

 A description of small gomphotheriid (Mammalia, Proboscidea) manuses from the earliest Pleistocene 111 Ranch Beds, southeast Arizona, USA
A description of small gomphotheriid (Mammalia, Proboscidea) manuses from the earliest Pleistocene 111 Ranch Beds, southeast Arizona, USA
Article number: 17.1.7A
https://doi.org/10.26879/421
Copyright Society for Vertebrate Paleontology, February 2014
Author biography
Plain-language and multi-lingual abstracts
PDF version
Submission: 23 July 2013. Acceptance: 23 January 2014
{flike id=661}
ABSTRACT
A partial left and right manus of a gomphotheriid are described from earliest Pleistocene deposits of 111 Ranch in southeastern Arizona, USA. The discovery of these well preserved manuses provides a rare opportunity to describe these elements of a gomphotheriid in North America. I provide detailed descriptions and measurements as well as comparisons of the manuses with those of other extant and extinct proboscideans. Analyses of the manuses indicate it was a small gomphotheriid, possibly a small female, having morphological attributes more similar to other gomphotheriids, then elephantids and mammutids. The gomphotheriids Stegomastodon and Rhynchotherium have both been previously reported from 111 Ranch.
Michael R. Pasenko. Environmental Planning Group, 4141 North 32nd Street, Suite 102, Phoenix, Arizona, 85018, USA, mammut501@yahoo.com, Mpasenko@epgaz.com
Keywords: manus; carpal; metacarpal; gomphotheriid; 111 Ranch
Final citation: Pasenko, Michael R. 2014. A description of small gomphotheriid (Mammalia, Proboscidea) manuses from the earliest Pleistocene 111 Ranch Beds, southeast Arizona, USA. Palaeontologia Electronica Vol. 17, Issue 1;7A; 15p. https://doi.org/10.26879/421
palaeo-electronica.org/content/2014/661-gomphotheriid-manus
INTRODUCTION
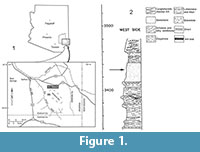 Gomphotheriidae is a diverse family of extinct proboscideans sharing the characteristic of bunolophodont teeth. Although much research on North American gomphotheriids has taken place, very little has focused on the postcrania, especially the feet. The recovery of two partial manuses in 1999, within the 111 Ranch Beds provided the opportunity to describe these elements in detail. The 111 Ranch is located north of the Whitlock Mountains, near Safford, Arizona (Figure 1.1), but the 111 Ranch Beds occur over a larger area west and southwest of the Whitlock Mountains (see QTlf of Richter et al., 1981). The original description of the geological units (Figure 1.2) at 111 Ranch included sands, gravels, silts, marls, limestones, diatomite, chert, and tuff (Seff, 1960; 1962; Galusha et al., 1984), which Houser (1990) considered indicative of lacustrine and fluvial conditions. The manuses were discovered in the silty sandstones at an approximate elevation of 1045 meters (3430 feet) above sea level (Skaff, 2001; Thrasher, personal commun.). The 111 Ranch local fauna and several other local faunas found in the San Simon River Valley are believed to be correlative and are now grouped together as part of the "111 Ranch Beds" (Tomida, 1987; Houser, 1990; Thrasher, 2007). Radiometric and fission-track dates for the 111 Ranch local fauna are between 2.3 and 2.47 Ma (Dickson and Izett, 1981; Galusha et al., 1984; Houser et al., 2004), indicating an earliest Pleistocene age. The gomphotheriids Rhynchotherium, Stegomastodon, and Cuvieronius have all been reported from the 111 Ranch Beds (Wood, 1962; Lindsay and Tessman, 1974; Skaff, 2001; Morgan and White, 2005; Pasenko, 2007; Pasenko and Lucas, 2011), but there are no published descriptions of the Cuvieronius material, which have been referred to as cf. Cuvieronius or Cuvieronius sp. in the literature (Lindsay, 1978, Galusha et al., 1984). A Stegomastodon molar was described from 111 Ranchin Wood's (1962) dissertation and more recently an undescribed skull, mandible, and partial skeleton of Stegomastodon have been found at 111 Ranch (AZMNH collections). Two specimens of Rhynchotherium, including two mandibles and a partial skeleton, have been previously found at 111 Ranch (Skaff, 2001; Pasenko, 2007, 2012).
