|
|
|
Systematic Paleontology
Afrotheria Stanhope et al. 1998 Type locality: Langebaanweg, South Africa
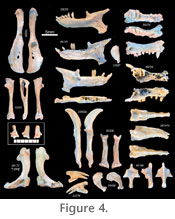
Holotype: SAM-PQL 20177 right humerus (Figure 4). Age: early Pliocene Diagnosis: Humeri show a significantly narrower distal margin (Figure 2) than those of extant C. asiatica. Based on relative abundance, humeri are associated with well-preserved maxillae and dentaries (Figure 4), containing 10 teeth in each dental quadrant and P3 protocones, similar in morphology to C. asiatica. Material: All specimens have the acronym SAM-PQL. The following are dentaries assigned to this species and sufficiently complete to measure mandibular condyle to the symphysis: 20261, 35070, 35074, 35397, 57981, 67943, 69712, 69713, 69714, 69715, 69716, 69717, 69718, 69719, 69720, 69721, 69722, 69723, 69725, 69728, 69729, 69731, 69732, 69733, 69734, 69735, 69736, 69737, 69738, 69739, 69740, 69741, 69742, 69744, 69748, 69749, 70253, 70254, 70266, 70267, 70268, 70269, 70270. Rostral fragments include SAM-PQL 35073 (left maxilla with C, P2, P4-M1), SAM-PQL 43200 (left maxilla with P3-P4), and SAM-PQL 69726 (right rostral fragment with I1-2, C, P3-4). Based on relative abundance, we assign a representative malleus (SAM-PQL 13337), ulna (SAM-PQL 70250), radius (SAM-PQL 35208), terminal phalanx (SAM-PQL 23776), innominate (SAM-PQL 70252), and tibia-fibula (SAM-PQL 70251) to this species. Description: Relatively complete dentaries and maxillae (Figure 4) show alveoli and/or teeth corresponding to a dental formula of 3.1.3.3, with a total of 10 teeth in each dental quadrant. In addition, all maxillary fragments that preserve cheek teeth show protocones on M1-2 and P3-4, but not on P2 or farther anteriorly. No specimen preserves M3, but alveoli for this locus are present. P2 exhibits a posterobucally extending mesostyle (e.g., SAM-PQL 35073). The canine is small and conical, as are I2-3. As in C. asiatica and other golden moles, I1 is the largest of the incisors and is rooted in the premaxilla at an angle, so that it points into the oral cavity when viewed laterally (Figure 4). Similarly, premaxillary remains exhibit two prongs jutting out anterolaterally from either side of the external nares. In addition, M1 is situated just medial to the anterior root of the zygomatic arch, immediately ventral to the short, bridge-like infraorbital canal, evident in dorsolateral aspect. No basicranial remains have yet been identified as chrysochlorid. However, hundreds of ear ossicles recovered from Langebaanweg are easily recognizable and resemble those of modern C. asiatica. They are elongate, ovoid, and tear-drop shaped (Figure 4), rather than the distinctive pea-shape of Eremitalpa or Chrysospalax. Relative abundance and size justifies their association with the most common humeral and mandibular morphotype, i.e., Chrysochloris arenosa. All of the dentaries assigned to C. arenosa are similar in overall size and show little variation in qualitative morphology (Figure 4). As in the modern Cape golden mole (C. asiatica), they have 10 teeth and/or alveoli, molariform p3-m3, and molars and premolars without talonids. As for all known chrysochlorids, the Langebaanweg dentaries also show an i2 larger than adjacent incisors and a reduced coronoid process, projecting dorsally towards the transverse level of the mandibular condyle. Variation exists in the presence of a 2-rooted p4; specimens that are otherwise identical may differ in this respect (e.g., SAM-PQL 69720 with a 2-rooted p4 vs. SAM-PQL 69737 without). We observed similar variation in modern C. asiatica: e.g., SAM-ZM 39362 with a 2-rooted p4 vs. SAM-ZM 37130 without. In addition, some specimens preserving the mandibular angle, known in chrysochlorids to articulate with the hyoid apparatus (Bronner 1991), show a flat hyoid articular surface (e.g., SAM-PQL 69725); in other specimens this surface is curved (SAM-PQL 35070; see Figure 4). We also attribute these differences to intraspecific variation. Humeri (Figure 4) show the characteristic chrysochlorid shape of the proximal humeral condyle, i.e., dorsoventrally elongate and somewhat teardrop-shaped. The greater tuberosity, just lateral to the humeral head, is smaller than the lesser tuberosity, which extends medially from the condyle. The shaft of the humerus shows a slight, medially concave