

 Angiosperm pollen grains from the Cuayuca Formation (Late Eocene to Early Oligocene), Puebla, Mexico
Angiosperm pollen grains from the Cuayuca Formation (Late Eocene to Early Oligocene), Puebla, Mexico
Article number: 18.1.2A
https://doi.org/10.26879/465
Copyright Paleontological Society, January 2015
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 27 February 2014. Acceptance: 19 December 2014
{flike id=1027}
ABSTRACT
Systematic descriptions and illustrations of the best preserved angiosperm pollen grains (Monocotyledonae or Liliopsida: n= 7 and Dicotyledonae or Magnoliopsida: n= 41) recovered from Cuayuca Formation (late Eocene-early Oligocene), Puebla State, Mexico are provided, some of them of chronostratigraphic importance ( Aglaoreidia pristina, Armeria, Bombacacidites, Corsinipollenites, Eucommia, Favitricolporites, Intratriporopollenites, Lymingtonia, Magnaperiporites, Malvacipollis spinulosa, Margocolporites aff. vanwijhei, Momipites coryloides, Momipites tenuipolus, Mutisiapollis, Ranunculacidites operculatus, and Thomsonipollis sabinetownensis ). Taxa identified from the Cuayuca Formation suggest local semiarid vegetation such as tropical deciduous forest, chaparral, grassland, and arid tropical scrub, in which angiosperms are one of the main representatives. Nevertheless, temperate taxa from Pinus forest and cloud forest were also registered from regional vegetation. It is noticeable that at the present time, such taxa are well represented in the vegetation of the Balsas River Basin, which would suggest the existence of this type of flora in the Cuayuca region since the Oligocene.
Elia Ramírez-Arriaga. Departamento de Paleontología, Instituto de Geología, Universidad Nacional Autónoma de México, Av. Universidad 3000, Ciudad Universitaria, C.P. 04510, México, Distrito Federal, elia@unam.mx.
Mercedes B. Prámparo. Centro Regional de Investigaciones Científicas y Tecnológicas, Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales, Unidad de Paleopalinología, Mendoza, Argentina. mprampar@mendoza-conicet.gob.ar
Enrique Martínez-Hernández. Departamento de Paleontología, Instituto de Geología, Universidad Nacional Autónoma de México, Av. Universidad 3000, Ciudad Universitaria, C.P. 04510, México, Distrito Federal. emar@unam.mx
Keywords: systematic palynology; angiosperm pollen grains; Cuayuca Formation; Paleogene vegetation; Mexican Cenozoic basins
Final citation: Ramírez-Arriaga, Elia, Prámparo, Mercedes B., and Martínez-Hernández, Enrique. 2014. Angiosperm pollen grains from the Cuayuca Formation (Late Eocene to Early Oligocene), Puebla, Mexico. Palaeontologia Electronica 18.1.2A: 1-38. https://doi.org/10.26879/465
palaeo-electronica.org/content/2015/1027-angiosperm-pollen-cuayuca-fm
INTRODUCTION
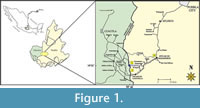 The Cuayuca Formation was described by Fries (1966), who designated a Miocene-Pliocene age. Subsequently, palynostratigraphic research has allowed it to be re-assigned to a late Eocene-early Oligocene age based on the presence of some index taxa, such as Cicatricosisporitesdorogensis, Aglaoreidiapristina, Ephedripitesclaricristatus, Malvacipollisspinulosa, Margocolporites aff. vanwijhei, Momipitestenuipolus, Ranunculaciditesoperculatus, Polyadopollenites aff. pflugii, and Thomsonipollis sabinetownensis (Ramírez-Arriaga et al., 2006, 2008). The Cuayuca Formation is within the Balsas Group; it was deposited in central Mexican Tertiary basins generated through tectonic events (Silva-Romo et al., 2000). Currently, those Paleogene basins are distributed throughout the Mezcala-Balsas hydrological system (Fries, 1960) also known as the Balsas River Basin. The Cuayuca Formation yielded a diverse and well preserved terrestrial palynological assemblage obtained from nine outcrop sections in Puebla State, southern Mexico (Figure 1). The palynoflora studied includes gymnosperm and angiosperm pollen grains, as well as pteridophyte and bryophyte spores. Ramírez-Arriaga et al. (2005) published systematic descriptions of spores and gymnosperm pollen grains.
The Cuayuca Formation was described by Fries (1966), who designated a Miocene-Pliocene age. Subsequently, palynostratigraphic research has allowed it to be re-assigned to a late Eocene-early Oligocene age based on the presence of some index taxa, such as Cicatricosisporitesdorogensis, Aglaoreidiapristina, Ephedripitesclaricristatus, Malvacipollisspinulosa, Margocolporites aff. vanwijhei, Momipitestenuipolus, Ranunculaciditesoperculatus, Polyadopollenites aff. pflugii, and Thomsonipollis sabinetownensis (Ramírez-Arriaga et al., 2006, 2008). The Cuayuca Formation is within the Balsas Group; it was deposited in central Mexican Tertiary basins generated through tectonic events (Silva-Romo et al., 2000). Currently, those Paleogene basins are distributed throughout the Mezcala-Balsas hydrological system (Fries, 1960) also known as the Balsas River Basin. The Cuayuca Formation yielded a diverse and well preserved terrestrial palynological assemblage obtained from nine outcrop sections in Puebla State, southern Mexico (Figure 1). The palynoflora studied includes gymnosperm and angiosperm pollen grains, as well as pteridophyte and bryophyte spores. Ramírez-Arriaga et al. (2005) published systematic descriptions of spores and gymnosperm pollen grains.
There are several publications dealing with Mexican Cenozoic palynology (Graham, 1976; Martínez-Hernández et al., 1980; Tomasini-Ortíz and Martínez-Hernández, 1984; Palacios and Rzedowsky, 1993; Martínez-Hernández and Ramírez-Arriaga, 1996, 1999; Graham, 1999), but taxonomic studies with detailed descriptions of Paleogene and Neogene pollen grains are lacking in most of those studies.
The present contribution deals with the taxonomy and systematics of selected angiosperm pollen grains (Monocotyledonae or Liliopsida and Dicotyledonae or Magnoliopsida) obtained from the Cuayuca Formation, some of them of chronostratigraphic importance. The Monocotyledonae (Liliopsida) pollen grains are represented by monosulcate and monoporate aperture types and Dicotyledonae (Magnoliopsida) pollen grains include colpate, colporoidate, colporate, porate, and inaperturate pollen types. This contribution constitutes one of the first taxonomic descriptions of angiosperm pollen grains recovered from the evaporites of the Cuayuca Formation, which provides useful information for a better understanding of the paleobiodiversity and distribution of the Mexican plant communities during the late Paleogene.
MATERIAL AND METHODS
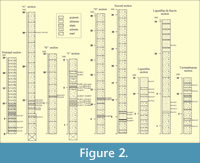 A total of 49 samples were collected from nine outcrop sections, with 21 samples bearing palynomorphs (Figure 2). All samples were processed through standard methods (HCI and HF). The slides obtained have been stored in the UNAM (Universidad Nacional Autónoma de México), at the Geology Institute Palynological Collection (IGLUNAM). Samples were examined with a Zeiss transmitted light microscope, and the photomicrographs were taken with a Sony digital camera. In order to highlight the morphology, some photographs were taken in phase-contrast. Coordinates of the illustrated specimens were provided, most of them using an England Finder (EF) slide. Total counts of taxa at the different sections studied are shown in Table.
