

 A revision of Heritschioides Yabe, 1950 (Anthozoa, Rugosa), latest Mississippian and earliest Pennsylvanian of western North America
A revision of Heritschioides Yabe, 1950 (Anthozoa, Rugosa), latest Mississippian and earliest Pennsylvanian of western North America
Article number: 17.1.11A
https://doi.org/10.26879/383
Copyright Palaeontological Association, February 2014
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 12 March 2013. Acceptance: 11 February 2014
{flike id=686}
ABSTRACT
New data from a detailed study of the type and topotype collections of the type species of Heritschioides confirm the unique status of the genus as colonial and bearing extra septal lamellae. The associated microfossils establish its age as late Serpukhovian to early Bashkirian. The close connection of the cardinal septum to the median lamella and the axial structure points to the family Aulophyllidae. However, the inconsistent role of the protosepta in the formation of the median lamella is unique for Heritschioides. This feature and the colonial growth form allow its assignment to a separate subfamily, the Heritschioidinae Sando, 1985, which is closely related to the subfamily Aulophyllinae. So far, the subfamily Heritschioidinae is known to occur only in rocks along the western margin of North America.
Jerzy Fedorowski. Institute of Geology, Adam Mickiewicz University, Maków Polnych 16, Pl-61-606, Poznań, Poland. jerzy@amu.edu.pl
E. Wayne Bamber. Geological Survey of Canada (Calgary), 3303-33rd Street N. W., Calgary, Alberta T2L 2A7, Canada. wabamber@nrcan.gc.ca
Calvin H. Stevens. Department of Geology, San Jose State University, San Jose, California 95192, USA. calvin.stevens@sjsu.edu
Keywords: Late Serpukhovian-early Bashkirian; colonial coral; type specimens; western North America; allochthonous terranes
Final citation: Fedorowski, Jerzy, Bamber, E. Wayne, and Stevens, Calvin H. 2014. A revision of Heritschioides Yabe, 1950 (Anthozoa, Rugosa), latest Mississippian and earliest Pennsylvanian of western North America. Palaeontologia Electronica Vol. 17, Issue 1;11A; 20p. https://doi.org/10.26879/383
palaeo-electronica.org/content/2014/686-the-coral-heritschioides
INTRODUCTION
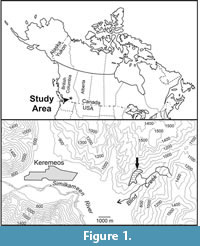 Corals here assigned to the genus Heritschioideswere first described by Smith (1935) from small fragments of colonies collected from the Blind Creek Limestone, which crops out in an isolated structural block on a ridge northwest of Blind Creek, approximately 6.5 kilometers east of Keremeos in southern British Columbia, Canada (48°12'20"N, 119°43'20"W; Figure 1). On the basis of complex axial structures similar to those in Waagenophyllum Hayasaka, 1924, combined with other unique morphological features in the type specimens, Smith (1935) introduced the new species Waagenophyllum columbicum. That identification was questioned at the generic level by Yabe (1950), who introduced the new generic name Heritschioides for Smith's (1935) species and suggested Corwenia Smith and Ryder, 1926 as a possibly related genus. Yabe's (1950) interpretation has found general acceptance, although the family status of Heritschioides and its taxonomic relationships have remained controversial. Wilson and Langenheim (1962) misinterpreted the tabulae in Heritschioides as clinotabellae, rather than biform tabulae in position I of Sutherland (1965), and placed the genus in the family Waagenophyllidae. They referred to Heritschioides as cerioid in their text (p. 509), but included fasciculate species within it. Also, they erroneously attributed the authorship of the family Waagenophyllidae to Hudson (1958) who only elevated its status from the subfamily level introduced by Wang (1950). Heritschioides was included by Minato and Kato (1965) in their new family Durhaminidae, a concept followed by Stevens (1967) and Hill (1981). Wilson (1980), who first revised the type collection of "Waagenophyllum" columbicum, did not assign it to any family (p. 89, family uncertain), and later avoided that question by using the term "Fasciculate Rugosa" (Wilson, 1982, p. 25) and placing species of Heritschioides immediately after those of Durhamina.
Corals here assigned to the genus Heritschioideswere first described by Smith (1935) from small fragments of colonies collected from the Blind Creek Limestone, which crops out in an isolated structural block on a ridge northwest of Blind Creek, approximately 6.5 kilometers east of Keremeos in southern British Columbia, Canada (48°12'20"N, 119°43'20"W; Figure 1). On the basis of complex axial structures similar to those in Waagenophyllum Hayasaka, 1924, combined with other unique morphological features in the type specimens, Smith (1935) introduced the new species Waagenophyllum columbicum. That identification was questioned at the generic level by Yabe (1950), who introduced the new generic name Heritschioides for Smith's (1935) species and suggested Corwenia Smith and Ryder, 1926 as a possibly related genus. Yabe's (1950) interpretation has found general acceptance, although the family status of Heritschioides and its taxonomic relationships have remained controversial. Wilson and Langenheim (1962) misinterpreted the tabulae in Heritschioides as clinotabellae, rather than biform tabulae in position I of Sutherland (1965), and placed the genus in the family Waagenophyllidae. They referred to Heritschioides as cerioid in their text (p. 509), but included fasciculate species within it. Also, they erroneously attributed the authorship of the family Waagenophyllidae to Hudson (1958) who only elevated its status from the subfamily level introduced by Wang (1950). Heritschioides was included by Minato and Kato (1965) in their new family Durhaminidae, a concept followed by Stevens (1967) and Hill (1981). Wilson (1980), who first revised the type collection of "Waagenophyllum" columbicum, did not assign it to any family (p. 89, family uncertain), and later avoided that question by using the term "Fasciculate Rugosa" (Wilson, 1982, p. 25) and placing species of Heritschioides immediately after those of Durhamina.
The family Heritschioididae was erected by Sando (1985). Subsequently, Wilson (1994) placed his newly described species of Heritschioides from California in that family. This assignment was later followed by Stevens and Rycerski (1989) for species from Permian strata of the accreted Stikine terrane of British Columbia, Canada, and by Kossovaya (1997) for "Corwenia" densicolumella Dobrolyubova, 1936 and Heritschioides cf. carneyi Wilson, 1982 from Cisuralian strata of the northern Urals. Here it should be noted that all species included in Heritschioides by authors such as Wilson and Langenheim (1962), Stevens (1967), Rowett (1969), Wilson (1982, 1994), and Kossovaya (1997) have already been transferred to other genera by Fedorowski et al. (2007), thereby establishing that Heritschioides is not present in Permian deposits and is perhaps restricted to Serpukhovian (?) to Moscovian strata.
Lin et al. (1995, p. 528) placed Heritschioides in the subfamily Aulophyllinae Dybowski, 1873, thus returning to the concept of Yabe (1950). That position and other aspects of the taxonomy of Heritschioides-like corals were briefly discussed by Fedorowski et al. (2007, p. 100-101) and are more widely treated below in the remarks on the family Aulophyllidae. In the most recent paper dealing with Heritschioides, Kawamura and Stevens (2012) accepted an independent status for the family Heritschioididae and placed it within the suborder Aulophyllina Hill, 1981.