curvature, culminating in the extremely elongate distal humeral margin, the medial tip of which would have articulated with an elongate, ossified flexor tendon, or third bone of the forearm. While no ossified flexor tendons have been identified from the Langebaanweg material, great similarity in other aspects of forelimb morphology makes it likely that these fossil chrysochlorids also possessed this structure. Along the anteromedial surface of the humeral shaft, towards the distal end of the diaphysis, a small flange is evident that contributes to a medial concavity of the distal humeral shaft, adding yet more surface area for forearm musculature. In the extant C. asiatica and A. hottentotus, humeri have a distal margin that is nearly as large as the humerus itself, showing a ratio of humeral length/distal width close to 1, and not exceeding 1.15 (Figure 2). In contrast, humeri from Langebaanweg assigned to C. arenosa (e.g., SAM-PQL 20177), have a shorter distal margin, yielding ratios of humeral length/distal width between 1.3 and 1.5, averaging 1.39 (Figure 2). This is the key feature distinguishing C. arenosa from modern C. asiatica. As discussed above, craniodental features clearly distinguish C. arenosa from other chrysochlorids with a relatively narrow distal humerus, such as Eremitalpa and Chrysospalax. Isolated synsacra (Figure 4) consist of 2–5 vertebrae, the neural spines of which fuse to form a dorsal, craniocaudally running ridge. Sacral centra also tend to fuse to one another. More variable is the position of the articular scar for the ilia. Most specimens (e.g., SAM-PQL 35108) show an articular surface on the lateral aspect of both S1 and S2 (Figure 4). However, in some specimens (e.g., SAM-PQL 35239) this surface is caudally displaced, comprised of S2-3 with no contribution from S1. These variations match intraspecific variability observed in recent specimens of C. asiatica. Isolated innominate bones (Figure 4) show typical chrysochlorid morphology. The pubis and ischium extend caudally from the acetabulum, parallel to one another, framing a minute obturator foramen that is smaller than the acetabulum. Occasionally the obturator foramen is partially fused, leading in some cases to two small foramina in this region. An articular surface for the sacrum is evident towards the cranial end of the ilium, but does not extend to the cranial tip of the ilium, which shows a slight lateral flare. The tibia-fibula is fused both proximally and distally (Figure 4). Proximally, the fibula shows a laterally projecting condyle, typical for all extant golden moles. Also typical is the concave facet at the base of the fibula for articulation with a ball-like process extending dorsally from the ectal facet of the calcaneus. A single calcaneus was attached in matrix to the distal end of SAM-PQL 70251 (Figure 4). Distolaterally it shows a prominent, dorsally projecting condyle that articulates with the aforementioned facet of the distal fibula. Its distal margin for articulation with other tarsal elements is relatively flat; a small peroneal process is present that extends slightly past the distal calcaneal articular surface. The sustentaculum tali is present but poorly defined, with little indication of the contour of the articulation with the astragalus. The manubrium sternae exhibits a sharp keel that juts anteroventrally (Figure 4). The keel grades in a straight line to form posteriorly a prominent condyle for articulation with the adjacent sternebra. The first rib articulates into the angle created by the caudally jutting condyle as it turns laterally to define alae on each side of the midline. The first rib (Figure 4) has a typical chrysochlorid appearance, i.e., it is flattened and expands distally to form a paddle-shape, with articular surfaces distally for the sternum and proximally for T1. Radii are distinguished proximally by a single, concave facet for articulation with the humeral capitulum (Figure 4). Distally the radius shows two facets for articulation with the scaphoid and lunate. Several specimens preserve a narrow bridge of bone near the radial distal margin that presumably enclosed a sheath of extrinsic digital flexors, often accompanied by a conspicuous trough. Most specimens that lack this bridge show evidence of postmortem abrasion on the distal radius and probably had