A total of 49 samples were collected from nine outcrop sections, with 21 samples bearing palynomorphs (Figure 2). All samples were processed through standard methods (HCI and HF). The slides obtained have been stored in the UNAM (Universidad Nacional Autónoma de México), at the Geology Institute Palynological Collection (IGLUNAM). Samples were examined with a Zeiss transmitted light microscope, and the photomicrographs were taken with a Sony digital camera. In order to highlight the morphology, some photographs were taken in phase-contrast. Coordinates of the illustrated specimens were provided, most of them using an England Finder (EF) slide. Total counts of taxa at the different sections studied are shown in Table.
Statistical Analysis
In order to determine vegetation types, taxa were classified by means of multivariate analysis via hierarchical clustering analysis, in which groups were defined using the Ward method with a Euclidian distance of 1.8; this was done using program R (Johnson, 2000). In addition, Constrained Cluster Analysis (CONISS) through the incremental sum of squares method was applied using the Tilia Graph program, in order to define paleopalynological changes in the Cuayuca Formation Mcy Member (Grimm, 1987). This analysis was also able to give information concerning the most representative elements in each paleocommunity.
The Parsimony Analysis of Endemicity (PAE)
Parsimony Analysis of Endemicity (Rosen, 1988; Morrone and Crisci, 1995; Ramírez-Arriaga et al., 2008, 2014) was used to compare the palynomorph diversity of Cuayuca Formation outcrop sections: 1) principal section; 2) section “A” (Cuay A); 3) section “B” (Cuay B); 4) section “F” (IzF); 5) section “H” (IzH); 6) second section (IzS); 7) Lagunillas section (Lag); 8) Lagunillas de Rayón section (LagRay); and 9) Tzompahuacan section (Tzo).
The WINCLADA data matrix from PAE analysis (Nixon, 2002) shows rows that correspond to outcrop sections and columns for palynomorphs, whose presence (1) or absence (0) in each section is indicated. In hierarchic grouping, sections were clustered according to the presence of taxa. NONA was used to carry out the parsimony analysis (Goloboff, 1993). The PAE analysis included a heuristic search with tree bisection and reconnection (TBR), branch swapping with 500 replications holding 10 trees per replication, and further expanding the memory to hold up to 10,000 trees. WINCLADA calculated three statistics that reflect the degree of conflict among data: a) Length=L; b) Consistency Index=CI; and c) Retention Index=RI. In the present work, clades are supported by the presence of palynomorphs, and unique occurrences are emphasized with black circles.
SYSTEMATIC PALAEONTOLOGY
A total of 114 taxa of angiosperm pollen grains were identified from the Cuayuca Formation (Ramírez-Arriaga et al., 2006). The Dicotyledonae group was the most representative and diverse, with 95 taxa. The Monocotyledonae showed a lower diversity of only 19 taxa. Because of the low number of specimens of each taxon present in the assemblage, the assignment to known or new species has been problematic. Thus, 48 taxa were classified only to the genus level, with open nomenclature at the species level (Appendix 1). These taxa are fully described, illustrated, presented as Monocotyledonae or Dicotyledonae, and are in alphabetical order: 41 of them are Dicotyledonae and seven Monocotyledonae, with the botanical affinities provided in Appendix 2. In general, scientific names have been cited according to a parataxonomic classification of fossil taxa. These diagnoses are based on Jansonius and Hills (1976) catalogue, as well as on other publications cited in the text. Only cases which show a clear affinity to a recent genus are cited as modern genus pollen types (e.g., Armeria sp.), using the natural classification system.
The classification down to family level and author names of orders and families follows the APG III (2009) and Cronquist (1981). In the case of some fossil genera of controversial botanical affinity, we gave the taxonomic range up to Class or Order rank. For each taxon, the distribution is given for different Mexican basins, and in North America and Europe, with the exception of some comparisons to Southern Hemisphere specimens. Descriptive terminology is based on El Ghazali (1990), Traverse (1988), and Punt et al. (2007).
Kingdom PLANTAE Haeckel, 1866
Division MAGNOLIOPHYTA Cronquist et al., 1966
Class LILIOPSIDA Cronquist et al., 1966
Genus AGLAOREIDIA Erdtman, 1960 emend. Fowler, 1971
Type species. Aglaoreidiacyclops Erdtman, 1960
Aglaoreidia pristina Fowler, 1971
Figure 3.7
 Material. Sample Pb-9334(4), Palynology Laboratory, IGLUNAM.
Material. Sample Pb-9334(4), Palynology Laboratory, IGLUNAM.
Description. Monad pollen, heteropolar, radiosymmetric, amb circular, spheroidal. Monoporate, pore with annulus 1.2 μm thick. Exine semitectate, columellate, 0.8 μm thick, reticulate, heterobrochate, brochi <0.8 μm near the aperture.
Dimensions. Equatorial diameter 24.8 μm; polar diameter 24 μm based on one measured specimen.
Other occurrences. Mexico - late Eocene to early Oligocene Pie de Vaca Formation, Puebla (Martínez-Hernández and Ramírez-Arriaga, 1999). U.S.A. - late Eocene to early Oligocene, Gulf Coast (Frederiksen, 1980a, 1988). Europe - late Eocene to middle Oligocene, northeastern Europe (Krutsch, 1963); late Eocene, England (Fowler, 1971; Collison et al., 1981).
Genus ARECIPITES Wodehouse, 1933 emend. Nichols et al., 1973
Type species. Arecipitespunctatus Wodehouse, 1933 ex Potonié, 1958
Arecipites sp.
Figure 3.1
Material. Samples Pb-9138, Pb-9337, Pb-8896, Pb-8897, Pb-8899, Pb-8902, and Pb-8872, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, heteropolar, bilateral symmetry, amb oval, oblate. Monosulcate, sulcus extending full length of the grain and flared at ends. Exine tectate, columellate, 0.8 µm thick, microfoveolate with the arrangement of the columellae showing a very fine reticulate pattern.
Dimensions. Length 25.6-32 µm, width 13.6-22 µm, two specimens measured.
Comparisons. This species is very similar to Arecipitessubverrucatus (Barreda, 1997b) although differing in the smaller dimensions of our specimens. The Arecipites sp. from the Cuayuca Formation is not faintly scabrate as in A. pseudotranquilus (Nichols et al., 1973), nor as granulate as in A. pertusus (Elsik in Stover et al., 1966). Arecipites sp. described by Jarzen and Klug (2010) has a scabrate, foveolate to finely reticulate surface, a pattern very similar to the Mexican specimens, although differing in the dimensions (Florida specimens: 45-57 µm long axis; 30-38 µm short axis; Mexican specimens: 25-32 µm long axis, 13-22 µm short axis). The Mexican specimens are very similar to the Arecipites sp. (Christopher et al., 1980, plate 3, figures 1-2) from the Paleocene of Warm Springs, Georgia (Christopher et al., 1980).
Other occurrences. This genus has been reported in Mexican paleobasins from Eocene to Oligocene Simojovel, Chiapas (Tomasini-Ortiz and Martínez-Hernández, 1984). Reports of the occurrence of Arecipites -type pollen in the Caribbean region and in Texas (U.S.A.) are discussed in Jarzen and Klug (2010).