Gomphotheriidae is a diverse family of extinct proboscideans sharing the characteristic of bunolophodont teeth. Although much research on North American gomphotheriids has taken place, very little has focused on the postcrania, especially the feet. The recovery of two partial manuses in 1999, within the 111 Ranch Beds provided the opportunity to describe these elements in detail. The 111 Ranch is located north of the Whitlock Mountains, near Safford, Arizona (Figure 1.1), but the 111 Ranch Beds occur over a larger area west and southwest of the Whitlock Mountains (see QTlf of Richter et al., 1981). The original description of the geological units (Figure 1.2) at 111 Ranch included sands, gravels, silts, marls, limestones, diatomite, chert, and tuff (Seff, 1960; 1962; Galusha et al., 1984), which Houser (1990) considered indicative of lacustrine and fluvial conditions. The manuses were discovered in the silty sandstones at an approximate elevation of 1045 meters (3430 feet) above sea level (Skaff, 2001; Thrasher, personal commun.). The 111 Ranch local fauna and several other local faunas found in the San Simon River Valley are believed to be correlative and are now grouped together as part of the "111 Ranch Beds" (Tomida, 1987; Houser, 1990; Thrasher, 2007). Radiometric and fission-track dates for the 111 Ranch local fauna are between 2.3 and 2.47 Ma (Dickson and Izett, 1981; Galusha et al., 1984; Houser et al., 2004), indicating an earliest Pleistocene age. The gomphotheriids Rhynchotherium, Stegomastodon, and Cuvieronius have all been reported from the 111 Ranch Beds (Wood, 1962; Lindsay and Tessman, 1974; Skaff, 2001; Morgan and White, 2005; Pasenko, 2007; Pasenko and Lucas, 2011), but there are no published descriptions of the Cuvieronius material, which have been referred to as cf. Cuvieronius or Cuvieronius sp. in the literature (Lindsay, 1978, Galusha et al., 1984). A Stegomastodon molar was described from 111 Ranchin Wood's (1962) dissertation and more recently an undescribed skull, mandible, and partial skeleton of Stegomastodon have been found at 111 Ranch (AZMNH collections). Two specimens of Rhynchotherium, including two mandibles and a partial skeleton, have been previously found at 111 Ranch (Skaff, 2001; Pasenko, 2007, 2012).
Institutional Abbreviations. AMNHAmerican Museum of Natural History, New York; AZMNH Arizona Museum of Natural History, Mesa; HSM Hot Springs Mammoth Site, South Dakota; LACM Natural History Museum Los Angeles County, California; UALP University of Arizona Laboratory of Paleontology, Tucson
MATERIALS AND METHODS
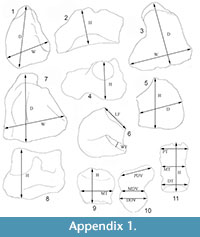 Specimen AZMNH p7487 consists of partial left and right gomphotheriid manuses. These manuses described here were first mentioned in Skaff (2001) who listed, but did not describe the elements recovered. The carpals are referred to by name and for the first use the corresponding synonymous names are added. The metacarpals are referred to as Mc followed by a number (e.g., Mc3 = metacarpal of the third digit). The following measurements are used for the carpals: the greatest proximal-distal height, greatest dorsal-volar depth (D-V), and greatest transverse width, as well as available measurements of the articular facets. For the metacarpals and phalanges the following measurements are used: greatest proximal-distal length (height), proximal dorsal-volar depth, proximal transverse width, distal dorsal-volar depth, distal transverse width, and minimum transverse width (Appendix 1). The identification of the carpals, metacarpals, and phalanges was based on their position when discovered, relative size, and comparisons to other proboscidean specimens from the literature and in museum collections. When possible, the size and morphology of specific elements were compared to the same elements of other extinct and extant proboscideans including Gomphotherium steinheimense (Göhlich, 1998), Haplomastodon chimborazi (Ferretti, 2010), Rhynchotherium falconeri (Frick, 1933; Pasenko, 2007, 2011), Stegomastodon primitivus (Alberdi et al., 2009; Lucas et al., 2011), Mammut americanum (Olsen, 1979; Graham et al., 1983; Miller, 1987), Mammuthus primigenius (Toepfer, 1957; Ziegler, 2001), Mammuthus columbi (HSM collections), Mammuthus trogontherii (Tong, 2012), and Elephas maximus (LACM collections).
Specimen AZMNH p7487 consists of partial left and right gomphotheriid manuses. These manuses described here were first mentioned in Skaff (2001) who listed, but did not describe the elements recovered. The carpals are referred to by name and for the first use the corresponding synonymous names are added. The metacarpals are referred to as Mc followed by a number (e.g., Mc3 = metacarpal of the third digit). The following measurements are used for the carpals: the greatest proximal-distal height, greatest dorsal-volar depth (D-V), and greatest transverse width, as well as available measurements of the articular facets. For the metacarpals and phalanges the following measurements are used: greatest proximal-distal length (height), proximal dorsal-volar depth, proximal transverse width, distal dorsal-volar depth, distal transverse width, and minimum transverse width (Appendix 1). The identification of the carpals, metacarpals, and phalanges was based on their position when discovered, relative size, and comparisons to other proboscidean specimens from the literature and in museum collections. When possible, the size and morphology of specific elements were compared to the same elements of other extinct and extant proboscideans including Gomphotherium steinheimense (Göhlich, 1998), Haplomastodon chimborazi (Ferretti, 2010), Rhynchotherium falconeri (Frick, 1933; Pasenko, 2007, 2011), Stegomastodon primitivus (Alberdi et al., 2009; Lucas et al., 2011), Mammut americanum (Olsen, 1979; Graham et al., 1983; Miller, 1987), Mammuthus primigenius (Toepfer, 1957; Ziegler, 2001), Mammuthus columbi (HSM collections), Mammuthus trogontherii (Tong, 2012), and Elephas maximus (LACM collections).