one in life. Modern C. asiatica also shows this structure (e.g., SAM-ZM 37130). Ulnae also match closely the morphology and size seen in C. asiatica (Figure 4). Both have an elongate, medially curved olecranon process, nearly as long as the shaft of the ulna distal to the humeral articular surface. The proximal articular facet for the humerus extends farther laterally than does the distal facet. The styloid process is narrow and pointed; articulations with the carpus are not limited to the styloid itself but are also present along the distal ulnar shaft. The enlarged, terminal phalanx closely resembles that of digit III in modern C. asiatica (Figure 4). The phalanx ends in two points, both of which would have been covered in life with a large, keratinous claw. Proximally, the lateral articular facet for the adjacent, intermediate phalanx is dorsoventrally longer but mediolaterally shorter than the medial articular facet. Other carpal elements have yet to be identified. Chrysochloris bronneri sp. nov. Type locality: Langebaanweg, South Africa
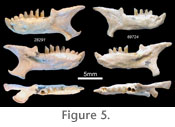
Holotype: SAM-PQL 28291 left dentary with i2-3, c, p2-m2 (Figure 5). Age: early Pliocene Diagnosis: The dentary is more robust than that of extant C. asiatica or contemporaneous C. arenosa, and shows a stout, enlarged lower i2 without a distinct posterior cusp and with a root that broadens as it enters the mandibular corpus. Based on relative abundance, we associate the second most common humeral morphotype (Figure 1) with the type dentary. These show a humerus length/distal width ratio significantly smaller than that of C. arenosa, close to that of extant N. julianae and C. asiatica (Figure 2). Material: SAM-PQL 69724 (right dentary with i2, p2-m1). Humeri include SAM-PQL 32817, 66814, 70255, 70256, 70256, 70257, 70258, 70259, 70260, 70261, 70261, 70262, 70263, 70264, 70265, 70271, 70272, 70273, 70274, 70275, 70276, 70277, 70278, 70279, 70280. Description: SAM-PQL 28291 and 69724 (Figure 5) show a broad, squat i2 without a posterior cusp and with a root that does not begin to narrow until it is well into the mandibular corpus, and which shows more curvature as it descends into the corpus than does the i2 of C. arenosa. The lack of a posterior cusp may be accentuated by wear in both specimens. However, the size and robustness of the tooth clearly indicate a marked difference from the gracile i2 of the many dentaries that preserve either the alveolus or the tooth itself, including 43 other specimens complete enough to measure condyle-symphysis distance. Deciduous teeth in golden moles, as with other afrotherians, may remain unreplaced for some time into adulthood (Leche 1907; Asher and Lehmann 2008). However, the possibility that these enlarged teeth represent deciduous precursors is unlikely, given the lack of erupting or unworn teeth at other loci, and since deciduous anterior teeth of chrysochlorids are generally similar in size or smaller than their permanent replacements. Enlarged, deciduous incisors that function throughout adulthood do occur among mammals (e.g., rodents and lagomorphs); we cannot rule out the possibility that this has occurred in a golden mole. If so, it would be a first for the group and would further comprise morphological support for the existence of a new species. Whatever the identity of the enlarged i2, we interpret the two Langebaanweg individuals that have it as a second, C. asiatica-sized species of early Pliocene golden mole, here named Chrysochloris bronneri. One of these dentaries (SAM-PQL 69724) appears at first glance to lack an alveolus for i1, suggesting only nine teeth in each dentary. SAM-PQL 69724 also seems to have a narrower angular process than SAM-PQL 28291. However, we believe both the dental formula and the angular process morphology in these specimens are in fact the same as in other specimens. SAM-PQL 69724 shows evidence of postmortem abrasion along the mandibular symphysis and angular process (Figure 5). Furthermore, dental reduction among living chrysochlorids, from 10 to 9 teeth in each quadrant, results from loss of the third molar, not the first incisor. Hence, as for all other chrysochlorid dentaries from Langebaanweg that preserve the relevant anatomy, it is likely that this specimen also had 10 teeth in each jaw and a relatively broad angular process. |
|