There also has been controversy concerning the stratigraphic position of the type locality, which has been assigned to ages ranging from Late Mississippian to Early Permian (Smith, 1935, p. 38; Barnes and Ross, 1975, p. 1933; Wilson, 1980, p. 90-91; Danner, 1997, p. 24; Danner et al., 1999, appendix 2). Wilson (1980) accepted a Permian age, although he mentioned (p. 90) doubts raised in a written communication by Danner, who pointed to the occurrence of the foraminifer Endothyra at the type locality indicating a Late Mississippian or Early Pennsylvanian age. However, Wilson (1980, p. 90-91) concluded that the type locality may be Lower Permian, because "the 10 other valid taxa of Heritschioides all are from Lower Permian rocks in western North America." In view of the misunderstanding of the taxonomic position of Permian species noted above, that conclusion cannot be correct. Danner (1997) and Danner et al. (1999) later supported his earlier informal position with more foraminiferal data (Danner et al., 1999, appendix 2). He reported the genera Eostaffella, Endothyra, Plectogyra and Tetrataxis from the type locality and suggested a late Serpukhovian or early Bashkirian age for the Blind Creek Limestone. His suggestion was confirmed by B.L. Mamet and S. Pinard (personal commun., 1998; see Fedorowski et al. 2007, p. 100) and is accepted in the present paper. A new collection of coral colonies made from the type locality in 1998 by one of us (EWB) and several associates (see acknowledgments) also produced Serpukhovian/early Bashkirian foraminifers and conodonts and allowed a new approach to the genus Heritschioides, previously presented in an introductory manner by Fedorowski et al. (2007, p. 79, 100-101).
Kawamura and Stevens (2012) described the new species Heritschioides armstrongi from the lower Atokan (Bashkirian) Baird Formation of the Klamath accreted terrane (northwestern California). That species exhibits some characters of the type and topotype specimens of Heritschioides, as mentioned by its authors, but does not have extra septal lamellae in its axial structure. Such lamellae, however, do occur in an unidentified specimen (Kawamura and Stevens, 2012, p. 519, figure 2.16) from the same area and stratigraphic level. The morphology of the axial structure and its taxonomic value are discussed below in the remarks on the family.
MATERIAL AND METHODS
All thin sections of the type collection were re-studied, but only a few were re-photographed to illustrate special features. The reader is referred to papers by Smith (1935), Wilson (1980) and Fedorowski et al. (2007) for more complete documentation. Unfortunately, the holotype and several paratypes are represented only by thin sections. The existence and location of the originally sectioned specimens have not been documented. One new thin section, prepared from paratype GSC 9061, was illustrated by Fedorowski et. al. (2007, plate 8, figure 5). From the recently collected topotype material, six of the best preserved specimens were selected for thin sectioning and serial peeling in Calgary and Poznań, producing 26 transverse and well oriented longitudinal thin sections, 12 peels of colony surfaces and three serial sections of offsetting corallites with several peels each. Several thin sections were photographed and some were used as the basis for drawings made by the senior author to better demonstrate the earliest morphology of offsets, one of the characters critical for the correct identification of specimens.
All colonies have been recrystallized, dolomitized or silicified, and several corallites have been compressed, crushed or partly destroyed by pressure solution at their peripheries. A detailed study of the macro-morphology was possible despite those diagenetic alterations. The original microstructure of the septa was destroyed in most corallites, but remains recognizable in some. Also, the serial peels are too poor to be illustrated. Thus, only drawings based on serial thin sections of offsets are illustrated.
The type specimens described in this paper are in the type collection of the Geological Survey of Canada, Ottawa.
SYSTEMATIC PALAEONTOLOGY
Subclass RUGOSA Milne-Edwards and Haime, 1850
Order STAURIIDA Verrill, 1865
Suborder AULOPHYLLINA Hill, 1981
Family AULOPHYLLIDAE Dybowski, 1873
Subfamily HERITSCHIOIDINAE Sando, 1985
Emended diagnosis. Colonial, dissepimented Aulophyllidae; offsetting lateral; complex axial column closely connected to cardinal septum; extra septal lamellae and median lamella reduced in some corallites; dissepimentarium interseptal; small lonsdaleoid dissepiments and pseudonaotic septa at maturity; septal flanges may occur; biform tabulae in some septal loculi.
Genera assigned. Copia Vassilyuk and Kozyreva, 1974; Heritschioides Yabe, 1950.
Remarks. The taxonomic rank of the Heritschioididae Sando, 1985 is here reduced to the subfamily level and is included in the family Aulophyllidae. Both of these decisions resulted from a close examination of the offsetting and some other characters, discussed in detail below. Hill (1981) divided the family Aulophyllidae into six subfamilies, comprising mostly solitary and mostly Viséan and Serpukhovian taxa, few of which continue into the Bashkirian. We have not analyzed those families, restricting our remarks to Corwenia Smith and Ryder, 1926 and Copia Vassilyuk and Kosyreva, 1974, which are the only colonial taxa in the family that have an axial column and can be considered as either related to or similar to Heritschioides. Within the family, Corwenia, included by Hill (1981) in the subfamily Dibunophyllinae Wang, 1950, is the only well documented colonial taxon, ranging from the Viséan in Britain into the Moscovian (Kashirskian and Podolskian) in northern Spain (de Groot, 1963; Rodríguez, 1984; Fedorowski, 2004). At the subfamily level Heritschioides is distinguished from Corwenia by variability in the length of the axial septum, the occurrence of extra septal lamellae and by insertion of the cardinal septum as a neoseptum. Therefore, Corwenia should be eliminated as a direct ancestor of Heritschioides.
Copia Vassilyuk and Kozyreva, 1974, from the upper Viséan of the Voronezh Uplift in the European part of the Russian Federation, is another genus to be considered as either similar to or related to Heritschioides. Unfortunately, its septal microstructure and mode of offsetting remain unknown. Offsetting corallites, firmly documenting coloniality, were not illustrated by Vassilyuk and Kosyreva (1974), and closely packed corallites do not necessarily indicate a colonial growth form. However, V.V. Ogar (Kiev University, Ukraine, personal commun., 2012) answered the question asked by one of us (JF) as follows: "I have no doubt that Protodurhamina and Copia are fully colonial corals."
The characters of Copia closely resembling those in Heritschioides are: 1) a continuous axial column shown in longitudinal sections and the possible occurrence of extra septal lamellae in the axial structure as seen in transverse sections, 2) a thin, monoseptal median lamella elongated toward and commonly united with the cardinal septum, and 3) a complex dissepimentarium with pseudonaotic structures occurring at its periphery and small lonsdaleoid vesicles in advanced maturity of the holotype. Features distinguishing Copia are: 1) its easily recognizable cardinal fossula, 2) longitudinal splitting of the inner parts of its major septa in advanced maturity and 3) more pronounced thickening of the inner margins of its major septa in the cardinal quadrants. We had no access to the Russian material for checking, but we accept a colonial growth form for Copia, consider it related to Heritschioides and include it in the subfamily Heritschioidinae.
The overwhelming majority of taxa indisputably belonging to the family Aulophyllidae are solitary taxa described from the Western and Eastern European Coral Provinces, North Africa and China. They are absent from Australia and their occurrence is mostly doubtful in the USA in the Southeastern Province of Sando et al. (1975). Specimens included by Moore and Jeffords (1945) in Rhodophyllum Thomson, 1874 and Pseudozaphrentoides Stuckenberg, 1904, in Dibunophyllum Thomson and Nicholson, 1876 by Newell (1935), Rowett and Sutherland (1964) and Cocke (1969, 1970), and in Koninckophyllum Thomson and Nicholson, 1876 by Rowett and Sutherland (1964) and Cocke (1970) need revision. The microstructure of septa remains unknown in all of those taxa. The early growth stages were investigated by Cocke (1970) in two species included by him in Dibunophyllum, but he did not illustrate the formation of the axial structure and its connection to either of the protosepta. That character, important for correct identification, is also unclear in the mature growth stages of corallites illustrated by Cocke (1969, 1970) and other authors listed above. The median lamella is connected to both protosepta in most specimens illustrated by those authors or remains connected to the counter rather than to the cardinal septum in some. The cardinal septum is commonly not shortened, and the cardinal tabular fossula is either absent or almost unrecognizable. Some of those characters occur in typical European Aulophyllidae (e.g., the long-lasting axial septum in Corwenia), but the occurrence of all of those characters in North American "dibunophyllids" do not fit any of the subfamilies of the Aulophyllidae present in Europe, Asia and North Africa. Thus, the relationship of North American taxa discussed above to the members of the Aulophyllidae, if it exists, is not closer than at the family level. That does not apply to corals in Nova Scotia (Bell, 1929; Lewis, 1935), which belong to the Western European Province (Fedorowski, 1981). Also, recent studies have documented the occurrence of some solitary aulophyllids in the middle Bashkirian of the Sverdrup Basin (Fedorowski and Bamber, 2012; Fedorowski et al., 2012) and in cratonal Serpukhovian strata of the southern Canadian Rocky Mountains (study in progress).