Genus LILIACIDITES Couper, 1953
Type species. Liliaciditeskaitangataensis Couper, 1953
Liliacidites sp. 1
Figure 3.2-3
Material. Samples Pb-8887, Pb-9136, Pb-9334, Pb-8897, Pb-9147, Pb-9138, Pb-9340, Pb-9343, Pb-8898, Pb-8872, and Pb-8896, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, heteropolar, bilateral symmetry, spherical outline. Monosulcate, sulcus extending full length of grain. Exine semitectate, collumelate, 0.8 µm thick, reticulate, heterobrochate, reticulum consisting of large brochi of 2-3 µm, with small brochi at the intersections 0.4 µm and muri 0.5-0.8 µm wide .
Dimensions. Length 22.4-25.6 µm, width 13.6-20.8 µm, six specimens measured.
Comparisons. This taxon has a distinctive reticulum conformed by large brochi with small brochi at the intersections of the muri; this kind of reticulum is neither observed in Liliacidites sp.2 nor in Liliacidites sp.3 of the Cuayuca assemblage. Liliacidites sp.1 has a reticulum very similar to Liliacidites sp. F, illustrated in Plate 3 (Doyle and Robbins, 1975, figures 23-24) from the Raritan Formation (Cretaceous), Delaware, U.S.A. (Doyle and Robbins, 1975).
Other occurrences. Liliaceous-type pollen has been widely distributed throughout the world since the Cretaceous. In Mexico, records of the Liliacidites genus are known from the late Eocene to early Oligocene Pie de Vaca Formation, Puebla (Martínez-Hernández and Ramírez-Arriaga, 1999); Oligocene of the San Gregorio Formation (Martínez-Hernández and Ramírez-Arriaga, 2006); Oligocene to Miocene of Chiapas (Biaggi, 1978); as well as the Neogene (Palacios and Rzedowski, 1993).
Liliacidites sp. 2
Figure 3.6
Material. Sample Pb-9334, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, heteropolar, bilateral symmetry, amb oval. Monosulcate. Exine semitectate, collumelate, 1 µm thick, reticulate, heterobrochate, brochi 1.5-2 µm diminishing size toward the aperture and muri of 0.8 µm wide.
Dimensions. Length 32 µm, width 22 µm, one specimen measured.
Comparisons. Liliacidites sp. 2 has muri distinctively wider (0.8 µm) than those of Liliacidites sp.1 and Liliacidites sp. 3 of the Cuayuca assemblage. Liliacidites sp. 2 has a reticulum very similar to Liliacidites sp. 1 (Wingate, 1983, plate 2, figure 8) from Elko, Nevada (U.S.A), although Liliacidites sp. 1 has greater dimensions: 38 µm length and 30 µm width (Wingate, 1983).
Liliacidites sp. 3
Figure 3.8
Material. Sample Pb-8871, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, heteropolar, bilateral symmetry, amb oval. Monosulcate, aperture reaching the ends of the equator. Exine semitectate, collumelate, 1 µm thick, reticulate, heterobrochate, reticulum consisting of large brochi of 1 µm with small brochi < 1 µm, both randomly distributed, muri < 0.5 µm.
Dimensions. Length 29-46 µm, width 17-28 µm, three specimens measured.
Comparisons. Liliacidites sp. 3 has thinner muri than those in Liliacidites sp. 1 and Liliacidites sp. 2 of the same association. This taxon resembles the specimen Liliaciditesquadrangularis illustrated by Roche and Schuler (1976) from Belgium (plate VII, figures 12-14), but L. quadrangularis has a thicker exine. Liliacidites sp. 3 from Cuayuca Formation closely resembles Liliacidites sp. A (Christopher et al., 1980, plate 3, figures 3-4) from Warm Springs, Georgia, U.S.A. (Christopher et al. , 1980).
Genus MONOCOLPOPOLLENITES Thomson and Pflug, 1953
Type species. Monocolpopollenites tranquillus (Potonié 1934) Thomson and Pflug, 1953
Monocolpopollenites aff. texensis Nichols et al., 1973
Figure 4.7
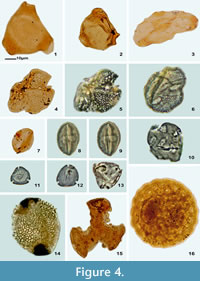 Material. Sample Pb-8872, Palynology Laboratory, IGLUNAM.
Material. Sample Pb-8872, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, heteropolar, bilateral symmetry, amb oval. Monosulcate. Exine tectate, the structure of the wall is difficult to discern, 0.8 µm thick, psilate.
Dimensions. Length 32 µm, width 22.4 µm, one specimen measured.
Remarks. In a monographic discussion concerning monosulcate genera, Nichols et al. (1973) proposed that the shape of the sulcus differentiates the genus Monocolpopollenites from Arecipites. Arecipites includes monosulcate grains with a tapered sulcus, not expanded or gaping at the ends. Monocolpopollenites includes grains with flared or round-ended sulcus.
Comparison. M. aff. texensis (in this contribution) has the sulcus extending the entire long axis of the grain, with margins tending to overlap centrally, as in M. texensis. However, the Mexican specimen is larger (32 µm) than M. texensis (Nichols et al. 1973: 17–19 x 24–28 µm) and presents a psilate instead of scabrate exine.
Other occurrences. There are no previous records of this taxon in Mexico. The genus has been reported from the US Gulf Coast (Frederiksen, 1980a) and in England (Wilkinson and Boulter, 1980).
Genus MONOPOROPOLLENITES Potonié, 1960
Type species. Monoporopollenites graminoides Potonié, 1960
Monoporopollenites sp.
Figure 3.5
Material. Samples Pb-9136, Pb-8870, Pb-9138, Pb-8896, Pb-9147, Pb-8872, and Pb-9334, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, heteropolar, radiosymmetric, amb subcircular. Monoporate, pore diameter 1.5-4 µm, with annulus of 1 to 2 µm thick. Exine tectate, collumelate, 1 µm thick, scabrate.
Dimensions. Diameter 15-39 µm, 19 specimens measured.
Comparison. Monoporopollenites sp. from Cuayuca Formation differs of Monoporopollenitesannulatus (Jaramillo and Dilcher, 2001) because the Mexican specimens are scabrate instead of psilate.
Fossil record. The genus Monoporopollenites has been widely reported in Mexico, the U.S.A., and Central and South America since the late Eocene (Tschudy, 1973; Leopold, 1974; Graham, 1975; Biaggi, 1978; Frederiksen, 1980; Graham, 1988, 1989, 1991; Jaramillo and Dilcher, 2001; Palacios and Rzedowski, 1993; Martínez-Hernández and Ramírez-Arriaga, 1999, 2006).
Class MAGNOLIOPSIDA Cronquist et al., 1966
Genus BROSIPOLLIS Krutzsch, 1968
Type species. Brosipollis salebrosus (Pflug, 1953) Krutzsch, 1968
Brosipollis sp.
Figure 5.14-15
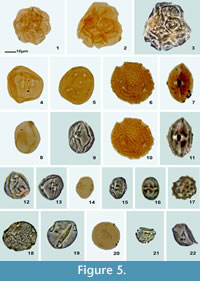 Material. Samples Pb-9340, Pb-8898, Pb-8872, Pb-9138, Pb-9340, and Pb-9334, Palynology Laboratory, IGLUNAM.
Material. Samples Pb-9340, Pb-8898, Pb-8872, Pb-9138, Pb-9340, and Pb-9334, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, subprolate. Tricolporate, endoapertures slightly protruding, lalongate 5 µm x 4 µm with converging transverse costae, meridional edges indistinct. Exine semitectate, columellate, 0.8 µm thick, striate, fine striae-reticulate, about 0.4 µm thick.