SYSTEMATIC PALEONTOLOGY
Class MAMMALIA Linnaeus, 1758
Order PROBOSCIDEA Illiger, 1811
Family GOMPHOTHERIIDAE Hay, 1922
 Description. The two manuses were found together, disarticulated, but with constituent elements closely associated. The right manus is more complete and includes the cuneiform, lunar, scaphoid, unciform, magnum, complete Mc1, several incomplete metacarpals, and complete series of phalanges for the 2nd, 3rd, and 4th digits. The left manus consists of the unciform, portions of three metacarpals, and complete series of phalanges for the 2nd, 3rd, and 4th digits. Measurements of the manuses are presented in Appendix 2.The right cuneiform (pyramidal) (Figure 2.1-3) is missing the distally-oriented portion, volar to the facet for the pisiform, which would have articulated with the Mc5. The facets for the ulna and unciform are preserved. The overall proximal outline of the cuneiform is convex, but the articular facet for the ulna is slightly depressed and roughly triangular in shape. This is similar to other proboscideans. The articular facet for the pisiform is located volarly in a slightly downward proximo-volar angle and is wider at the volar end. The distal articular facet on the cuneiform, for the unciform, is undulating, slightly concave and semi-triangular in shape. The articular facet for the Mc5 is partially preserved and overlaps the distal-lateral surfaces near the volar end. The articular facet for the lunar is narrow at the margins, but increases in height toward the center. The greatest height and depth of the cuneiform is 53.5 mm and 86.4+ mm respectively. The greatest width of the articular facet for the ulna is 92.4 mm.
Description. The two manuses were found together, disarticulated, but with constituent elements closely associated. The right manus is more complete and includes the cuneiform, lunar, scaphoid, unciform, magnum, complete Mc1, several incomplete metacarpals, and complete series of phalanges for the 2nd, 3rd, and 4th digits. The left manus consists of the unciform, portions of three metacarpals, and complete series of phalanges for the 2nd, 3rd, and 4th digits. Measurements of the manuses are presented in Appendix 2.The right cuneiform (pyramidal) (Figure 2.1-3) is missing the distally-oriented portion, volar to the facet for the pisiform, which would have articulated with the Mc5. The facets for the ulna and unciform are preserved. The overall proximal outline of the cuneiform is convex, but the articular facet for the ulna is slightly depressed and roughly triangular in shape. This is similar to other proboscideans. The articular facet for the pisiform is located volarly in a slightly downward proximo-volar angle and is wider at the volar end. The distal articular facet on the cuneiform, for the unciform, is undulating, slightly concave and semi-triangular in shape. The articular facet for the Mc5 is partially preserved and overlaps the distal-lateral surfaces near the volar end. The articular facet for the lunar is narrow at the margins, but increases in height toward the center. The greatest height and depth of the cuneiform is 53.5 mm and 86.4+ mm respectively. The greatest width of the articular facet for the ulna is 92.4 mm.
The right lunar (lunatum) is fairly complete, missing only small portions of the medial and lateral edges (Figure 2.4-6). In proximal view, the lunar is roughly triangular in shape with the medial and lateral sides narrowing as they approach the volar end. The overall shape of the lunar is similar to other proboscideans. However, the narrowing towards the volar end is more pronounced in a specimen of Haplomastodon chimborazi (Ferretti, 2010), which than slightly expands. The lateral side has a pronounced concave outline for articulation with the cuneiform. The large articular facet for the radius is concave and semi-crescent in shape. This facet is separated from the smaller facet for the ulna by a raised area. The facet for the ulna is located proximo-laterally and is semi-oval in shape. The articular facet for the cuneiform is present on the lateral side as a thin, irregular-shaped facet that follows the curvature of the distal concave surface. The medial side of the lunar has two articular facets for the scaphoid; one proximally and the other distally. There is a small, flat articular surface on the distal end of the lunar, near the dorsal-medial border. It is probable this articular surface is for the trapezoid. The dorsal portion of the distal surface is mostly flat, but the volar portion is slightly concave. This articulating surface corresponds with the undulating proximal surface for the magnum. The proximal articular facet for the scaphoid is curved and contained within a slightly depressed area on the medial side. The distal facet for the scaphoid is a thin undulating facet that is widest about midway. The greatest height and depth of the lunar are 60.3 mm and 99.2 mm.