Fasciculate colonies from the Middle Pennsylvanian (Atokan) of the eastern Brooks Range, Alaska, USA, identified by Armstrong (1972) as the new species Corwenia jagoensis, have a few characters in common with "Lonsdaleia" rugosa McCoy, 1849, the type species of Corwenia. Some of Armstrong's colonies may belong to Corwenia, some to Heritschioides and others to a third genus, perhaps Heintzella Fedorowski, 1967. Also, there are several other uncertainties in Armstrong's (1972) descriptions and illustrations that require clarification.
The presence of a complex, continuous axial column, lateral extension of the axial structure towards the cardinal fossula and the development of pseudonaotic septa confirm to a large extent the generic identification of Heritschioides armstrongi Kawamura and Stevens, 2012. That assignment is indirectly supported by an unidentified specimen of Kawamura and Stevens (2012, text-figure 2.16) having extra septal lamellae in its axial structure. Strong variation in components of the axial structure in the type and topotype specimens of Heritschioides columbicum and undescribed colonies from Kuiu Island, southeastern Alaska, presently under investigation, suggests that similar variation may occur in H. armstrongi, including the temporary appearance of extra septal lamellae in its axial structure.
No other North American rugose coral genera or species appear to be closely related to Heritschioides, although the continuous axial column and extra septal lamellae, typical for that genus, are found in the axial structures of the Lower Permian genera Sandolasma and Shastalasma, both of Fedorowski et al. (2007). The latter genera, however, have early growth stages typical for the families Durhaminidae Minato and Kato, 1965 and Kleopatrinidae Fedorowski et al., 2007, respectively, indicating that they are morphotypes structurally resembling Heritschioides, but are not related to it. Thus, neither ancestors nor descendents of Heritschioides can be presently pointed out. Also, as discussed above, true representatives of Heritschioides are absent from the Lower Permian and may be restricted to Serpukhovian- Moscovian strata mostly in allochthonous oceanic platforms later accreted to the western North American cratonic margin.
Genus HERITSCHIOIDES Yabe, 1950
Type Species. Waagenophyllum columbicum Smith, 1935, p. 38, pl. 8, fig. 9; pl. 9, figs. 1-10.
Species included. Waagenophyllum columbicum Smith, 1935; Heritschioides armstrongi Kawamura and Stevens, 2012.
Emended Diagnosis. Fasciculate Heritschioidinae with median lamella commonly reduced at advanced maturity; pseudonaotic septa common, carinate septa rarely present at advanced maturity; dissepimentarium complex with small lonsdaleoid dissepiments.
Remarks. As for the subfamily.
Heritschioides columbicum (Smith, 1935)
Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9
1935 Waagenophyllum columbicum Smith, p. 38, pl. 8, fig. 9; pl. 9, figs. 1-10.
1941 Heritschia columbica (Smith); Moore and Jeffords, pl. 4, fig. 4.
non 1944 Waagenophyllum columbicum Stanley Smith; Heritsch, p. 54, pl. 3, figs. 4-7.
1950 Heritschioides columbicum (Smith); Yabe, p. 75.
1965 Heritschioides columbicum (Smith); Minato and Kato, p.53.
1980 Heritschioides columbicum (Smith); Wilson, p. 90, pl. 1, 2.
2007 Heritschioides columbicum (Smith, 1935); Fedorowski, Bamber and Stevens,
pl. 8, fig. 5; pl. 9, figs. 1-4.
 Type Specimen. Holotype GSC 9059.
Type Specimen. Holotype GSC 9059.
Type Locality and Age. Blind Creek Limestone, near Keremeos, southern British Columbia, Canada (Figure 1). Late Serpukhovian/early Bashkirian.
Emended Diagnosis. Heritschioides with number of major septa to mean corallite diameter values 32:12.5 mm to 36:16.5 mm; major septa closely approach or penetrate axial structure; minor septa extend well into tabularium; both cycles of septa may bear flanges and are pseudonaotic at maturity; cardinal septum shortened in some mature corallites; dissepiments mostly interseptal, but small lonsdaleoid dissepiments common; elongate tabellae in axial column closely spaced and very steeply dipping; periaxial tabellae mostly vesiculate; complete tabulae rare.
Material. Holotype GSC 9059: Four transverse and one longitudinal thin sections without remnants of colony (Figure 2.1-4) and paratype GSC 9060 - three transverse and one longitudinal sections without remnants of colony (Figure 2.5-7) all illustrated by Smith (1935), Wilson (1980) and herein (Figure 2.1-7). One holotype transverse thin section also illustrated by Fedorowski et al. (2007). In addition the following specimens were investigated: Paratype GSC 9061 - small fragment of a colony, with mature, mostly incomplete, compressed corallites, two transverse acetate peels, one transverse thin section illustrated by Fedorowski et al. (2007). Topotype colonies: GSC 124100 – 21 fragments, seven transverse and one longitudinal thin sections, 46 transverse acetate peels; topotype GSC 136092, seven fragments, one transverse and one longitudinal section; topotype GSC 136093, 2 small fragments (not thin sectioned or peeled); topotype GSC 136094, 11 fragments, seven transverse and one longitudinal thin sections, seven transverse acetate peels; topotype GSC 136095, 11 fragments, three transverse and two longitudinal thin sections, four transverse acetate peels; topotype GSC 136096, 16 fragments, two transverse and five longitudinal thin sections, and two transverse acetate peels.
Remarks on Types and Additional Description
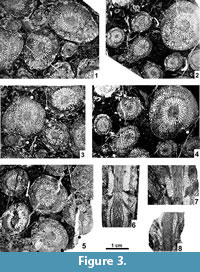 The original type material was described and illustrated by Smith (1935) and thoroughly revised by Wilson (1980), who also offered a supplementary description. Thus, only details of types that were either omitted or misinterpreted by those two authors are added here. Supplementary data were obtained from the new topotype collection. All existing thin sections of the original collection were illustrated by Smith (1935) and Wilson (1980), and only those needed to support our descriptions are re-illustrated. These include supplementary photographs of the holotype, paratypes and topotypes, illustrating their general morphology (Figure 2.1-7, Figure 3.1-8). Details of morphology, blastogeny, microstructure, diagenesis and drawings of two short series showing offsetting corallites are illustrated separately.
The original type material was described and illustrated by Smith (1935) and thoroughly revised by Wilson (1980), who also offered a supplementary description. Thus, only details of types that were either omitted or misinterpreted by those two authors are added here. Supplementary data were obtained from the new topotype collection. All existing thin sections of the original collection were illustrated by Smith (1935) and Wilson (1980), and only those needed to support our descriptions are re-illustrated. These include supplementary photographs of the holotype, paratypes and topotypes, illustrating their general morphology (Figure 2.1-7, Figure 3.1-8). Details of morphology, blastogeny, microstructure, diagenesis and drawings of two short series showing offsetting corallites are illustrated separately.
