| |
RESULTS
 In
Figure 5 we display the data obtained from the classical mesowear analysis of the upper molars. The result of the hierarchical cluster analysis is shown in
Figure 6. We observe that the cusps of the Recent extreme grazer Ceratotherium simum are low and almost exclusively blunt. In the fossil species Coelodonta antiquitatis the cusps are also mainly low and mostly blunt and rounded, but other kinds of cusps are also present in small numbers. In the Recent graze-dominated mixed feeder Rhinoceros unicornis the cusps are rounded and blunt, and mostly low. Conversely, in the Recent browser species Diceros bicornis, Dicerorhinus sumatrensis, and R. sondaicus the cusps are mainly high and sharp or rounded, which is also true of the cusps in the fossil species Stephanorhinus hundheimensis and S. kirchbergensis. In
Figure 5 we display the data obtained from the classical mesowear analysis of the upper molars. The result of the hierarchical cluster analysis is shown in
Figure 6. We observe that the cusps of the Recent extreme grazer Ceratotherium simum are low and almost exclusively blunt. In the fossil species Coelodonta antiquitatis the cusps are also mainly low and mostly blunt and rounded, but other kinds of cusps are also present in small numbers. In the Recent graze-dominated mixed feeder Rhinoceros unicornis the cusps are rounded and blunt, and mostly low. Conversely, in the Recent browser species Diceros bicornis, Dicerorhinus sumatrensis, and R. sondaicus the cusps are mainly high and sharp or rounded, which is also true of the cusps in the fossil species Stephanorhinus hundheimensis and S. kirchbergensis.
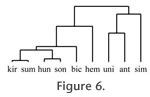 The fossil species S. hemitoechus in turn differs from Recent browsers in that the cusps are mostly rounded, with some blunt and sharp cusps present as well. In hierarchical cluster analysis the browsers, including the Recent species R. sondaicus, D. sumatrensis, and D. bicornis as well as the fossil species S. hundsheimensis and S. kirchbergensis, form a separate group. Another group is formed by the species R. unicornis, C. antiquitatis, and C. simum, all of whose diet is more abrasive. The species S. hemitoechus falls between these two main groups in the analysis. The fossil species S. hemitoechus in turn differs from Recent browsers in that the cusps are mostly rounded, with some blunt and sharp cusps present as well. In hierarchical cluster analysis the browsers, including the Recent species R. sondaicus, D. sumatrensis, and D. bicornis as well as the fossil species S. hundsheimensis and S. kirchbergensis, form a separate group. Another group is formed by the species R. unicornis, C. antiquitatis, and C. simum, all of whose diet is more abrasive. The species S. hemitoechus falls between these two main groups in the analysis.
 In
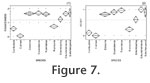
Figure 7 we display the data obtained from the mesowear analysis of the upper molars, with the data factored according to roundedness in
Figure 7.1 and according to height in
Figure 7.2. With respect to both of these factors separately, the browsers D. bicornis, D. sumatrensis, R. sondaicus, S.
hundsheimensis, and S. kirchbergensis clearly form a group, which is separated from the species S. hemitoechus. Thus we conclude that the cusps are, on average, both lower and more rounded in the species S. hemitoechus than in browsers. On the other hand, the species C. antiquitatis in turn falls between the extreme grazer C. simum and the graze-dominated mixed-feeder R. unicornis, which suggests that its cusp apices are, on the average, not as low or as blunt as in C. simum. In
Figure 7 we display the data obtained from the mesowear analysis of the upper molars, with the data factored according to roundedness in
Figure 7.1 and according to height in
Figure 7.2. With respect to both of these factors separately, the browsers D. bicornis, D. sumatrensis, R. sondaicus, S.
hundsheimensis, and S. kirchbergensis clearly form a group, which is separated from the species S. hemitoechus. Thus we conclude that the cusps are, on average, both lower and more rounded in the species S. hemitoechus than in browsers. On the other hand, the species C. antiquitatis in turn falls between the extreme grazer C. simum and the graze-dominated mixed-feeder R. unicornis, which suggests that its cusp apices are, on the average, not as low or as blunt as in C. simum.