The right scaphoid (naviculare) is mostly complete missing only some of the edges around the articular facets (Figure 2.7-10). The scaphoid is proximo-distally compressed, which is similar to that in other gomphotheriids such as Gomphotherium steinheimense, Haplomastodon chimborazi, and Stegomastodon primitivus, as well as the mastodont Mammut americanum, and contrasts with the extended scaphoids of Mammuthus primigenius, Mammuthus columbi, and Elephas maximus. The scaphoids of H. chimborazi (Ferretti, 2010) and S. primitivus (Lucas et al., 2011) are even more compressed than AZMNH p7487. The scaphoid of AZMNH p7487 only slightly decreases in dorsal-volar depth, from the distal to proximal ends, compared to those of M. primigenius, M. columbi and E, maximus, which have a considerable decrease in depth. Other specimens of gomphotheriids such as H. chimborazi, S. primitivus, and G. steinheimense also show a more uniform dorsal-volar depth for the scaphoid. As shown from the medial and lateral aspects, the dorsal and volar borders of the scaphoid are curved, creating a concave dorsal border and a convex volar border. Although M. primigenius and M. columbi have volar borders that are convex, this is the result of a mostly straight volar border with one small angle near the distal end. The scaphoid of AZMNH p7487 is not angled dorsally, as in mammoths, but instead curves dorsally from the distal to the proximal end. The curved condition of the volar border of the scaphoid was also shown for specimens of G. steinheimense, H. chimborazi, and S. primitivus. The distal edge of the scaphoid's surface is relatively flat throughout most of its length. This is noticed in other gomphotheriids as well. In mammoths and mastodonts, the distal end begins to ascend as it approaches the volar border. The lateral side of the scaphoid displays two articular facets on the distal end. A dorso-lateral facet that is semi-oval in shape for articulation with the lunar, and a second, more volar facet, for articulation with the magnum. The articular facet for the magnum is semi-oval in shape. This facet extends around the distal end where it joins with the articular facet for the trapezoid. The medial side of the scaphoid has a rough surface and a small depression close to the distal end that encompasses the dorsal half. On the proximal end, the scaphoid has a large articular facet for the radial styloid process. This facet is semi-oval in shape, concave, and joins with the upper articular facet for the lunar that is located on the lateral side. However, this facet for the lunar is not completely preserved. The articular facets on the distal end, for the trapezoid and trapezium, are narrow, with the facet for the trapezium being semi-rectangular in shape, and that of the trapezoid being semi-triangular in shape.
The right unciform (hamatum) is fairly complete missing only portions of the medial and lateral edges (Figure 2.11-13). The unciform narrows volarly. For Haplomastodon chimborazi the unciform does not narrow as much volarly, but does display a slight restriction about mid-way between the dorsal and volar ends. This restriction is not evident in AZMNH p7487. The large proximal articular facet for the cuneiform is undulating and semi-triangular in shape. Three distal articular facets are present. The largest, central one, of these facets is for articulation with the Mc4, is solely on the distal surface, and is triangular in shape. The medial and lateral facets, for articulation with the Mc3 and Mc5, are narrow, semi-rectangular in shape, and continue onto the respective medial and lateral sides. The articular facet for the Mc3 is wider. There are two articular facets for the magnum on the medial side: one proximal and the other distal. The proximal facet is irregular in shape having a greater height dorsally that lessens about midway, and then increases again at the volar end. The distal facet for the magnum is narrow and irregular in shape. This configuration of facets mirrors that of the corresponding facets on the magnum. The greatest height of the unciform is 86.5 mm. The left unciform is incomplete, missing a portion of the volar end. The distal facets of the left unciform for the Mc3, Mc4, and Mc5 are present as well as the proximal facet for the cuneiform.
The right magnum (capitatum) is similar to those of most other proboscideans (Figure 2.14-16). However, for the magnum of H. chimborazi the proximal-distal height is significantly less at the volar end than the dorsal. For AZMNH p7487 the height is almost the same at the dorsal and volar ends. There are two articular facets for the unciform, on the lateral side. The proximal articular facet, for the unciform, is larger at the dorsal and volar ends than in the middle. This is similar to the associated articular facets on the unciform for the magnum. The articular facet for the trapezoid, on the medial side, is irregular in shape beginning with a semi-square shaped surface at the dorsal end with a small extension of this facet that continues, volarly, along the distal end. This portion of the articular facet for the trapezoid partially continues along the distal surface where it joins with the articular facet for the Mc2. A portion of the magnum is not preserved at the proximo-volar end. The articular facet for the lunar is smooth and undulating. The distal surface is smooth and slightly undulating for its articulation with the Mc3. The width of the magnum could not be accurately measured since part of the medial side is incomplete. The preserved width is 64.4+ mm. The greatest height of the magnum is 84.9 mm.