Major and Minor Septa. Both cycles of septa are mainly indistinguishable from one another in the dissepimentarium. The minors invariably penetrate the tabularium, but are restricted to its periphery, where their inner margins become thickened. In the immature growth stage all septa are slightly wavy along their length, and their thickness is either equal to or slightly less than in the tabularium (Figure 4.1-5). Within the tabularium, the septa become thicker during corallite growth, but their thickness remains uniform within any growth stage (Figure 2.1-4, 2.7, Figure 3.1-5, Figure 4.1-5). In the dissepimentarium the septa may be variously modified, generally dependent on growth stage. However, most corallites of the holotype colony have simple dissepimentaria and unchanged septa up to a comparatively large diameter, at which stage a complex axial structure develops (Figure 2.1-4).
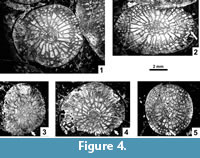 Modification of the septa within the dissepimentarium began with increased lateral distortion (Figure 5.6, 5.8-9), resulting in: a) the break-up of some of their middle parts into small lonsdaleoid dissepiments (Figure 5.1-2, 5.5), and b) the formation of incipient pseudo-naotic structures expressed as slight fragmentation and thickening of their peripheral margins. The corallites preserved in the holotype colony did not advance beyond that stage (Figure 5.8-9). This may indicate that fully mature parts of the holotype colony were not collected. However, the corallites in the holotype developed offsets and therefore should be considered mature. Due to the limited nature of the material available, we have accepted large intraspecific variability in the morphology of offsetting corallites and in the process of offsetting (see below).
Modification of the septa within the dissepimentarium began with increased lateral distortion (Figure 5.6, 5.8-9), resulting in: a) the break-up of some of their middle parts into small lonsdaleoid dissepiments (Figure 5.1-2, 5.5), and b) the formation of incipient pseudo-naotic structures expressed as slight fragmentation and thickening of their peripheral margins. The corallites preserved in the holotype colony did not advance beyond that stage (Figure 5.8-9). This may indicate that fully mature parts of the holotype colony were not collected. However, the corallites in the holotype developed offsets and therefore should be considered mature. Due to the limited nature of the material available, we have accepted large intraspecific variability in the morphology of offsetting corallites and in the process of offsetting (see below).
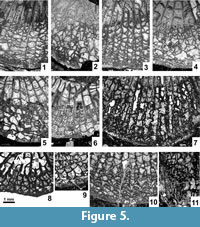 Several corallites in the topotype colonies, morphologically more advanced than the holotype and paratype corallites, developed such strong lateral distortion in their septa that short flanges were formed, i.e., they became carinate (Figure 5.7). Such carination has not been observed either as a long-lasting feature or occurring over the entire transverse section of a given corallite. Thus, it should be considered ephemeral. Its appearance is difficult to explain. The carinate septa may in some instances be interrupted by lonsdaleoid dissepiments (Figure 5.4). Peripheral parts of major and minor septa in the morphologically most advanced corallites of the topotype colonies become fragmented to form naotic and pseudonaotic structures (Figure 5.10). Longitudinal splitting of septa into two forks with dissepiments developed between them (Figure 5.3) should be treated as an introductory step in that process. True naotic structures, i.e., those composed solely of fragments of septa, are absent from the type material, but rare peripheral parts of septa almost reach that stage of fragmentation (Figure 5.10). Pseudonaotic structures, i.e., those combining both fragmented septa and accompanying, small, peripheral dissepiments, are much more common. They vary greatly in shape and content, but occur in all large corallites in the topotype colonies. A longitudinal section (Figure 5.11) exposing both fragmented septa and the dissepiments associated with them confirms the relationship of those two skeletal structures. Corallites of the holotype colony show pseudonaotic structures in a very early stage of development (Figure 5.8-9).
Several corallites in the topotype colonies, morphologically more advanced than the holotype and paratype corallites, developed such strong lateral distortion in their septa that short flanges were formed, i.e., they became carinate (Figure 5.7). Such carination has not been observed either as a long-lasting feature or occurring over the entire transverse section of a given corallite. Thus, it should be considered ephemeral. Its appearance is difficult to explain. The carinate septa may in some instances be interrupted by lonsdaleoid dissepiments (Figure 5.4). Peripheral parts of major and minor septa in the morphologically most advanced corallites of the topotype colonies become fragmented to form naotic and pseudonaotic structures (Figure 5.10). Longitudinal splitting of septa into two forks with dissepiments developed between them (Figure 5.3) should be treated as an introductory step in that process. True naotic structures, i.e., those composed solely of fragments of septa, are absent from the type material, but rare peripheral parts of septa almost reach that stage of fragmentation (Figure 5.10). Pseudonaotic structures, i.e., those combining both fragmented septa and accompanying, small, peripheral dissepiments, are much more common. They vary greatly in shape and content, but occur in all large corallites in the topotype colonies. A longitudinal section (Figure 5.11) exposing both fragmented septa and the dissepiments associated with them confirms the relationship of those two skeletal structures. Corallites of the holotype colony show pseudonaotic structures in a very early stage of development (Figure 5.8-9).
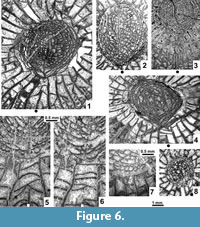 Length of the cardinal septum. Smith (1935) drew special attention to the morphology seen in longitudinal section, but did not mention the cardinal septum, whereas Wilson (1980) characterized it as "somewhat shorter than other major septa." Fedorowski et al. (2007, p. 100) were of a different opinion, pointing to "short plates attached to intercepts of tabulae between the cardinal septum and the median lamella." Those short plates are fragments of the cardinal septum cut between individual tabulae within the shallow cardinal fossula. Such an image resulted from the low elevation of the cardinal septum above the tabulae as it extends to the median lamella along the cardinal fossula floor. We now confirm that the cardinal septum is permanently elongate in all type corallites preserved adequately to demonstrate that character (Figure 2.1-4, 2.7) and in most young and medium-sized corallites within individual topotype colonies. However, several exceptions occur among immature corallites in those colonies. Their cardinal septa may either be slightly shortened (Figure 4.1-2) or hardly recognizable (Figure 4.3-5). Such exceptions are common among very large corallites of the topotype colonies in which the cardinal septum is truly, although indistinctly shortened, as documented by lack of plates attached to intercepts of tabulae within the cardinal fossula (Figure 6.4-5) or by inconsistent development of such plates, i.e., attached to only some sections of tabulae (Figure 6.1, 6.6). Two sections of the same corallite (Figure 6.5-6), one with the intercepts absent and the other with them present, document the inconsistent length of the cardinal septum. Thus, the length of the cardinal septum should be characterized as most commonly long in small and medium-sized corallites, but variable in the largest specimens.
Length of the cardinal septum. Smith (1935) drew special attention to the morphology seen in longitudinal section, but did not mention the cardinal septum, whereas Wilson (1980) characterized it as "somewhat shorter than other major septa." Fedorowski et al. (2007, p. 100) were of a different opinion, pointing to "short plates attached to intercepts of tabulae between the cardinal septum and the median lamella." Those short plates are fragments of the cardinal septum cut between individual tabulae within the shallow cardinal fossula. Such an image resulted from the low elevation of the cardinal septum above the tabulae as it extends to the median lamella along the cardinal fossula floor. We now confirm that the cardinal septum is permanently elongate in all type corallites preserved adequately to demonstrate that character (Figure 2.1-4, 2.7) and in most young and medium-sized corallites within individual topotype colonies. However, several exceptions occur among immature corallites in those colonies. Their cardinal septa may either be slightly shortened (Figure 4.1-2) or hardly recognizable (Figure 4.3-5). Such exceptions are common among very large corallites of the topotype colonies in which the cardinal septum is truly, although indistinctly shortened, as documented by lack of plates attached to intercepts of tabulae within the cardinal fossula (Figure 6.4-5) or by inconsistent development of such plates, i.e., attached to only some sections of tabulae (Figure 6.1, 6.6). Two sections of the same corallite (Figure 6.5-6), one with the intercepts absent and the other with them present, document the inconsistent length of the cardinal septum. Thus, the length of the cardinal septum should be characterized as most commonly long in small and medium-sized corallites, but variable in the largest specimens.
