|
|
|
Discussion aND CONCLUSIONSHesperolagomys was originally described as an ochotonid by Clark et al. (1964), who noted its dental similarities to both Amphilagus fontannesi, a late Miocene European ochotonid, and Desmatolagus gobiensis from the Oligocene of Asia. Whereas most subsequent authors have accepted this interpretation, Storer (1984) challenged it, suggesting that the genus should belong to the Leporidae (see also McKenna and Bell 1997, p. 109; for an additional alternate view, see Erbajeva (1988; 1994). The basis for Storer's suggestion appears to be his assertion that undescribed maxillary fragments assigned to Hesperolagomys lack a premolar foramen. Absence of this feature, if confirmed by published descriptions, would clearly necessitate radical reassessment of the affinities of the genus because a premolar foramen has been regarded as diagnostic of the Ochotonidae by major students of lagomorph evolution beginning with Bohlin (1942) and followed by Dawson (1967; see also Dawson 2008) and McKenna (1982). All of the Hesperolagomys maxillae reported here, including one from the type locality of Fish Lake Valley, have a distinct premolar foramen, confirming that the genus belongs in the Ochotonidae. Other diagnostic ochotonid features (Bohlin 1942) of Hesperolagomys were also confirmed in the present study: (1) bases of upper cheek teeth curving into the zygoma (not extending into orbit); (2) in lower molariform teeth, trigonid and talonid not united lingually with wear; (3) M3 absent; (4) P3 nonmolariform; (5) m3 lacking talonid; (6) mental foramen posterior to its 'usual' position in leporids. An additional characteristic considered diagnostic of Ochotonidae by McKenna (1982), lower incisor extending posteriorly at least to p4, is also confirmed for all three Hesperolagomys species studied here. In summary, there is no longer any reason to doubt that Hesperolagomys is an ochotonid. Characteristics of HesperolagomysClark et al. (1964) described Hesperolagomys as "unusually primitive" compared to other Miocene ochotonids, possessing plesiomorphic characters including: (1) rooted cheek teeth; (2) buccal folds persisting on the occlusal surface of P4 and M1; and (3) talonids transversely narrower than trigonids in p4-m2. Additionally, the posterior extent of the lower incisor to beneath m1 is plesiomorphic. Apomorphic characters of Hesperolagomys noted by Clark et al. (1964) are the unique position and emphasis of mental foramina, and "marked anterior projections of the talonids on p4-m2." Additional apomorphies are P3 with two persistent cement-filled lingual striae, and P4-M2 with persistent crescentic valleys and deep persistent lingual hypostriae. At the time of its initial description, Hesperolagomys was arguably the most 'primitive' late Miocene ochotonid; Oreolagus (possessing many 'advanced' dental characters including rootless cheek teeth) was the only other North American Miocene ochotonid known. A greater diversity of 'primitive' North American forms have now been described, including Gripholagomys, Russellagus, and ?Desmatolagus schizopetrus, but the relationship of Hesperolagomys to other ochotonid taxa has not been defined. Clark et al. (1964) suggested that Hesperolagomys might be derived from Desmatolagus, either from North American or Asian forms. Storer (1975) noted that Hesperolagomys and Russellagus are clearly related, but show distinct morphological differences. Working with only small samples of primarily isolated teeth, Storer successfully described pertinent characteristics of Russellagus vonhofi and H. fluviatilis, but was unable to correctly identify some isolated teeth due to the remarkable morphological similarity between some wear stages of teeth of the two taxa. Martin (1976) suggested a close relationship between the more poorly known taxon Gripholagomys and Hesperolagomys. Revision of Russellagus and Gripholagomys by the author (in progress) should elucidate the nature of their relationship to Hesperolagomys and other North American taxa. Characteristics and Relationships of Hesperolagomys SpeciesMajor differences between Hesperolagomys species are summarized in Table 7. Size of most dimensions is important in distinguishing the species, with H. fluviatilis smallest, H. galbreathi intermediate, and H. niobrarensis largest. H. galbreathi overlaps with both H. fluviatilis and H. niobrarensis in some dimensions and is difficult to distinguish from the older taxa on size only; most elements of H. fluviatilis are distinguishable from those of H. niobrarensis on the basis of size alone. Lower third premolar of H. fluviatilis is anteroposteriorly longer than that of the other species, both absolutely and relatively. Morphological characters differing between species are primarily those of lower cheek tooth shape. The trigonid of p3 of H. fluviatilis extends more anteriad than in other species, in which it extends more lingually; p3 of H. fluviatilis also lacks an anterolingual groove, present in H. niobrarensis and H. galbreathi. The talonid of lower molariform teeth of H. galbreathi is consistently more buccolingually symmetrical than those of H. fluviatilis and H. niobrarensis; although the occlusal morphology of the talonid does change with wear, those of H. fluviatilis and H. niobrarensis are more asymmetrical and can be distinguished from H. galbreathi in all but the earliest stages of wear.
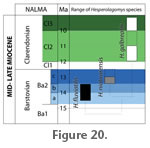
Age ranges of Hesperolagomys species are shown in Figure 20. The first appearance of Hesperolagomys is with H. fluviatilis near the beginning of the late Barstovian (Ba2ba), reinforcing the view of Tedford et al. (2004). H. fluviatilis apparently persisted until near the middle of the late Barstovian (Ba2b). H. niobrarensis (sp. nov.) is restricted to the late late Barstovian (Ba2c) and is currently known only from Nebraska. All known Hesperolagomys specimens of medial and late Clarendonian age (Cl2-3) are consistent with H. galbreathi. At this time, no ochotonid specimens of early Clarendonian (Cl1) age are known; in Nebraska, at least, their absence from the Burge and equivalent faunas appears to reflect poor recovery of small mammal fossils as compared with their much greater diversity in Barstovian faunas (e.g., Voorhies 1990a, Table 2 and Table 3). The last appearance of Hesperolagomys is near the end of the Clarendonian, apparently disappearing as part of an extinction event extirpating many taxa characteristic of the medial Miocene (Tedford et al. 2004). The cause of the extinction of Hesperolagomys and other dentally archaic ochotonids at the end of the Clarendonian is unknown. Bair (2007) used a geometric model of wear in curved mammal teeth to demonstrate that the upper cheek teeth of Hesperolagomys and other dentally archaic ochotonids were constrained from attaining greater crown height while maintaining their unilateral hypsodonty and curved shape, and suggested that these constraints may account for the Miocene transition in dominance from dentally archaic ochotonids to dentally "advanced" leporids. Potts et al. (1992) suggest that ochotonids were among mammalian taxa which "successfully colonized the closed habitat of the savanna mosaic but did not radiate significantly into the more open environments during the Miocene." Preliminary analysis suggests that Clarendonian Hesperolagomys may have been restricted to relatively closed, mesic settings such as those near ponds or lakes (e.g., Fish Lake Valley, Nevada; fossil fish-rich deposits of Tiensvold Ranch, Nebraska). While these hypotheses are compelling and internally consistent, further work testing both is necessary. |
|