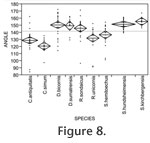 The data obtained from the analyses of the lower molars are displayed in Figure 8 and
Figure 9. The angles between the enamel edges and the buccal surfaces as measured in the hypoconid, and protoconid tip regions are shown in
Figure 8. The angles are clearly larger on the average for the browsers D. bicornis, D. sumatrensis, R. sondaicus, S.
hundsheimensis, and S. kirchbergensis than for other species. Thus these browsers form a clearly separate group. The angles related to the species S. hemitoechus are somewhat smaller. On the average, the smallest angles were those measured from the molars of the Recent grazer C. simum, whereas the average of the angles of the species C. antiquitatis were smaller than the average of the angles of the Recent mixed feeder R. unicornis. The data obtained from the analyses of the lower molars are displayed in Figure 8 and
Figure 9. The angles between the enamel edges and the buccal surfaces as measured in the hypoconid, and protoconid tip regions are shown in
Figure 8. The angles are clearly larger on the average for the browsers D. bicornis, D. sumatrensis, R. sondaicus, S.
hundsheimensis, and S. kirchbergensis than for other species. Thus these browsers form a clearly separate group. The angles related to the species S. hemitoechus are somewhat smaller. On the average, the smallest angles were those measured from the molars of the Recent grazer C. simum, whereas the average of the angles of the species C. antiquitatis were smaller than the average of the angles of the Recent mixed feeder R. unicornis.
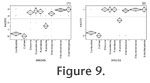 In
Figure 9.1 we show the data obtained when we classified the lower molars into five groups according to the number of facets on the buccal side of the enamel edge.
Figure 9.2 in turn shows the data obtained with the simpler variation of the method, where the molars were classified into two groups according to whether they have facets or not. In both of these datasets the browsers clearly form a separate group, which is explained by well-developed facets. The species S. hemitoechus presents facets on most molars, the mixed feeder R. unicornis presents some facets, whereas the grazer C. simum does not present facets at all. Once again, the species C. antiquitatis falls between the species C. simum and R. unicornis. In
Figure 9.1 we show the data obtained when we classified the lower molars into five groups according to the number of facets on the buccal side of the enamel edge.
Figure 9.2 in turn shows the data obtained with the simpler variation of the method, where the molars were classified into two groups according to whether they have facets or not. In both of these datasets the browsers clearly form a separate group, which is explained by well-developed facets. The species S. hemitoechus presents facets on most molars, the mixed feeder R. unicornis presents some facets, whereas the grazer C. simum does not present facets at all. Once again, the species C. antiquitatis falls between the species C. simum and R. unicornis.
It is rather laborious to manually measure the angle of the enamel edge and, contrary to mesowear analysis, it cannot therefore be used to process large numbers of samples "at a glance." Therefore we also investigated whether the lower molars could be analysed by a fast visual method. The only feature that can be determined with sufficient rapidity and simplicity for our purposes is whether or not the buccal side enamel edge of a lower molar does or does not have facets. We applied this idea in two different ways. In the first case we studied separately the lack or existence of the facets one, two, six, and seven, and counted the total number of facets present. Hence we arrived at a number between 0 and 4, with 0 indicating no facets and 4 indicating that facets are present everywhere on the enamel edge. The results we obtained with this method (Figure 9.1) match the results obtained by mesowear analysis of upper molars (Figure 7). The second analysis we applied was even simpler. There we classified the lower molars into two groups according to whether they have buccal side facets or not. As can be seen from Figure 9.2, the data obtained by this analysis also allow us to separate browsers, mixed feeders, and grazers from each other. The benefit of this particular analysis is that it is easy to use and fast, but it is slightly less accurate than the other analyses considered here. Its accuracy is sufficient for browsers and grazers to be discriminated from each other, but it places the species S. hemitoechus close to the browsers as only one of the studied molars was completely facetless. On the other hand, as the other analyses studied, it is able to separate the Recent mixed feeder R. unicornis from the grazer C. simum and the browsers, and the species C. antiquitatis from the species C. simum and R. unicornis.
|