Derivation of the median lamella. The derivation of the median lamella in the type specimens and topotypes is discussed in detail in the blastogeny section below. Its elongation towards the cardinal septum (Figure 2.1-4, 2.7, Figure 3.1-5, Figure 4.1-5), observed in both juvenile and mature corallites and irrespective of the length of the cardinal septum, is typical for the family Aulophyllidae. It is crucial for the distinction of Heritschioides s.s. from Permian species included in that genus by various authors (e.g., Wilson and Langenheim, 1962; Minato and Kato, 1965; Stevens, 1967; Rowett, 1969; Wilson, 1982). The median lamella in those Permian species is either lacking in early blastogeny and underdeveloped at the mature growth stage or is derived from the counter septum. They were transferred by Fedorowski et al. (2007) to various genera within the families Durhaminidae Minato and Kato, 1965 (the former group) and Kleopatrinidae Fedorowski, Bamber and Stevens, 2007 (the latter group) and are omitted from further discussion.
Morphology of the axial structure as seen in transverse section. The morphology of the axial structure has not previously been adequately characterized. The median lamella is simple, weak and variable in shape and length in young corallites and in most of those of medium size. It tends to elongate towards the cardinal septum when well developed, rarely joins it. The median lamella is thin and simple, but easily distinguishable in some of the largest mature corallites (Figure 6.1-2), whereas it is totally reduced or barely distinguishable from the septal lamellae in others (Figure 3.1-5, Figure 6.3-4, 6.8). Over a few millimeters of corallite growth, the median lamella may change from being solid to broken (Figure 6.5-6, respectively). It may also be difficult to recognize because of diagenetic alteration (Figure 2.3, 2.7, Figure 6.3). The septal lamellae vary greatly in number and shape (Figure 6.1-4, 6.8). They are seldom continuous, expanding from the periphery of the axial structure to its axis or to the median lamella. More commonly, thin segments of septal lamellae attached to axial tabellae are observed. Some of them may occur as short plates attached to the sclerenchyme surrounding the axial structure (Figure 6.7). A combination of comparatively long septal lamellae and short plates may also occur. It must be pointed out that the general morphology of the axial structure in transverse section closely resembles that of genera in the subfamily Aulophyllinae.
Neither Smith (1935) nor Wilson (1980) mentioned septal lamellae in the axial structure corresponding to minor septa. Those lamellae, called extra septal lamellae by Fedorowski et al. (2007, p. 43), are another important character pointing towards the Aulophyllidae. Rare extra septal lamellae appear in the holotype fairly early in its hystero-ontogeny (Figure 2.1, right), but only as barely recognizable plates at the periphery of the axial structure in some septal loculi. They are absent from the transverse section of the complete paratype GSC 9060. However, observations on a single transverse section cannot be considered conclusive when dealing with such a variable characteristic. Our study of the topotypes has demonstrated great intracolony variability of the extra septal lamellae within the axial structures of the largest corallites (Figure 6.1-2, 6.5-8). They are either absent or occur in advanced maturity as very short plates attached to axial tabellae or exclusively to the sclerenchymal thickening at the periphery of the axial structure. Long extra septal lamellae are exceptional. Thus, they commonly follow the morphology and shape of the regular septal lamellae. They are definitely absent from the immature corallites of the topotype colonies, and their occurrence cannot be confirmed in most fully mature corallites sectioned. Therefore, their appearance may be temporary and generally short-lived during the growth of individual corallites. As a result they are rarely seen in randomly cut transverse sections. Thus the occurrence of extra septal lamellae should not be taken as an obligatory diagnostic character for the genus, but only as an auxiliary feature which is present, but is not always intersected in transverse section. Extra septal lamellae occur in several genera of the Aulophyllidae (e.g., Aulophyllum Milne Edwards and Haime, 1850; Berkhia Gorsky, 1951; Nervophyllum Vassilyuk, 1959), but are absent from other taxa of that family. Nevertheless their occurrence in the holotype and other colonies of Heritschioides supports the idea of a relationship between the Heritschioidinae and the Aulophyllinae.
Morphology of the axial structure as seen in longitudinal section. This feature was adequately described by Smith (1935) and Wilson (1980). Thus, only the continuous nature of that structure, i.e., the presence of a continuous axial column (Fedorowski et al., 2007) needs mentioning here as diagnostic for the genus. Continuous axial columns occur in longitudinally sectioned corallites from both the type collection (Figure 2.5-6) and the topotypes (Figure 3.6-8). An occasional extension of tabellae from the tabularium to the peripheral part of the axial column, pointed out by Smith (1935), means only that the boundary of the latter may not be sharp throughout its length. The corallite width in longitudinal section may be larger than the maximum diameter listed in the diagnosis because some corallites commonly are oval rather than circular. The largest diameter (21 mm) corresponds to a strongly oval corallite sectioned along its long axis. Proportions of particular structural elements: the dissepimentarium, the tabularium and the axial column remain similar regardless of the general diameter.
Biform morphology in the tabularium. This feature was first mentioned for Heritschioides by Fedorowski et al. (2007). We confirm their information, but add the following: 1. Biform morphology is absent from some septal loculi of the holotype corallites as shown by the symmetrical arrangement of tabula intercepts with respect to some minor septa. However, it is present in the adjacent loculi, as shown by the different arrangement of those intercepts (Figure 5.8). 2. The abundance of incomplete tabulae masks the biform morphology to the extent that it is difficult to recognize (Figure 2.5, upper left). 3. Clinotabellae mentioned by Wilson (1980) do not occur in Heritschioides. Tabellae in position I of Sutherland (1965) are perhaps what he had in mind. The same is true for the U-shaped tabellae mentioned by Smith (1935).
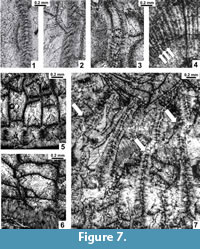 Microstructure of septa and diagenesis. The septal microstructure has been diagenetically altered in all specimens studied. Three types of artificial images can be distinguished: 1) A solid, narrow, straight or zig-zagged "middle dark line," white in pictures (Figure 7.3, upper, Figure 7.7), i.e., the primary septum flanked by light crystalline fibrils, may represent the most transformed structure. Suspected trabeculae, approximately corresponding in size to the width of the "dark line," may have been destroyed and combined. The middle parts of the "dark lines" may be replaced by white crystalline calcite during further diagenesis. In some septa this transformation is observed (Figure 7.3, lower and upper). 2) Symmetrically arranged bunches of crystalline fibrils (Figure 7.1) represent remnants of trabeculae, slightly enlarged by recrystallization that followed the main directions of crystal growth. This is the most common feature observed. The fans of trabeculae visible in longitudinal sections (Figure 7.4, arrows) confirm the trabecular microstructure of septa, although primary diameters of trabeculae may be slightly smaller. 3) Bodies arranged obliquely to the length of a septum (Figure 7.2) also represent remnants of trabeculae deformed by both further diagenetic alteration and biased by slight obliqueness of the section. All of the above features may be present within the same transverse section of a given corallite.