Dimensions. Equatorial diameter 13-15 µm, polar diameter 15-27 µm, eight specimens measured.
Comparisons. Brosipollis sp. has similar dimensions, endoaperture, and ornamentation to Rugulitricolporites sp. 1 from the Eocene of California (Frederiksen et al., 1983, plate 23, figures 1-4); but, Brosipollis sp. from the Cuayuca Formation is striate-reticulate instead of simply striate.
Other occurrences. Records of the genus Bursera ( Brosipollis ) are known for the late Eocene - early Oligocene Pie de Vaca Formation of Puebla, Mexico (Martínez-Hernández and Ramírez-Arriaga, 1999); Oligocene San Gregorio Formation (Martínez-Hernández and Ramírez-Arriaga, 2006); Miocene (Palacios and Rzedowski, 1993); and Pliocene (Graham, 1975). U.S.A. - Eocene of California (Frederiksen, 1989; Frederiksen et al. , 1983). This genus was described originally as being from the Malaysian Eocene (Muller, 1968).
Genus FAVITRICOLPORITES Sah, 1967
Type species. Favitricolporites eminens Sah, 1967
Favitricolporites sp.
Figure 3.10
Material. Sample Pb-9138, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, amb circular. Tricolporate syncolpate, colpi with margins 1 µm wide, extending nearly to the poles. Exine semitectate, collumelate, 1 µm thick, reticulate, homobrochate.
Dimensions. Equatorial diameter 16 µm, one specimen measured.
Comparisons. Favitricolporites sp. 1 described by Frederiksen et al. (1983) is similar in ornamentation to the Favitricolporites from the Cuayuca Formation, although Favitricolporites sp. 1 is larger (Frederiksen et al., 1983: 36-50 µm). Unfortunately, no specimens were observed in meridional (equatorial) view, and for this reason the endoaperture could not be described.
Other occurrences. This taxon has been reported from the U.S.A. middle Eocene Mission Valley Formation (Frederiksen et al. , 1983). There are no previous records of the genus in Mexico.
Genus INTRATRIPOROPOLLENITES Thomson and Pflug, 1953 emend . Mai, 1961
Type species. Intratriporopollenites instructus (Potonié, 1931) Thomson and Pflug, 1953
Intratriporopollenites sp.
Figure 3.17
Material. Sample Pb-9340, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, amb circular. Triporate, planaperturate, pores annulate, annulus 1 µm thick. Exine semitectate, columellate, 0.5 µm thick, microreticulate, homobrochate.
Dimensions. Equatorial diameter 20 µm, one specimen measured.
Comparisons. Elsik and Dilcher (1974, plate 29, figures 91-92) reported a similar specimen as Bombacaceae, Sterculiaceae, Tiliaceae? from middle Eocene Claiborne Formation, Tennessee. Other similar Intratriporopollenites (Frederiksen, 1980a, plate 14, figure 17) and Tiliaepollenites (Tschudy and van Loenen, 1970, plate 5, figures 11a, 11b) were reported from upper Eocene, Jackson Group. Tilia was recovered from middle Eocene Kisinger Lake locality (Leopold, 1974: plate 44, figure 18) and Intratriporopollenitesceciliensis from the Eocene of California (Frederiksen, 1989, table 5, figures 2, 3). Intratriporopollenites differs from Bombacacidites because the later taxa is suprareticulate with the greatest diameter of the lumina at the poles, becoming gradually finer toward the apertures and in the intercolpial areas (Nichols, 2010).
Other occurrences. The genus has been reported in Mexico as Oligo-Miocene, Chiapas (Biaggi, 1978); Miocene, Pichucalco, Chiapas (Palacios and Rzedowski, 1993). This taxon has been identified in U.S.A., Tennessee (Elsik and Dilcher, 1974), Kisinger Lakes locality (Leopold, 1974), and California (Frederiksen, 1989).
Genus LYMINGTONIA Erdtman, 1960
Type species. Lymingtonia rhetor Erdtman, 1960
Lymingtonia sp.
Figure 4.10
Material. Samples Pb-9334 and Pb-9340, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, apolar, radiosymmetric, spherical. Pericolpate, 12 costate colpi 7.9-19 µm long and 2.4-4 µm wide, with microverrucate on membrane. Exine semitectate, columellate, 0.8 µm thick, microreticulate.
Dimensions. Diameter 28-31 µm, four specimens measured.
Comparisons. The Lymingtonia sp. described herein is smaller than L. rhetor (Erdtman, 1972: 50 µm), although both have perforations of similar dimensions: about 1.0 µm wide (Jansonius and Hills, 1976: card number 1562). L. cenozoica, described by Pocknall and Mildenhall (1984) from New Zealand, has smaller perforations (< 5 µm), a sparsely conate/spinulose ornamentation and raised crater-like punctae; in contrast, our specimens are microreticulate.
Other occurrences. This genus has been recovered in the U.S.A. - early Eocene of Tennessee (Elsik and Dilcher, 1974); late Eocene of Jackson Group, Mississippi and Alabama (Frederiksen, 1980a). Caribbean islands - middle Miocene of the Saramaguacán Formation, Cuba (Graham, 2000; Graham et al., 2000).
Genus MALVACIPOLLIS Harris, 1965
Type species. Malvacipollis diversus Harris, 1965
Malvacipollis spinulosa Frederiksen, 1983
Figure 5.12
Material. Sample Pb-9334(4), Palynology Laboratory, IGLUNAM.
Description. Monad, isopolar, radiosymmetric, amb circular, spheroidal. Stephanoporate, pores annulate. Exine tectate, supraechinate, echinae solid, 1 - 1.2 µm in height, distributed homogeneously over the surface of pollen grain.
Dimensions. Equatorial diameter 23 (25) 27 µm based on three measured specimens.
Other occurrences. Mexico - late Eocene to early Oligocene Pie de Vaca Formation (Martínez-Hernández and Ramírez-Arriaga, 1999). U.S.A. - late Paleocene Silverado Formation, California (Gaponoff, 1984); Paleocene - Oligocene, South Carolina (Frederiksen, 1980b); late Eocene - early Oligocene of the Elko Formation, Nevada (Wingate, 1983); it was cited as Echiperiporites from the Eocene of Mississippi embayment (Tschudy, 1973); middle Eocene, San Diego, California (Frederiksen et al., 1983). Argentina -middle Miocene, Argentina (Barreda et al., 1998).
Genus RHOIPITES Wodehouse, 1933
Type species. Rhoipites bradleyi Wodehouse, 1933
Rhoipites aff. cryptoporus Srivastava, 1972
Figure 3.21-22
Material. Sample Pb-8890, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, prolate. Tricolporate, endoaperture circular 2 µm in diameter. Exine tectate, columellate, 1 µm thick, micropitted.
Dimensions. Equatorial diameter 15 µm, polar diameter 22 µm, one specimen measured.
Comparisons. This pollen grain resembles a specimen illustrated by Jardine and Harrington (2008) from the Paleocene of Mississippi, U.S.A. It is characterized by its circular endoaperture and its subtectate-reticulate exine; but the specimen from Mississippi is larger (Jardine and Harrington, 2008: 29 µm polar diameter). Differs from R. cryptoporus, which is heterobrochate (Srivastava, 1972).
Other occurrences. Rhoipites has shown worldwide distribution since the late Cretaceous (Srivastava, 1972). R. cryptoporus has been recovered from the late Paleocene of Mississippi, U.S.A. (Jardine and Harrington, 2008) and Alabama (Srivastava, 1972). There are no previous records of this taxon in Mexico.