 The right Mc1 (Figure 3.1.1-2) is complete, but the remaining metacarpals (Mc2, Mc3, and Mc4) of the right manus are missing some portions of their proximal ends. The Mc1 is similar to other proboscideans. An Mc1 previously described for Rhynchotherium falconeri from 111 Ranch, Arizona (UALP 23404, now housed at AZMNH) and R. edense from California (F:AM 18225), are both smaller. The Mc1 reported here has a greatest length of 57 mm; while that of R. falconeri (UALP 23404) has a greatest length of 132 mm (Pasenko, 2007) and that of R. edense (AMNH F:AM 18225) has a length of 70 mm (Frick, 1933). The Mc1 has a slightly concave proximal articular surface for the trapezium and a small bulge on the dorsal surface at the distal end. The proximal end of the Mc1 extends volarly where it would articulate with the prepollex, but this articular surface appears to be smaller than that of Elephas maximus (Hutchinson et al., 2011; personal examination).The Mc2 (Figure 3.2), Mc3 (Figure 3.3), Mc4 (Figure 3.4), and Mc5 (Figure 3.5) are represented from the left manus and all are missing varying portions of their proximal surfaces. The lateral facet for the Mc4 is preserved on the left Mc3. The greatest length of the Mc5 is 82 mm compared to 90 mm reported for R. edense (Frick, 1933). Metacarpals 2 through 4 are wider medio-laterally (transverse width) than they are craniocaudally (dorsal-volar depth).All three phalanges of the 2nd, 3rd, and 4th digits, for the right and left manuses, are present. The phalanges of the 2nd, 3rd, and 4th digits of the left manus are shown in Figure 3.2-4. In general, the first phalanges are almost square-shaped with the proximal and distal articular facets, for the metacarpals and second phalanges, being smooth and slightly concave. This is also true for the second phalanges. However, the concavity on the distal surface of the second phalanges is greater and coincides with the more rounded proximal ends of the third phalanges. For mammoths, phalanges 1 and 2 appear to have more of a medio-laterally constriction about mid-way between the proximal and distal ends. For some of the phalanges of AZMNH p7487, this constriction is less evident or not present at all. For AZMNH p7487, the proximal-distal heights (length) of the second phalanges are only slightly lesser than those of the first phalanges.
The right Mc1 (Figure 3.1.1-2) is complete, but the remaining metacarpals (Mc2, Mc3, and Mc4) of the right manus are missing some portions of their proximal ends. The Mc1 is similar to other proboscideans. An Mc1 previously described for Rhynchotherium falconeri from 111 Ranch, Arizona (UALP 23404, now housed at AZMNH) and R. edense from California (F:AM 18225), are both smaller. The Mc1 reported here has a greatest length of 57 mm; while that of R. falconeri (UALP 23404) has a greatest length of 132 mm (Pasenko, 2007) and that of R. edense (AMNH F:AM 18225) has a length of 70 mm (Frick, 1933). The Mc1 has a slightly concave proximal articular surface for the trapezium and a small bulge on the dorsal surface at the distal end. The proximal end of the Mc1 extends volarly where it would articulate with the prepollex, but this articular surface appears to be smaller than that of Elephas maximus (Hutchinson et al., 2011; personal examination).The Mc2 (Figure 3.2), Mc3 (Figure 3.3), Mc4 (Figure 3.4), and Mc5 (Figure 3.5) are represented from the left manus and all are missing varying portions of their proximal surfaces. The lateral facet for the Mc4 is preserved on the left Mc3. The greatest length of the Mc5 is 82 mm compared to 90 mm reported for R. edense (Frick, 1933). Metacarpals 2 through 4 are wider medio-laterally (transverse width) than they are craniocaudally (dorsal-volar depth).All three phalanges of the 2nd, 3rd, and 4th digits, for the right and left manuses, are present. The phalanges of the 2nd, 3rd, and 4th digits of the left manus are shown in Figure 3.2-4. In general, the first phalanges are almost square-shaped with the proximal and distal articular facets, for the metacarpals and second phalanges, being smooth and slightly concave. This is also true for the second phalanges. However, the concavity on the distal surface of the second phalanges is greater and coincides with the more rounded proximal ends of the third phalanges. For mammoths, phalanges 1 and 2 appear to have more of a medio-laterally constriction about mid-way between the proximal and distal ends. For some of the phalanges of AZMNH p7487, this constriction is less evident or not present at all. For AZMNH p7487, the proximal-distal heights (length) of the second phalanges are only slightly lesser than those of the first phalanges.
Discussion. For AZMNH p7487, the magnum and unciform, of the distal carpal row, have a greater height than those of the lunar and cuneiform. The carpals of AZMNH p7487 represent the serial arrangement previously discussed for proboscideans (Lister, 1996; Palombo and Villa, 2003). In the serial arrangement the lunar is not wider than the magnum. Although the medial side of the magnum is incomplete and therefore unknown if the width matched that of the lunar, the scaphoid does have an articular facet for the magnum. There is also a small articular surface on the lunar, for the trapezoid. The distal end of the scaphoid for AZMNH p7487 is more level across its entire length, as noticed in other gomphotheriid specimens. This contrasts to mammoths in which the distal end of the scaphoid rises volarly.The proboscidean manus, during stance, is oriented in a semi-digitigrade position as a result of resting on a fibrous cushion. This orientation of the manus, in Loxodonta africana and Elephas maximus includes a nearly vertical carpal-metacarpal arrangement with a scaphoid that is inclined volarly, as it articulates with the trapezoid and trapezium, yet maintains articulation with radius (Weissengruber et al., 2006; personal examinations). For AZMNH p7487, the more level distal surface and curvature of the scaphoid, and the position of the articular facet for the radius, indicates the scaphoid may have been inclined further volarly as it contacted the trapezoid and trapezium, but still allow articulation with the styloid process of the radius, or the articulation with the styloid process was arranged slightly different. If so, the orientation of the manus for AZMNH p7487 may have been slightly different then that displayed in L. africana and E. maximus, with a carpal-metacarpal arrangement slightly less vertical. It is believed that the semi-digitigrade form of extinct proboscideans varied somewhat from being less vertical in earlier proboscidean forms to more vertical in recent extinct and extant proboscideans (Court, 1994; Hutchinson et al., 2011).