Microstructure of septa and diagenesis. The septal microstructure has been diagenetically altered in all specimens studied. Three types of artificial images can be distinguished: 1) A solid, narrow, straight or zig-zagged "middle dark line," white in pictures (Figure 7.3, upper, Figure 7.7), i.e., the primary septum flanked by light crystalline fibrils, may represent the most transformed structure. Suspected trabeculae, approximately corresponding in size to the width of the "dark line," may have been destroyed and combined. The middle parts of the "dark lines" may be replaced by white crystalline calcite during further diagenesis. In some septa this transformation is observed (Figure 7.3, lower and upper). 2) Symmetrically arranged bunches of crystalline fibrils (Figure 7.1) represent remnants of trabeculae, slightly enlarged by recrystallization that followed the main directions of crystal growth. This is the most common feature observed. The fans of trabeculae visible in longitudinal sections (Figure 7.4, arrows) confirm the trabecular microstructure of septa, although primary diameters of trabeculae may be slightly smaller. 3) Bodies arranged obliquely to the length of a septum (Figure 7.2) also represent remnants of trabeculae deformed by both further diagenetic alteration and biased by slight obliqueness of the section. All of the above features may be present within the same transverse section of a given corallite.
The external corallite walls were originally septothecal, formed by peripheral margins of strongly and equally thickened major and minor septa, which thin rapidly inward. The peripheral septal margins are embedded in the thin outermost fibrous layer of the external wall (Figure 7.5). The original microstructure is only rarely preserved in the specimens studied, being most commonly replaced by the irregularly fibrous material of the external wall to which thin, dissepimental sectors of septa are attached. Only slight protrusions of the external wall at the septal attachments (Figure 7.6) can be interpreted as peripheral margins of septa completely altered diagenetically.
 Diagenetic alteration has affected the macrostructures of corallites and colonies as well. Replacement of calcite by dolomite or silica has occurred in all colonies investigated, including the type specimens (Figure 2.2-3, 2.6-7, Figure 3.1, 3.3, 3.5, Figure 4.1, 4.3-5, Figure 8.1-3, whitened areas). Its distribution is irregular and appears random. Deformation by compaction, resulting in fragmentation of some skeletal elements, is common as well (Figure 2.6-7, Figure 6.3). It may be accompanied by a displacement of parts of colonies (Figure 3.5) or parts of individual corallites (Figure 4.5, left, Figure 7.7, arrows). Also, corallites within colonies are not necessarily in their original positions. This is difficult to recognize unless interference and destruction of adjacent external corallite walls is evident (Figure 4.1-2). Therefore, the distances between individual corallites within colonies should be used as a diagnostic feature for species only when lack of compression within a given colony can be demonstrated. Corrosion of the external parts of corallites, advanced in some instances (Figure 4.4), should also be mentioned.
Diagenetic alteration has affected the macrostructures of corallites and colonies as well. Replacement of calcite by dolomite or silica has occurred in all colonies investigated, including the type specimens (Figure 2.2-3, 2.6-7, Figure 3.1, 3.3, 3.5, Figure 4.1, 4.3-5, Figure 8.1-3, whitened areas). Its distribution is irregular and appears random. Deformation by compaction, resulting in fragmentation of some skeletal elements, is common as well (Figure 2.6-7, Figure 6.3). It may be accompanied by a displacement of parts of colonies (Figure 3.5) or parts of individual corallites (Figure 4.5, left, Figure 7.7, arrows). Also, corallites within colonies are not necessarily in their original positions. This is difficult to recognize unless interference and destruction of adjacent external corallite walls is evident (Figure 4.1-2). Therefore, the distances between individual corallites within colonies should be used as a diagnostic feature for species only when lack of compression within a given colony can be demonstrated. Corrosion of the external parts of corallites, advanced in some instances (Figure 4.4), should also be mentioned.
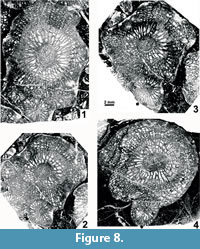
Offsetting. The development of offsets has not been described by earlier authors dealing with H. columbicum and is incompletely described here. Offsetting corallites are absent from the type material, whereas recrystallization and other diagenetic alterations of the topotype colonies prevent their complete study by means of serial sections with peels and photographs. A short series of thin sections (Figure 8.1-3) and two short series of drawings (Figure 9) demonstrate inconsistency during offsetting. The following important, generally applicable characters can be pointed out:
1. 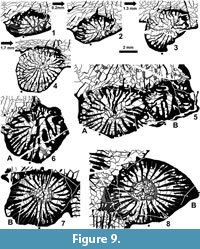 There is a long-lasting connection between the offsets and the parent polyp. A partition proving full connection of the parent and the offset body remains active up to the growth stage having a well developed axial structure (Figure 8.2-4, Figure 9.8).
There is a long-lasting connection between the offsets and the parent polyp. A partition proving full connection of the parent and the offset body remains active up to the growth stage having a well developed axial structure (Figure 8.2-4, Figure 9.8).
2. The counter septum is inherited from the parent polyp, and the cardinal septum is either inserted as a neoseptum or it is inherited (?). Both of those protosepta are long, approaching and temporarily reaching the offset's axis, but their axial union is rarely obvious (e.g., Figure 9.1-2, 9.7) and is inconsistent in all offsets studied.
3. All offsets observed show an unstable connection between the protosepta, considerable elongation of several major metasepta to approach or reach the axial area, and irregular insertion of neosepta. In this combination of characters, the early growth stages of Heritschioides resemble those in the family Durhaminidae.
4. No permanent axial septum is formed (see point 2), and domination of either the cardinal or counter protoseptum either does not occur (Figure 9.3-4) or may differ in adjacent offsets from the same corallite (Figure 9.5A vs 9.6B). Such inconsistency results in the development of a weak and inconsistent median lamella, appearing late in the blastogeny. The inconsistency of the median lamella and the variability of the axial structure as a whole continue up to maturity. The median lamella is never strong, but it may either be easily recognizable in the form of a straight, lath-like body intersecting the axial structure or it may be thin and wavy, barely distinguishable from septal lamellae. Intermediate stages between those two extremes always occur. Transverse sections of some corallites lack a recognizable median lamella. Combinations of all those variants appear in various proportions in all colonies studied.
5. Separation of septal lamellae from the inner margins of the major septa, leading to the development of an axial structure, is a slow and irregular process. Thin, inner margins of major septa with only rare, separated septal lamellae form the earliest axial structure in some offsets (Figure 9.4), whereas in other corallites, the thick inner margins of septa may be separated from their corresponding thin lamellae by strong sclerenchymal secretions (Figure 8.2-3, Figure 9.7-8). Such variation may be the result of differences in the degree of sclerenchymal secretion in offsets of the two colonies observed. The morphology of mature corallites in both colonies is similar irrespective of differences in the morphology of the offsets and the degree of sclerenchymal thickening. Thus, we regard those morphological differences as intraspecific variation.
6. In spite of differences in the formation of the axial structure and in the development of the median lamella, both tend to be connected to the cardinal septum. This is clearly demonstrated by an immature corallite of the holotype colony (Smith, 1935; Wilson, 1980; Fedorowski et al., 2007; Figure 2.1) and is recognizable in the comparable growth stage of several immature corallites of the topotype colonies. Elongation of the axial structure toward the cardinal septum in mature corallites exemplifies that connection best when the axial lamella is recognizable (see above), but may also be demonstrated by the shape of the axial structure in transverse sections whether a weak median lamella is present or not (Figure 6.4).