Rhoipites sp.
Figure 4.8-9
Material. Samples Pb-9334 and Pb-8890, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, subprolate. Tricolporate, endoaperture lalongate 12x3 µm. Exine semitectate, columellate, 1 µm thick, reticulate, homobrochate.
Dimensions. Equatorial diameter 22 µm, polar diameter 27 µm, one specimen measured.
Comparisons. The Rhoipites sp. is distinguished by its homobrochate reticulum instead of the heterobrochate reticulum very evident in Rhoipiteslatus and R. subprolatus from the Eocene of Alabama, U.S.A. (Frederiksen, 1980a). A comparable form has been illustrated by Quattrochio et al. (2000) from the Paleogene of Argentina.
Other occurrences. Rhoipites has a worldwide distribution since Late Cretaceous. There are no previous records of this taxon in Mexico.
Genus STRIATRICOLPORITES Van der Hammen, 1956 ex Leidelmeyer, 1966
Type species. Striatricolporites primulis Leidelmeyer, 1966
Striatricolporites sp.
Figure 5.8-9
Material. Samples Pb-9334, Pb-9340, Pb-8898, Pb-9138, Pb-8896, and Pb-9136, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, subprolate. Tricolporate, endoaperture lalongate with converging transverse costae, meridional edges indistinct. Exine semitectate, 0.8 µm thick, striate, striae 0.5 µm parallel to polar axis.
Dimensions. Equatorial diameter 16-21 µm, polar diameter 28-33 µm, five specimens measured.
Comparisons. Striatricolporitesdigitatus (Jaramillo and Dilcher, 2001) has similar dimensions to Striatricolporites sp. However, S. digitatus has a thicker exine (Jaramillo and Dilcher, 2001: 1 µm) and ectocolpi costate.
Other occurrences. Striatricolporites has been recovered in Mexico from the late Eocene to early Oligocene of Puebla (Martínez-Hernández and Ramírez-Arriaga, 1999). Jaramillo and Dilcher (2001) reported Striatricolporites digitatus from the middle Paleocene of Colombia. Africa - Oligocene of Cameroon (Salard-Cheboldaeff, 1979). Type species from the Paleocene-Eocene of Guyana (Leidelmeyer, 1966).
Genus THOMSONIPOLLIS Krutzsch, 1960 emend. Elsik, 1968
Type species. Thomsonipollismagnificus (Thomson and Plug, 1953) Krutzsch, 1960
Thomsonipollissabinetownensis Elsik, 1968
Figure 5.19
Material. Sample Pb-9334(4), Palynology Laboratory, IGLUNAM.
Description. Monad, isopolar, radiosymmetric, amb circular. Triporate, pores 5 x 4 μm, annulate, annulus 1 µm thick. Exine tectate, 0.5 µm thick, scabrate.
Dimensions. Equatorial diameter 24.8 µm x 28 µm, one specimen measured.
Other occurrences. U.S.A. - Eocene of Gulf Coast (Elsik, 1974)
Order ASTERALES Lindley, 1833
Family ASTERACEAE Berchtold and Presl, 1820
Genus MUTISIAPOLLIS Macphail and Hill, 1994
Type species. Mutisiapollis patersonii Macphail and Hill, 1994
Mutisiapollis sp.
Figure 5.7, 5.11
Material. Sample Pb-8872, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, prolate. Tricolporate, endoaperture lalongate, 12 µm x 4.8 µm. Exine tectate, columellate, clearly stratified with two layers, 3.2 µm thick in the mesocolpi, and 2.4 µm in the apocolpi, supramicroechinate, echinae <0.5 µm high .
Dimensions. Equatorial diameter 24.8 µm, polar diameter 38.4 µm, one specimen measured.
Comparisons. M. viteauensis Barreda et al., 1998 from the Miocene “Serie del Yeso” of Argentina has a thicker exine (Barreda et al., 1998: 3.5 to 6 µm) and spines less than 1 µm high. However, Mutisiapollis sp. from the Cuayuca Formation has lalongate endoapertures with dimensions similar to those of M. viteauensis.
Other occurrences. There are no records of this taxon in Mexico. Mutisieae pollen grains have been recovered from the Miocene Gatun Formation, Panamá (Graham, 1990a) and in Argentina from the Miocene of San Juan Province (Barreda et al., 1998).
Genus TUBULIFLORIDITES Cookson, 1947 ex Potonié, 1960
Type species. Tubulifloridites antipodicus Cookson, 1947 ex Potonié, 1960
Tubulifloridites sp.
Figure 5.17
Material. Samples Pb-9138, Pb-9340, Pb-8896, Pb-8897, Pb-8871, Pb-9141, Pb-8868, Pb-8869, Pb-9136, Pb-8870, and Pb-9147, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, amb circular. Tricolporate. Exine tectate, columellate, 4 µm thick, cavate, echinae columellate at the base, conical, 4 µm long x 3 µm wide at the base, distance between echinae 4-5 µm.
Dimensions. Equatorial diameter 20-23 µm, three specimens measured.
Comparisons. Compositae specimens have been recovered from the Monterrey Formation (Miocene), although they are larger (Srivastava, 1984: 40 µm) with echinae 6 µm high (Srivastava, 1984).
Other occurrences. Compositae specimens have been reported in Mexico - Eocene of Baja California (Cross and Martínez-Hernández, 1980). U.S.A.- late Miocene Valentine Formation, Nebraska (MacGinitie, 1962). Originally described from the Tertiary of Australia (Cookson, 1947) and Germany (Potonié, 1960).
Order CARYOPHYLLALES Bentham and Hooker, 1862
Family AMARANTHACEAE Jussieu, 1789
Genus CHENOPODIPOLLIS Krutzsch, 1966
Type species. Chenopodipollis multiplex (Weyland and Pflug, 1957) Krutzsch, 1966
Chenopodipollis sp.
Figure 3.9
Material. Samples Pb-9147, Pb-9136, Pb-9343, Pb-8869, Pb-8872, Pb-9138, Pb-8898, and Pb-9334, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, apolar, radiosymmetric, spheroidal. Periporate, 22 pores 2.5-3 µm wide, annulate, annulus 1.5 µm thick. Exine tectate, collumelate, 1 µm thick, with microreticulate pattern.
Dimensions. Diameter: 16-25 µm, 10 specimens measured.
Comparison. Our specimens have dimensions and annulus similar to Chenopodipollismultiplex (Bebout, 1980). However, the exine of the Chenopodipollis sp. from the Cuayuca Formation shows a microreticulate pattern, and this trait is not observed in C.multiplex.
Other occurrences. This genus has been reported across the globe since the Paleocene. In Mexico there are records of this genus from the late Eocene to early Oligocene Pie de Vaca Formation, Puebla (Martínez-Hernández and Ramírez-Arriaga, 1999); Oligocene San Gregorio Formation, Baja California Sur (Martínez-Hernández and Ramírez-Arriaga, 2006); Oligocene-Miocene of Chiapas (Biaggi, 1978); and Neogene (Palacios and Rzedowski, 1993; Graham, 1975).
Family NYCTAGINACEAE Jussieu, 1789
Genus MAGNAPERIPORITES González-Guzmán, 1967
Type species. Magnaperiporites spinosus González-Guzmán, 1967
Magnaperiporites sp.