Based on available measurements and comparisons, AZMNH p7487 was a small proboscidean. The small size could indicate a sub-adult, or, because of sexual dimorphism, that AZMNH p7487 was a female. The complete Mc1 and mostly complete Mc5 are smaller than those reported for Rhynchotherium edense (AMNH F:AM 18225), which is believed to be a young female based on incompletely erupted M3s/m3, small tusks, and smaller postcrania (Frick, 1933; Pasenko, 2007). All of the carpals and metacarpals, of AZMNH p7487, are smaller than those described for other gomphotheriids, mammoths, and mastodonts (Appendix 2). Several metacarpals are missing their proximal epiphyses, but this appears to be from breakage rather than non-joining of the diaphyses and epiphyses. The distal epiphyses are completely fused to the diaphyses. The preserved portion of the proximal end on the left Mc3 and the mostly complete Mc1 and Mc5 display no conjoining surface or suture line for fusion. No studies have been performed on the fusion sequence of the epiphyses to the diaphyses for proboscidean metacarpals. Based on the small size of the manuses and the lack of any type of conjoining surface or suture lines on the metacarpals, it is more likely that AZMNH p7487 represents a small female gomphotheriid than a sub-adult.
The morphology of several carpals of AZMNH p7487 is similar to those of other proboscideans, but the compressed, curved shape of the scaphoid compares to other gomphotheriids and contrast with those of elephantids, and mammutids. The earliest reported mammoth in North America occurs at 1.7 mya (Harington, 1978), which is younger than the 111 Ranch Beds. The mammutid Mammuthus americanum is known from several late Blancan and early Pleistocene localities (Saunders, 1996), but no remains of M. americanum, or other mammutid, have been found in the 111 Ranch Beds.The gomphotheriids Stegomastodon and Rhynchotherium are the only confirmed proboscideans found at 111 Ranch (Wood, 1962; Skaff, 2001; Pasenko, 2007; AZMNH collections). Cuvieronius is the only other gomphotheriid that temporally correspond with the age of the 111 Ranch Beds, but there are no confirmed specimens of this taxon from the geological unit. Cuvieronius is considered to have derived from Rhynchotherium (Lucas, 2008). Biometrical data for Rhynchotherium and Stegomastodon indicate that the latter was typically larger than the former (Gidley, 1926; Frick, 1933; Miller, 1990; Pasenko, 2007, 2011; Alberdi et al., 2009; Lucas et al., 2011).The elements of the manuses for AZMNH p7487, morphologically, are similar to those shown in Lucas et al. (2011) for S. primitivus except for the scaphoid, which is less curved or rounded than AZMNH p7487 and has a more blocky shape with a broader distal end that tapers proximally. A scaphoid of Haplomastodon chimborazi also shows a blockier, less curved outline than that of AZMNH p7487 (Ferretti, 2010). In addition, the lunar of H. chimborazi has a slightly more expanded volar end.Since no other elements were found associated with the manus, AZMNH p7487 can only presently be identified to the family Gomphotheriidae.
CONCLUSIONS
The manus described here expands our knowledge of these elements for gomphotheriids in North America and hopefully can be used as a basis for further studies. The manuses, described here, are small compared to other extinct and extant proboscideans, and probably represent a young female gomphotheriid. Comparisons to other extinct proboscideans indicate some morphological differences arenoticeable, but more comparisons are needed. Further studies on these elements may provide diagnostic attributes for late Cenozoic gomphotheriids in North America that helps to distinguish between gomphotheriids such as Stegomastodon, Rhynchotherium, and Cuvieronius. At this time, the manuses can only be assigned to the family Gomphotheriidae.
ACKNOWLEDGEMENTS
I would like to acknowledge R. McCord of the Arizona Museum of Natural History for granting me access to the specimens. I also thank L. Thrasher and R. White for their discussions on the 111 Ranch beds. Thanks also goes to J. Saunders and S. Lucas for their comments and suggestions on an earlier draft of this manuscript. I also thank R. Scheetz of Brigham Young University, J. Dines of the Natural History Museum of Los Angeles County, and L. Agenbroad of the Hot Springs Mammoth Site for their assistance in obtaining comparative specimens and/or photographs of specimens from their collections. I would also like to thank the anonymous reviewers for their suggestions and comments.
REFERENCES
Alberdi, M.T., Juárez-Woo, J., Polaco, O.J., and Arroyo-Cabrales, J. 2009. Description of the most complete skeleton of Stegomastodon (Mammalia, Gomphotheriidae) recorded for the Mexican late Pleistocene. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 251/2:239-255.
Court, N. 1994. Limb posture and gait in Numidotherium kohlense, a primitive proboscidean from the Eocene of Algeria. Zoological Journal of the Linnean Society, 111:297-338.
