridge lacks a strong pointed process like that in Stauroglanis (de Pinna 1989).
The preopercle is a long, boomerang-shaped bone. It partially overlies the synchondral articulation between the quadrate and the hyomandibula in lateral view (Figure 7.1-2). The preopercle is strongly associated with these bones. Ventrally, it contacts the interopercle. The main body of the interopercle is square, but a posteriorly directed process is present on the posteroventral corner. There are no odontodes present on the interopercle. The opercle is approximately the same length as the hyomandibula. Posteriorly, there is a long process with a robust distal end from which three opercular odontodes project. Anteriorly, there is a pointed process that associates with a notch in the interopercle. A third, flat, well-developed process is present midway along the opercle that is aligned with the hyomandibular ridge and is likely the attachment site of the dilatator operculi muscles that in these trichomycterids is involved in elevating the patch of odontodes (Figure 7.1). This process is not present in Microcambeva and Stauroglanis; however, both these fishes have a shorter process that is continuous with the hyomandibular edge of the opercle (de Pinna 1989;
Costa et al. 2004).
|