|
|
|
DiscussionAll extant golden moles examined here except for Chrysospalax show dentaries that are slightly longer than their femora, comprising an average dentary length/femur length ratio near 1.1, one that is statistically indistinguishable in Neamblysomus, Amblysomus, Chrysochloris, and Eremitalpa. Association of the smaller femoral morphotype with dentaries assigned to either of the two C. asiatica-sized species named here also yields jaw-femur length ratios slightly over 1.1 (Table 1). The two extant species of Chrysospalax have femoral lengths between 40-120% larger than the largest chrysochlorid fossil recovered from Langebaanweg (Table 1). Due to its large size and the fact that larger mammals tend to have relatively smaller heads, C. trevelyani shows a dentary/femur length ratio below 1.0. Dentaries are substantially longer than humeral shafts in extant Chrysochloris, Amblysomus, Neamblysomus, and Chlorotalpa, exceeding a dentary/humerus length ratio of 1.24. This ratio is smaller in extant Eremitalpa and Chrysospalax, about 1.14 in the former and just over 1.0 in the latter. The associations of dentaries with humeri for the two species named here yield ratios of 1.19 for C. arenosa and 1.32 for C. bronneri. These ratios underscore our interpretation that C. arenosa is convergent on Eremitalpa and that C. bronneri more closely resembles other, similarly sized chrysochlorids in skeletal proportions (Table 1). Given that individual collecting sub-localities at Langebaanweg result from many years of opportunistic excavation and invariably represent a substantial amount of temporal and geographic averaging, we hesitate to ascribe much significance to the distribution within Langebaanweg of the chrysochlorid material described here. However, it is worth pointing out that the Langebaanweg "TCCW" sub-locality has produced two of the five large femora and three of the five large humeri. Other elements assigned to both new species described here derive from about 16 distinct sub-localities.
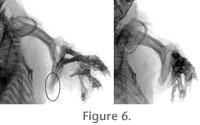
The distal humeral margin in the largest genus of golden mole, Chrysospalax, is also narrower relative to humerus length than it is in other chrysochlorids (Figure 2). This is consistent with the less fossorial and more terrestrial habitat of this golden mole relative to other genera (Nowak 1999). Furthermore, Chrysospalax humeral dimensions reflect the fact that longbones scale with positive allometry relative to most other body indices (Christiansen 2002). By virtue of its more terrestrial habitat and relatively large size, Chrysospalax shows a lower ratio of humerus length/distal width. We hypothesize that C. arenosa represents a distinctive early Pliocene chrysochlorid, closely related to the extant C. asiatica. Like modern Eremitalpa, this extinct golden mole was adapted to an arid environment in which it regularly burrowed in poorly-indurated sands and/or spent relatively more time aboveground. While remains of Eremitalpa have recently been recorded from the nearby West Coast National Park (Avery et al. 2005), the possibility that the Langebaanweg fossils represent a population of Eremitalpa can be ruled out by the fact that none of the cranial remains (e.g., mallei, maxillae, dentaries) resemble the distinctive morphology of that taxon (e.g., short palates, absent protocones on P3, ovoid mallei). Another genus of golden mole, Cryptochloris, is sympatric with some Eremitalpa populations (Meester et al. 1986). Indeed, specimens of Cryptochloris are frequently mislabeled as Eremitalpa in museum collections as the two are externally very similar in appearance. Unfortunately, properly macerated skeletons of Cryptochloris are nonexistent in museum collections, and we therefore lack comparative data on their humeral proportions. However, radiographic images (Figure 6) show that the distal humeral width is not as reduced in Cryptochloris wintoni as it is in Eremitalpa or our hypodigm of C. arenosa. Otherwise, Cryptochloris is anatomically similar to Chrysochloris, showing 10 teeth in each molar quadrant, no talonids on its molars or premolars, and a club-shaped malleus. It differs from C. asiatica in that its temporal bulla is smaller, and the maxillae are more expanded dorsally on the rostrum than on the alveolar margins of the palate. Cryptochloris also shows similarly sized upper incisors and a larger first digit than C. asiatica (Helgen and Wilson 2003). We believe the morphology of the maxilla, premaxilla, incisors, and humerus of the Langebaanweg chrysochlorids rule out their possible membership in Cryptochloris. No craniodental material is yet known that fits the size range expected for the large morph, which also exhibits a relatively narrow distal humeral margin. This taxon is significantly smaller than extant Chrysospalax, and as in the small Langebaanweg morph may have been adapted for sand-swimming and/or spending relatively more time aboveground than extant golden moles in its size range. |
|