Dickson, J.J. and Izett, G.A. 1981. Fission-track ages of air-fall tuffs in Pliocene basin-fill sediments near 111 Ranch, Graham County, Arizona. Isochron/West, 32:13-15.
Ferretti, M.P. 2008. A review of South American Proboscideans, p. 381-391. In Lucas, S.G., Morgan, G.S., Spielmann, J.S., and Prothero, D.R. (eds.), Neogene Mammals. New Mexico Museum of Natural History and Science Bulletin 44, Albuquerque.
Ferretti, M.P. 2010. Anatomy of Haplomastodonchimborazi (Mammalia, Proboscidea) from the late Pleistocene of Ecuador and its bearing on the phylogeny and systematics of South American Gomphotheres. Geodiversitas, 32(4):663-721.
Frick, C. 1933. New Remains of Trilophodont-tetrabelodont mastodons. Bulletin American Museum of Natural History, 59:505-652.
Galusha, T., Johnson, N.M., Lindsay, E.H., Opdyke, N.N., and Tedford, R.H. 1984. Biostratigraphy and magnetostratigraphy, late Pliocene rocks, 111 Ranch, Arizona. Bulletin Geological Society of America, 95:714-722.
Gidley, J.W. 1926. Fossil Proboscidea and Edentata of the San Pedro Valley, Arizona. United States Geological Survey, Professional Paper, 131-E:83-95.
Göhlich, U.B. 1998. Elephantoidea (Proboscidea, Mammalia) aus dem Mittel und Obermiozän der Oberen Süßwassermolasse Süddeutschlands: Odontologie und Osteologie. Munchner Geowassenschaftliche Abhandlungen, Reihe A, No. 36, Munich.
Graham, R.W., Holman, J.A., and Parmalee, P.W. 1983. Taphonomy and paleoecology of the Christenson Bog Mastodon Bone Bed, Hancock County, Indiana. Illinois State Museum Reports of Investigations, Number 38, Springfield.
Harington, C.R. 1978. Quaternary vertebrate faunas of Canada and Alaska and their suggested chronologic sequence. Syllogeus, Number 15, National Museum of Canada, Ottawa.
Hay, O.P. 1922. Further observations on some extinct elephants. Proceedings of the Biological Society of Washington, 35:97-101.
Houser, B.B. 1990. Late Cenozoic stratigraphy and tectonics of the Safford Basin, southeastern Arizona, p. 20-24. In Gehrels, G.E. and Spencer, J.E. (eds.), Geologic excursions through the Sonoran Desert region, Arizona and Sonora: Arizona Geological Survey of America, Cordilleran Section, 86th Annual Meeting, Tucson, Arizona, Arizona Geological Survey Special Paper 7.
Houser, B.B., Pearthree, P.A., Homburg, J.A., and Thrasher, L.C. 2004. Quaternary stratigraphy and tectonics, and late prehistoric agriculture of the Safford Basin (Gila and San Simon River Valleys), Graham County, Arizona. U.S. Geological Survey and U.S. Department of the Interior Open File Report, 2004-1062.
Hutchinson, J.R., Delmer, C., Miller, C.E., Hildebrandt, T., Pitsillides, A.A., and Boyde, A. 2011. From flat foot to fat foot: Structure, ontogeny, function, and evolution of elephant "sixth toes." Science, 334:1699-1703.
Illiger, C.D. 1811. Prodromus Systematis Mammalium et Avium Additis Terminus Zoographicis Uttriusque Classis. Salfeld, Berlin.
Lindsay, E.H. 1978. Late Cenozoic vertebrate faunas, southeastern Arizona. New Mexico Geological Society Guidebook 29th Field Conference, Land of Cochise, p. 269-275.
Lindsay, E.H. and Tessman, N. 1974. Cenozoic vertebrate faunas in Arizona. Journal of Arizona Academy of Science, 9(1):3-28.
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species cum caracteribus, differentiis, synonymis, locis. Editio decima, reformata, Volume I. Laurentii Salvii, Holmiae.
Lister, A.M. 1996. Evolution and taxonomy of Eurasian mammoths, p. 203-213. In Shoshani, J. and Tassy, P. (eds.), The Proboscidea: Evolution and Palaeoecology of Elephants and Their Relatives. Oxford University Press, New York.
Lucas, S.G. 2008. Cuvieronius (Mammalia, Proboscidea) from the Neogene of Florida, p. 31-38. In Lucas, S.G., Morgan, G.S., Spielmann, J.S., and Prothero, D.R (eds.), Neogene Mammals. New Mexico Museum of Natural History and Science Bulletin 44, Albuquerque.
Lucas, S.G., Aguilar, R.H., and Spielmann, J.A.. 2011. Stegomastodon (Mammalia, Proboscidea) from the Pliocene of Jalisco, Mexico and the species-level taxonomy of Stegomastodon, p. 517-553. In Sullivan, R.M., Lucas, S.G., and Spielmann, J.A. (eds.). Fossil Record 3. New Mexico Museum of Natural History and Science Bulletin 53, Albuquerque.