Figure 4.16
Material. Sample Pb-9334, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, apolar, radiosymmetric, spheroidal, periporate. Pores circular or subcircular, 3 µm in width, distance between pores from 8 to 10 µm, pattern areolate. Exine tectate collumelate, 4 µm thick without echinae, perforate supraechinate, echinae 1 µm high.
Dimensions. Diameter 65 µm, one specimen measured.
Comparisons. Magnaperiporites sp. has smaller dimensions than M. spinosus illustrated by González-Guzmán (1967) and Salard-Cheboldaeff (1979). González-Guzmán (1967) gave a diameter for M. spinosus of 90 to 105 µm, with an exine thickness of 13.5 µm. Salard-Cheboldaeff (1979) reported M. spinosus with a diameter of 92-93 µm and exine 13 µm thick.
Other occurrences. This is the first mention of the genus in Mexican fossil microfloras. Magnaperiporites spinosus has been recovered from the Oligocene of Cameroon, Africa (Salard-Cheboldaeff, 1979) and from the early to middle Eocene of Colombia (González-Guzmán, 1967).
Family PLUMBAGINACEAE Jussieu, 1789
Genus ARMERIA Willdenow, 1809
Type species. Armeria vulgaris Willdenow, 1809
Armeria sp.
Figure 3.11
Material. Samples Pb-9340, Pb-8898, Pb-8872, Pb-9137, Pb-9334, and Pb-8896, Palynology Laboratory, IGLUNAM.
Description. Monad, isopolar, radiosymmetric, amb circular, prolate spheroidal. Tricolpate. Exine semitectate, columellate, 5 µm thick, reticulate, supramicroequinate, muri with large columellae of 4.2 µm and lumina 3.6-5.6 µm in diameter.
Dimensions. Equatorial diameter 37-40 µm, polar diameter 35-45 µm, two specimens measured.
Remarks. Armeria is included in Plumbaginaceae. This family is cosmopolitan, although its distribution center is the Mediterranean (Papanicolaou and Kokkini, 1982).
Comparison. The Armeria pollen type is very similar to the Armeria sp. recovered from the Monterey Formation (Miocene) phosphatic facies, California (Srivastava, 1984). Both have similar dimensions and ornamentation.
Other occurrences. Mexico - late Eocene to early Oligocene Pie de Vaca Formation, Puebla (Martínez-Hernández and Ramírez-Arriaga, 1999). U.S.A. - Miocene of the Monterey Formation, California (Srivastava, 1984).
Order FABALES Bromhead, 1838
Family FABACEAE Lindley, 1836
Fabaceae pollen type 1
Figure 3.12-13
Material. Sample Pb-9334, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, subprolate. Tricolporoidate. Exine semitectate, columellate, 1.2 µm thick, microreticulate, lumina 0.5 µm, muri 0.5 µm .
Dimensions. Equatorial diameter 26 µm, polar diameter 32 µm, one specimen measured.
Comparisons. Fabaceae pollen type 1 differs from Fabaceae pollen type 2 and Fabaceae pollen type 3 (in this paper) because it is tricolporoidate and microreticulate whereas Fabaceae pollen type 2 and Fabaceae pollen type 3 are tricolporate and not reticulate.
Other occurrences. Leguminosae fossil pollen grains have been recovered from late Eocene to early Oligocene Pie de Vaca Formation, Puebla (Martínez-Hernández and Ramírez-Arriaga, 1999).
Fabaceae pollen type 2
Figure 3.18-20
Material. Sample Pb-8890, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, prolate. Tricolporate, ectoaperture reaching apocolpia, endoaperture lolongate 3 x 7 µm. Exine tectate, columellate, 1 µm thick, psilate.
Dimensions. Equatorial diameter 18µm, polar diameter 30 µm, one specimen measured.
Comparisons. The colporate and tectate - psilate condition makes this specimen different from Fabaceae pollen type 1, and Fabaceae pollen type 3 (of the same association, described in this paper).
Other occurrences. Fabaceae fossil pollen grains have been recovered from late Eocene to early Oligocene Pie de Vaca Formation, Puebla (Martínez-Hernández and Ramírez-Arriaga, 1999).
Fabaceae pollen type 3
Figure 4.13
Material. Sample Pb-9334, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, amb triangular. Tricolporate. Exine tectate, 1µm thick, foveolate.
Dimensions. Equatorial diameter 25 µm, one specimen measured.
Comparisons. Observations on morphology was only achievable in polar view, thus it was not possible to describe the endoaperture. Nevertheless, it differs from other Fabaceae described in this contribution, because it is tectate-foveolate.
Other occurrences. Fabaceae fossil pollen grains ( Caesalpinia, Acacia, and Mimosa ) have been recovered from late Eocene to early Oligocene Pie de Vaca Formation, Puebla (Martínez-Hernández and Ramírez-Arriaga, 1999).
Genus MARGOCOLPORITES Ramanujam, 1966 ex Srivastava, 1969 emend. Pocknall and Mildenhall, 1984
Type species. Margocolporites tsukadai Ramanujam 1966 ex Srivastava, 1969
Margocolporites aff. vanwijhei Germeraad et al., 1968
Figure 4.6
Material. Samples Pb-9334(1) and Pb-9334(4), Palynology Laboratory, IGLUNAM.
Description. Monad, isopolar, radiosymmetric, amb circular. Tricolporate, exoapertures present colpal membrane with fine baculae and margo 1 µm thick, endoaperture lolongate 2.4 µm X 5.6 µm. Exine subtectate, collumelate, 1.2 µm thick, reticulate, heterobrochate, lumina 0.8- 1.5 µm, simplicolumelate muri.
Dimensions. Equatorial diameter 34.3 (38) 44 µm, polar diameter34 µm, four specimens measured.
Other occurrences. Mexico - Oligocene San Gregorio Formation, Baja California Sur (Martínez-Hernández and Ramírez-Arriaga, 2006). Originally described from the Paleogene of the Caribbean region and Nigeria (Germeraad et al., 1968). South America - late Oligocene to Miocene Chenque Formation (Barreda, 1997c); early Miocene Chinches Formation (Ottone et al., 1998); and middle Miocene “Serie el Yeso,” Argentina (Barreda et al., 1998).
Margocolporites sp.
Figure 4.4-5
Material. Sample Pb-9340, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, amb circular. Tricolporate, colpi with membrane bearing fine bacula. Exine semitectate, columellate, 0.8 µm thick, reticulate, heterobrochate, lumina 1-2 µm.
Dimensions. Equatorial diameter 45 µm, one specimen measured.
Comparisons. This taxon is similar to Margocolporites sp. 2 from central Colombia (Jaramillo and Dilcher, 2001); but, Margocolporites sp. 2 is larger than Margocolporites sp. from the Cuayuca Formation. Margocolporites sp. (this work) has a larger equatorial axis and larger brochi than M. vanwijhei (Germeraad et al., 1968). The diameter of the endoaperture is almost equal to the width of the ectoaperture in the Margocolporites sp. from the Cuayuca Formation. The diameter of the endoaperture is smaller than the width in M. vanwijhei (Macphail, 1999; Eisawi and Schrank, 2008).
Other occurrences. The genus Margocolporites has been recovered from late Eocene to early Oligocene Pie de Vaca Formation, Puebla (Martínez-Hernández and Ramírez-Arriaga, 1999); Oligocene San Gregorio Formation, Baja California Sur (Martínez-Hernández and Ramírez-Arriaga, 2006). M. vanwijhei has been recovered from middle Paleogene of central Colombia (Jaramillo and Dilcher, 2001); Germeraad et al. (1968) reported this species from late Eocene of Nigeria and in the Caribbean region Graham (2010) reported it from Eocene to the present time. It has also been recovered from the Murray Basin of southeastern Australia (Macphail, 1999) and southeastern Sudan (Eisawi and Schrank, 2008).