7. As noted above, extra septal lamellae in the axial structure are rare, short and not always recognizable. Also, the growth level at which they occur is variable and has not been established firmly.
Summing up, the blastogeny described above differs significantly from that in other similar families(i.e., Lithostrotionidae d'Orbigny, 1852; Aulophyllidae Dybowski, 1873; Durhaminidae Minato and Kato, 1965; and Kleopatrinidae Fedorowski, Bamber and Stevens, 2007) so little comparison is required except with the Durhaminidae. There are two main features distinguishing the blastogeny of Heritschioides from that in the Durhaminidae: 1. The leading role of the protosepta in the formation of the axial structure of Heritschioides; temporary domination of either the cardinal or the counter septum early in the blastogeny can be considered unimportant in that context. 2. Extension of the median lamella in Heritschioides and elongation of the axial structure toward the cardinal septum, beginning in the late hystero-neanic growth stage and extending into maturity. At the same time, these differentiating features of Heritschioides suggest that it is related to the family Aulophyllidae. Thus, we consider similarities and differences in the blastogeny as one of the main reasons for both accepting the subfamily Heritschioidinae Sando, 1985 and regarding it as a member of the family Aulophyllidae.
SUMMARY
New data from a detailed study of the type and topotype collections of the type species of Heritschioides confirm that the status of the genus is different than considered previously, and associated microfossils (see above) establish its age as late Serpukhovian/early Bashkirian.
The development and morphology of the axial structure, in particular its connection to the protosepta, are crucial for establishing the taxonomic position of Heritschioides. The close connection of the cardinal septum to the median lamella and the axial structure as a whole, established in mature corallites of the type species of Heritschioides and in five other species of Heritschioides from Alaska and California (Fedorowski et al., in press), points to the family Aulophyllidae. However, in all completely investigated taxa included in that family (e.g., solitary Dibunophyllum and colonial Corwenia) the cardinal septum consistently plays a leading role in the formation of the median lamella and the axial structure, starting from early ontogeny or hystero-ontogeny. In all species of Heritschioides, however, the cardinal septum plays an inconsistent role in that process early in the blastogeny and a connection of the median lamella with the cardinal septum or its elongation towards that septum does not begin until early maturity. A similar development from early hystero-ontogeny to maturity is also present in a new cerioid-aphroid genus from Alaska (authors, personal data). Such a coincidence cannot be treated as either random or as the result of diagenetic alteration, but should be considered an important character for phylogenetic reconstructions. Thus, two different lineages can be distinguished, i.e., taxa with a constant vs an inconsistent role of protosepta in the formation of the median lamella early in ontogeny and/or hystero-ontogeny. Both of these lineages end with a similar relationship of the median lamella and the axial structure to the cardinal septum and the cardinal fossula at maturity. Similarities and differences established in several taxa within each lineage permit inclusion of the subfamily Heritschioidinae within the family Aulophyllidae. Extra septal lamellae present in both Aulophyllum and Heritschioides suggests that the subfamily Heritschioidinae is more closely related to the nominative subfamily than to other subfamilies of the Aulophyllidae.
ACKNOWLEDGMENTS
Specimens from the type locality of Heritschioides were collected by E.W. Bamber, B.C. Richards and M.J. Orchard (Geological Survey of Canada), and W.R. Danner (University of British Columbia). Dating with microfossils was provided by B.L. Mamet and S. Pinard (foraminifers) and M.J. Orchard (conodonts). We express our sincere thanks for the help given by these colleagues. We gratefully acknowledge G. Edwards and D. Then for their help in finalizing the photography and digital processing of figures, and G. Martin for preparing thin sections at GSC Calgary. We especially thank M. Bartkowiak for making additional thin sections and peels and Adam Mickiewicz University in Poznań for providing all facilities necessary for the completion of the first author's part of this paper. Field work and preparation of this paper were supported by the Geological Survey of Canada. Finally we are grateful to the anonymous Referee 2 for his/her constructive critique.
REFERENCES
Armstrong, A.K. 1972. Pennsylvanian carbonates, paleoecology and rugose colonial corals, north flank, eastern Brooks Range, arctic Alaska. USGS Professional Paper, 747:1-21.
Barnes, W.C. and Ross, J.V. 1975. The Blind Creek Limestone, Keremeos, British Columbia: structure and regional tectonic significance. Canadian Journal of Earth Sciences, 12:1929-1933.
Bell, W.A. 1929. Horton-Windsor-District, Nova Scotia. Geological Survey of Canada Memoir155, 1-268.
Cocke, J.M. 1969. Taxonomic and environmental study of two dibunophyllid coral species from Upper Pennsylvanian rocks of Kansas. Kansas University Paleontological Contributions, Paper 44:1-11.
Cocke, J.M. 1970. Dissepimental rugose corals of Upper Pennsylvanian (Missourian) rocks of Kansas. Kansas University Paleontological Contributions, Article 54 (Coelenterata 4):1-67.
Danner, W.R. 1997. Fusulinids and other Paleozoic Foraminifera of accreted terranes, southwestern British Columbia and northwestern Washington, p. 21-25. In Ross, C.A., Ross, J.R.P., and Brenckle, P.L. (eds.), Late Paleozoic Foraminifera, their biostratigraphy, evolution and paleoecology and the mid-Carboniferous boundary. Cushman Foundation for Foraminiferal Research, Special Publication 36, Harvard University, Cambridge, Massachusetts, USA.
Danner, W.R., Nestell, M.K., and Nestell, G.P. 1999. Geology and paleontology of the Carboniferous and Permian of the exotic terranes of southwestern British Columbia. Precongress field trip 9, XIV International Congress on the Carboniferous and Permian.
de Groot, G.E. 1963. Rugose corals from the Carboniferous of northern Palencia (Spain). Leidse Geologische Mededelingen, 29:1-124.
Dobrolyubova, T.A. 1936. Koralli Rugosa Srednego i Verkhnego Karbona i Nizhnei Permi severnogo Urala (123-i list). Trudy Polyarnoi Komissii, 28:77-158. (In Russian)
d'Orbigny, A. 1852. Cours Elémentaire de Paleontologie et Géologie Stratigraphique (Volume 2, part 1). Victor Masson, Paris (after Hill, 1981).
Dybowski, W.N. 1873. Monographie der Zoantharia Sclerodermata Rugosa aus der Silurformation Estlands, Nordlivlands und der Insel Gotland. Archiv für Naturkunde der Livlands, Estlands, Kurlands, 1:257-414.
Fedorowski, J. 1967. The Lower Permian Tetracoralla and Tabulata from Treskelodden, Vestspitsbergen. Norsk Polarinstitutt Skrifter, 142:11-44.
Fedorowski, J. 1981. Carboniferous corals: distribution and sequence. Acta Palaeontologica Polonica, 26:87-160.
Fedorowski, J. 2004. Considerations on most Rugosa and the Dividocorallia from de Groot (1963) collection. Scripta Geologica, 127:71-311.
Fedorowski, J. and Bamber, E.W. 2012. Paleobiogeographic significance of Bashkirian (Pennsylvanian) rugose corals from northernmost Ellesmere Island, Arctic Canada. Geologica Belgica, 15:350-354.
Fedorowski, J., Bamber, E.W., and Baranova, D.V. 2012. An unusual occurrence of Bashkirian (Pennsylvanian) rugose corals from the Sverdrup Basin, arctic Canada. Journal of Paleontology, 86:979-995.
Fedorowski, J., Bamber, E.W., and Stevens, C.H. 2007. Lower Permian Rugose Corals, Western and Northwestern Pangea: Taxonomy and Distribution. National Research Council Research Press, Ottawa, Ontario, Canada.