Miller, W.E. 1987. Mammut americanum, Utah's first record of the American Mastodon. Journal of Paleontology, 61:168-183.
Miller, W.E. 1990. A Rhynchotherium skull and mandible from southeastern Arizona. Geology Studies, 36:57-67.
Morgan, G.S. and White, R.S., Jr. 2005. Miocene and Pliocene vertebrates from Arizona, p. 115-136. In Heckert, A.R. and Lucas, S.G. (eds.), Vertebrate Paleontology in Arizona. New Mexico Museum of Natural History and Science Bulletin Number 29, Albuquerque.
Olsen, S.J. 1979. Osteology for the archaeologist, Number 1,The American Mastodon and the Woolly Mammoth. Papers of the Peabody Museum of Archaeology and Ethnology, 56(3):1-46.
Palombo, M.R. and Villa, P. 2003. Sexually dimorphic characters of Elephas (Palaeoloxodon) antiquus from Grotte Santo Stefano (Viterbo, Central Italy). Deinsea, 9:293-315
Pasenko, M.R. 2007. Rhynchotherium falconeri (Proboscidea, Gomphotheriidae) from 111 Ranch, southeastern Arizona with a discussion of the genus. Journal of Vertebrate Paleontology, 27(1):195-203.
Pasenko, M.R. 2011. Observations of postcranial elements of Rhynchotherium edense (Proboscidea, Mammalia) from the Pliocene Horned Toad Formation, southern California, p. 583-91. In Sullivan, R.M., Lucas, S.G., and Spielmann, J.A. (eds.), Fossil Record 3. New Mexico Museum of Natural History and Science Bulletin 53, Albuquerque.
Pasenko, M.R. 2012. New remains of Rhynchotherium falconeri (Mammalia, Proboscidea) from the earliest Pleistocene 111 Ranch, Arizona, USA with a discussion on sexual dimorhism and paleoenvironment of rhynchotheres. Palaeodiversity, 5:89-97.
Pasenko, M.R. and Lucas, S.G. 2011. A review of gomphotheriid (Proboscidea, Mammalia) remains from the Pliocene Benson and Curtis Ranch Local Faunas, southern Arizona, with a discussion of gomphotheriids in Arizona, p. 592-601. In Sullivan, R.M., Lucas, S.G., and. Spielmann, J.A. (eds.), Fossil Record 3. New Mexico Museum of Natural History and Science Bulletin 53, Albuquerque.
Richter, D.H., Shafiqullah, M., and Lawrence, V.A.. 1981. Geologic map of the Whitlock Mountains and vicinity, Graham County, Arizona. United States Geological Survey Miscellaneous Investigations Series Map I-1302, 1 sheet, scale 1:48,000.
Saunders, J.J. 1996. North American Mammutidae, p. 271-279. In Shoshani, J. and Tassy, P. (eds.), The Proboscidea: Evolution and Palaeoecology of Elephants and Their Relatives. Oxford University Press, New York.
Seff, P. 1960. Preliminary report of the stratigraphy of the 111 Ranch, Graham County, Arizona. Arizona Geological Digest, 3:13.
Seff, P. 1962. Stratigraphic geology and depositional environments of the 111 Ranch Area, Graham County, Arizona. Unpublished PhD thesis, University of Arizona, Tucson.
Skaff, A. 2001. Analyses of proboscideans from the Chandler and 111 Ranch localities, central Arizona. Unpublsihed Master's thesis, Arizona State University, Phoenix.
Thrasher, L. 2007. Fossil tracks and trackways from the Bowie zeolite mines, Graham County, Arizona, p. 269-273. In Lucas, S.G., Spielmann, J.A., and Lockley, M.G. (eds.), Cenozoic Vertebrate Tracks and Traces. New Mexico Museum of Natural History and Science Bulletin 42, Albuquerque.
Tomida, H. 1987. Small mammal fossils and correlations of Continental Deposits, Safford and Duncan Basins, Arizona. National Science Museum, Tokyo.
Toepfer, V. 1957. Die Mammutfunde von Pfännerhall im Geiseltal. Veröffentlichungen des Landesmuseums für Vorgeschicte in Halle, Heft 16:1-58.
Tong, H. 2012. New remains of Mammuthus trogontherii from the early Pleistocene Nihewan beds at Shanshenmiaozui, Hebei. Quaternary International, 255:217-230.
Weissengruber, G.E., Egger, G.F., Hutchinson, J.R., Groenwald, H.B., Elsässer, L., Famini, D., and Forstenpointer, G. 2006. The structure of the cushions in the feet of African elephants (Loxodonta Africana). Journal of Anatomy, 209:781-792.
Wood, P.A. 1962. Pleistocene fauna from the 111 Ranch, Graham County, Arizona. Unpublished PhD. thesis, University of Arizona, Tuscon.
Ziegler, R. 2001. An extraordinary small mammoth (Mammuthus primigenius) from SW Germany. Stuttgarter Beiträge zur Naturkunde Serie B (Geologie und Paläontologie), 300:1-41.