Genus POLYADOPOLLENITES Thomson and Pflug, 1953
Type species. Polyadopollenites multipartitus Thomson and Pflug, 1953
Polyadopollenites sp.1
Figure 5.2-3
Material. Sample Pb-9334, Palynology Laboratory, IGLUNAM.
Description. Polyad with 16 pollen grains: eight central grains and eight peripheral. Inaperturate. Exine atectate, 0.8-1.6 µm thick, regulate.
Dimensions. Polyad 48.8 µm x 46.4 µm. Monad: Equatorial diameter 16-20 µm, polar diameter 11.2 -14.4 µm, one specimen measured.
Comparisons. Polyadopollenites sp. 1 is distinguished by its suprarrugulate exine instead of a psilate tectum in Acaciapollenites from late Oligocene of New Zealand (Pocknall and Mildenhall, 1984). Polyadopollenites sp. 1 presents similar ornamentation to Polyadopollenitesmulleri (Cavagnetto and Guinet, 1994); however, Polyadopollenites sp. 1 is not as big as P. mulleri (Cavagnetto and Guinet, 1994: 65-72 µm). Furthermore, P. mulleri from the Oligocene of Spain has porate monads (Cavagnetto and Guinet, 1994).
Other occurrences. This genus has been recovered in Mexico from the late Eocene to early Oligocene Pie de Vaca Formation (Martínez-Hernández and Ramírez-Arriaga, 1999); Oligocene San Gregorio Formation, Baja California Sur (Martínez-Hernández and Ramírez-Arriaga, 2006); Oligo-Miocene of Chiapas (Biaggi, 1978); Neogene (Palacios and Rzedowski, 1993). Central America - early Miocene La Culebra Formation, Panamá (Graham, 1988). Caribbean islands- Oligocene San Sebastián Formation, Puerto Rico (Graham and Jarzen, 1969). Originally described from the Eocene-Oligocene in Germany (Thomson and Pflug, 1953).
Polyadopollenites sp. 2
Figure 5.1
Material. Sample Pb-9334, Palynology Laboratory, IGLUNAM.
Description. Polyad with 16 pollen grains: eight central grains and eight peripheral. Syncolpate. Exine tectate columellate, 0.8-1.6 µm thick, foveolate.
Dimensions. Polyad 36.8 µm, one specimen measured.
Comparisons. Polyadopollenites sp. 2 has distinctively syncolpate, tectate and foveolate monads; in contrast, Acaciapollenites has inaperturate, tectate and psilate monads (Pocknall and Mildenhall, 1984). This taxon does not have a superficial reticulum as Polyadopollenitescooksonii (Cavagnetto and Guinet, 1994); furthermore, Polyadopollenites sp. 2 has syncolpate apertures instead of subsidiary apertures.
Other occurrences. Polyadopollenites sp. 2 shows the same geographic pattern as Polyadopollenites sp. 1.
Family POLYGALACEAE Hoffmannsegg and Link, 1809
Genus POLYGALACIDITES Sah and Dutta, 1966
Type species. Polygalacidites clarus Sah and Dutta, 1966
Polygalacidites sp.
Figure 5.16
Material. Sample Pb-9340, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, subprolate. Stephanocolporate, endoaperture lalongate with costae. Exine atectate, 1.2 µm thick, psilate.
Dimensions. Equatorial diameter 11 µm, polar diameter 20 µm, one specimen measured.
Comparisons. This taxon differs from the Polygalacidites sp. from the Neogene of southeastern Sudan (Eisawi and Schrank, 2008) as the latter has more numerous and larger ectoapertures than the Polygalacidites sp. from the Cuayuca Formation. Nevertheless, both taxa have almost the same polar diameter.
Other occurrences. Mexico - late Eocene - early Oligocene (Martínez-Hernández, 1999), Oligocene San Gregorio Formation (Martínez-Hernández and Ramírez-Arriaga, 2006). Originally described from Paleogene in Assam, India (Sah and Dutta, 1966).
Order FAGALES Engler, 1892
Family CASUARINACEAE Brown, 1814
Genus CASUARINIDITES Cookson and Pike, 1954
Type species. Casuarinidites cainozoicus Cookson and Pike, 1954
Casuarinidites sp.
Figure 3.4
Material. Samples Pb-9337, Pb-9340, Pb-8898, Pb-9141, Pb-9147, Pb-9136, Pb-9339, and Pb-8896, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, amb triangular. Triporate, angulaperturate, aspidate pores, annulate, annulus with a 5.6 μm diameter. Exine tectate, granulate, 1.2 μm thick, scabrate. The granular infratectal structure gives a microreticulate pattern.
Dimensions. Equatorial diameter 26-37 µm, two specimens measured.
Comparisons. The size of the Casuarinidites sp. from the Cuayuca Formation is similar to that of C. granilabrata described by Srivastava (1972); however the exine of C. granilabrata is 2 µm thick.
Other occurrences. This genus has been reported in Mexico from the Oligocene San Gregorio Formation (Martínez-Hernández and Ramírez-Arriaga, 2006); Oligo-Miocene of Chiapas (Biaggi, 1978). U.S.A. - Miocene, Massachussets (Frederiksen, 1984); Paleocene, Alabama (Srivastava, 1972).
Family JUGLANDACEAE de Candolle ex Perleb, 1818
Genus MOMIPITES Wodehouse, 1933
Type species. Momipites coryloides Wodehouse, 1933
Momipites coryloides Wodehouse, 1933
Figure 4.12
Material. Samples Pb-9138’(1), Pb-9334(4) and Pb-9340(2), Palynology Laboratory, IGLUNAM.
Description. Monad, isopolar, radiosymmetric, amb triangular. Triporate, equidistant zonate pores. Exine tectate, 1 µm thick with uniformly spaced spinules.
Dimensions. Equatorial diameter 14 (17.8) 20 µm, five specimens measured.
Other occurrences. Mexico - late Eocene to early Oligocene Pie de Vaca Formation (Martínez-Hernández and Ramírez-Arriaga, 1999); Oligocene San Gregorio Formation (Martínez-Hernández and Ramírez-Arriaga, 2006); Oligo-Miocene of Chiapas (Langenheim et al., 1967). U.S.A. - late Cretaceous to Paleocene, Colorado (Newman, 1965); late Paleocene, South Carolina (Van Pelt et al., 2000); late Eocene, Yazoo Clay, Mississippi (Tschudy and van Loenen, 1970); late Eocene, Mississippi and Alabama (Frederiksen, 1980a); Paleocene - late Eocene, South Carolina (Frederiksen, 1980b); Eocene, Mississippi embayment (Tschudy, 1973); Miocene, Massachusetts (Frederiksen, 1984); early Eocene, Tennessee (Elsik and Dilcher, 1974). Central America - Oligocene San Sebastión Formation, Puerto Rico (Graham and Jarzen, 1969); Eocene Gatuncillo Formation, Panamá (Graham, 1985); early Miocene Cucaracha Formation and La Boca Formation (Graham, 1988); Mio-Pliocene of Haiti (Graham, 1990b). Europe - Eocene, England (Wilkinson and Boulter, 1980).
Momipites tenuipolus Anderson, 1960
Figure 4.11
Material. Samples Pb-8872(2), Pb-8898(3) and Pb-9334(4), Palynology Laboratory, IGLUNAM.