Fedorowski, J., Stevens, C.H., and Katvala, E. in press. New Late Carboniferous Heritschioidinae (Rugosa) from the Kuiu Island area and Brooks Range, Alaska. Geologica Acta.
Gorsky, I.I. 1951. Kamennougolnye i permskie korally Novoy Zemli. Trudy Nauchno-issledovatelskogo Instituta Geologii Arktiki, 32:1-168. (In Russian with English summary)
Hayasaka, I. 1924. On the fauna of the anthracolitic limestone of Omi-mura in the western part of Echigo. Tohoku Imperial University, Scientific Reports, Series 2 (geology), 8:1-83.
Heritsch, F. 1944. Permische Korallen aus dem Bükk-Gebirge in Ungarn. Annales Historico-Naturales Musei Nationalis Hungarici, 37:48-65.
Hill, D. 1981. Supplement 1. Rugosa and Tabulata p. i-xl+F1-F762. In Teichert, C. (ed.), Treatise on invertebrate paleontology. Part F. Coelenterata. Geological Society of America and University of Kansas Press. Boulder, Colorado and Lawrence, Kansas.
Hudson, R.G.S. 1958. Permian corals from northern Iraq. Palaeontology, 1:174-192.
Kawamura, T. and Stevens, C.H. 2012. Middle Pennsylvanian rugose corals from the Baird Formation, Klamath Mountains, northwestern California. Journal of Paleontology, 86:513-520.
Kossovaya, O.L. 1997. Rugozi tipovikh razrezov Gzhel'skogo-Artinskogo Yarusov Severnogo Timana i zapadnogo sklona Urala. Glava 3, p. 53-96, 106-115, 128-154. In Stukalina, G.A. (ed.), Atlas etalonnikh kompleksov Paleozoiskoi bentosnoi fauni Severo-vostoka Evropeiskoi Rossii. Vserossiiskiy Nauchno-issledovatelskiy Geologicheskiy Institut im A. P. Karpinskogo (VSEGEI), St. Petersburg. (In Russian)
Lewis, H.P. 1935. The Lower Carboniferous corals of Nova Scotia. Annals and Magazine of Natural History. Series 10, 16:118-142.
Lin, B.Y., Xu, S.Y., Jia, H.Z., Guo, S.Z., Ouyang, X., Wang Z.J., Ding Y.J., Cao X.D., Yan,Y., and Chen, H.C. 1995. Monograph of Palaeozoic Corals. Rugosa and Heterocorallia. Geological Publishing House, Beijing. (In Chinese with English summary)
McCoy, F. 1849. On some new genera and species of Palaeozoic corals and Foraminifera. Annals and Magazine of Natural History, 2:1-20, 119-136.
Milne-Edwards, H. and Haime, J. 1850. A Monograph of the British Fossil Corals. Palaeontographical Society, London.
Minato, M. and Kato, M. 1965. Durhaminidae (Tetracoral). Journal of the Faculty of Science, Hokkaido University. Series 4, Geology and Mineralogy, 13:13-86.
Moore, R.C. and Jeffords, R.M. 1941. New Permian corals from Kansas, Oklahoma, and Texas. State Geologcial Survey of Kansas Bulletin, 38, 1941 Reports of Studies, 3:65-120.
Moore, R.C. and Jeffords, R. 1945. Description of Lower Pennsylvanian corals from Texas and adjacent states. University of Texas, Publication 4401:1-208.
Newell, N.D. 1935. Some Mid-Pennsylvanian invertebrates from Kansas and Oklahoma. H, Stromatoporoidea, Anthozoa and Gastropoda. Journal of Paleontology, 9:341-355.
Rodrìguez, S. 1984. Corales rugosos del Carbonifero del este de Asturias. Published PhD Thesis, Departamento de Paleontologia, Facultad de Ciencias Geológicas, Universidad Complutense de Madrid, España.
Rowett, C.L. 1969. Upper Paleozoic stratigraphy and corals from the east-central Alaska Range, Alaska. Arctic Institute of North America, Technical paper, 23:7-120.
Rowett, C.L. and Sutherland, P.K. 1964. Biostratigraphy and rugose corals of the Lower Pennsylvanian Wapanucka Formation of Oklahoma. Geological Survey of Oklahoma Bulletin, 104:1-124.
Sando, W.J. 1985. Paraheritschioides, a new rugose coral genus from the Upper Pennsylvanian of Idaho. Journal of Paleontology, 59:979-985.
Sando, W.J., Bamber, E.W., and Armstrong, A.K. 1975. Endemism and similarity indices: Clues to the zoogeography of North American corals. Geology, 3:661-664.
Smith, S. 1935. Two Anthracolithic corals from British Columbia and related species from the Tethys. Journal of Paleontology, 9:30-42.
Smith, S. and Ryder, T.A. 1926. The genus Corwenia, gen. nov. Annals and Magazine of Natural History, Series 9, 17:149-159.
Stevens, C.H. 1967. Leonardian (Permian) compound corals of Nevada. Journal of Paleontology, 41:423-431.
Stevens, C.H. and Rycerski, B. 1989. Early Permian colonial rugose corals from the Stikine River Area, British Columbia, Canada. Journal of Paleontology, 63:158-181.
Stuckenberg, A.A. 1904. Korally i mshanki nizhnego otdela srednerusskogo, kamennougol'nogo izvestnyaka. Trudy Geologischeskogo Komiteta, 14:1-109. (In Russian and German)
Sutherland, P.K. 1965. Henryhouse rugose corals. Oklahoma Geological Survey Bulletin, 109:1-92.
Thomson, J. 1874. Description of new corals from the Carboniferous Limestone of Scotland. Geological Magazine, 2:556-559.
Thomson, J and Nicholson, H.A. 1876. Contribution to the study of the chief generic types of the Palaeozoic corals. Annals and Magazine of Natural History, 17:60-70, 123-128, 290-305, 451-461.
Vassilyuk, N.P. 1959. Novie tetracoralli iz Nizhnego Karbona Donetskogo Basseina. Paleontologicheskiy Zhurnal, 4:85-89. (In Russian)
Vassilyuk, N.P. and Kosyreva, T.A. 1974. Noviy rod korallov Copia (Rugosa) iz Nizhnego Karbona Voronezhskoi Anteklizi. Paleontologicheskiy Sbornik, 11:31-34. (In Russian)
Verrill, A.E. 1865. Classification of polyps (extract condensed from a synopsis of the polypi of the North Pacific Exploring Expedition under captains Ringgold and Rodgers, U.S.N). The Essex Institute Proceedings, 4:145-149.
Wang, H.C. 1950. A revision of the Zoantharia Rugosa in the light of their minute skeletal structures. Philosophical transactions of the Royal Society of London. Series B, Biological Sciences, 611:175-264.
Wilson, E.C. 1980. Redescription of type specimens of the Permian rugose coral Waagenophyllum columbicum Smith, 1935, type species of Heritschioides Yabe, 1950. Journal of Paleontology, 54:85-92.
Wilson, E.C. 1982. Wolfcampian rugose and tabulate corals (Coelenterata; Anthozoa) from the Lower Permian McCloud Limestone of northern California. Contributions in Science of Natural History Museum of Los Angeles County, 337:1-90.
Wilson, E.C. 1994. Early Permian corals from the Providence Mountains, San Bernardino County, California. Journal of Paleontology, 68:938-951.
Wilson, E.C. and Langenheim, L.R., Jr. 1962. Rugose and tabulate corals from Permian rocks in the Ely Quadrangle, White Pine County, Nevada. Journal of Paleontology, 36:495-520.
Yabe, H. 1950. Permian corals resembling Waagenophyllum and Corwenia. Proceedings of the Japanese Academy, 26:74-79.