Description. Monad, isopolar, radiosymmetric, amb triangular. Triporate. Exine tectate, psilate, 0.8 µm thick with a polar thinning.
Dimensions. Equatorial diameter 16.8 (17.3) 18 µm, based upon three measured specimens.
Other occurrences. U.S.A. - early Paleogene, San Juan Basin, New Mexico (Anderson, 1960); late Paleocene Silverado Formation, California (Gaponoff, 1984); Paleocene, Virginia (Frederiksen, 1979, 1998); early Eocene, Gulf Coast (Frederiksen, 1980b).
Order GARRYALES Lindley, 1846
Family EUCOMMIACEAE Engler, 1907
Genus EUCOMMIA Oliver, 1890
Type species. Eucommia ulmoides Oliver, 1890
Eucommia pollen type
Figure 3.16
Material. Samples Pb-9138, Pb-8898, Pb-9340, Pb-8896, and Pb-9334, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, amb circular, subprolate. Tricolporoidate, with two large colpi: one measuring 24 μm long and a smaller one 17.6 µm long. Exine atectate, 0.8 - 1 µm thick, psilate.
Dimensions. Equatorial diameter 17-30 µm, polar diameter 22-35 µm, 13 specimens measured, five specimens measured.
Comparison. Eucommia sp. from the Cuayuca Formation has similar dimensions to E. ? leopoldae (Frederiksen et al. , 1983) and the Eucommia type (Frederiksen, 1988), but differs from both because they are tricolporate whereas the Eucommia herein described is distinctively tricolporoidate as seen in the Eucommia described by Erdtman (1972). It is also similar to Eucommia cf. E. ulmoides illustrated by Leopold (1974, plate 39, figure 13) and Wingate (1983, plate 2, figure 10).
Other occurrences. This genus was recorded in Mexico from the late Eocene to early Oligocene Pie de Vaca Formation, Puebla (Martínez-Hernández and Ramírez-Arriaga, 1999); Oligocene of the San Gregorio Formation (Martínez-Hernández and Ramírez-Arriaga, 2006). U.S.A. - middle Eocene of Kisinger, Wyoming (Leopold, 1974); Eocene, Nevada (Wingate, 1983); early Eocene into middle Eocene, Gulf Coast (Frederiksen, 1988); middle Eocene, Hominy Peak Formation, San Diego, California (Frederiksen, 1983); Eocene, Teton Range, Wyoming (Love et al. , 1978); late Oligocene of Florissant Formation, Colorado (Leopold and Scott, 2001).
Order GENTIANALES Lindley, 1833
Family APOCYNACEAE Jussieu, 1789
Genus LANDOLPHIA Palisot de Beauvois, 1806
Type species. Landolphia owariensis Palisot de Beauvois, 1806
Landolphia pollen type
Figure 5.4
Material. Sample Pb-9334, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, oblate spheroidal. Tricolporate, endoaperture with costae converging with meridional edges of endoapertures indistinct (El Ghazali, 1990). Exine semitectate, columellate, 0.8-1 µm thick, reticulate.
Dimensions. Equatorial diameter 27-33 µm, polar diameter 27-34 µm, two specimens measured. Equatorial diameter in polar view 27-29 µm, two specimens measured.
Comparisons. Landolphia comorensis Benth. and Hook has similar breviectoapertures as the Landolphia pollen type from Cuayuca Fm., although L. comorensis has endoapertures with transverse converging edges and indistinct meridional edges (Erdtman, 1972). The Landolphia pollen type herein described has endoapertures with converging transverse costae and indistinct meridional edges.
Other occurrences . This is the first report of the genus in Mexican fossil associations. Apocynaceae pollen grains have been reported from late Eocene to early Miocene from Cameroon (Salard-Cheboldaeff, 1981).
Family RUBIACEAE Jussieu, 1789
Genus SABICEA Aublet, 1775
Type species. Sabicea cinerea Aublet, 1775
Sabicea pollen type
Figure 5.20
Material. Sample Pb-9334, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, amb circular. Triporate, lolongate pore. Exine semitectate, columellate, 2 µm thick, microreticulate, simplicolumellate.
Dimensions. Equatorial diameter 27 μm, one specimen measured.
Comparisons. Pollen grains of the genus Sabicea, with finely reticulate exine and dimensions ranging from 32 to 36 μm, have been reported for the Miocene of Panama (Graham, 1987); however, the Sabicea pollen type recovered from the Cuayuca Formation is smaller (27 μm in equatorial diameter).
Other occurrences. Central America - early Miocene La Culebra Formation, Panama (Graham, 1987). There are no previous records of this genus in Mexico.
Order MALPIGHIALES Martius, 1835
Family EUPHORBIACEAE Jussieu, 1789
Genus CLAVAINAPERTURITES Hammen and Wymstra, 1964
Type species. Clavainaperturitesclavatus Hammen and Wymstra, 1964
Clavainaperturites sp.
Figure 4.14
Material. Sample Pb-9147, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, apolar, radiosymmetric, spheroidal. Inaperturate. Exine tectate, 2.4 µm thick, sculture baculate with crotonoid pattern showing groups of five to six supratectal bacula.
Dimensions. Diameter 55 x 52 µm, one specimen measured .
Comparisons. Clavainaperturites sp. is very similar to the C. cf. clavatus (35 µm) from the Nuer Formation, Southeast Sudan (Eisawi and Schrank, 2008), although the specimen from the Cuayuca Formation is larger than the taxon from Sudan.
Other occurrences. There are no previous records of this taxon in Mexico. This genus has been reported in the Paleocene of Southeast Sudan (Eisawi and Schrank, 2008); and in the Oligocene of British Guyana (Van der Hammen and Wymstra, 1964).
Genus GLYCYDENDRON Ducke, 1922
Type species. Glycydendron amazonicum Ducke, 1922
Glycydendron pollen type
Figures 3.14-15
Material. Sample Pb-9137, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, amb circular. Tricolpate, colpi with irregular margin. Exine intectate, 1.2-1.5 µm thick, echinate, spines 1.2-2 µm high are very thick at their bases in contrast with the thin flexible apices.
Dimensions. Equatorial diameter 31 µm, one specimen measured.
Comparisons. Glycydendron pollen type is intectate, echinate, and similar to the specimen described from the Miocene Uscari Formation (Graham, 1987), but it is of smaller dimensions. In contrast, the Glycydendron pollen type from the Cuayuca Formation has echinae with flexible apices which are thinner than those described by Graham (1987) from the Miocene of Costa Rica.
Other occurrences. Central America - Miocene Uscari Formation, Costa Rica (Graham, 1987). There are no previous records of the genus in Mexico.
Family LINACEAE de Candolle ex Perleb, 1818
Genus LINUM Linnaeus, 1753
Type species. Linum usitatissimum Linnaeus, 1753
Linum pollen type
Figure 4.15
Material. Sample Pb-9334, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, amb subcircular. Tricolpate, the broad and large colpi make a small polar area. Exine semitectate columellate, 3 µm thick, microreticulate to striate, supragemmate, gemmae 2-3.5 µm in diameter, distributed irregularly over the surface of the grain.
Dimensions. Equatorial diameter 48-53 µm, one specimen measured.
Comparisons. Tricolpate pollen grain characterized by large size (Wingate, 1983: 44 x 49 µm) and gemmate ornamentation has been reported from the Elko Formation (Wingate, 1983, plate 2, figure 12). The tricolpate pollen type 1 from the Elko Formation is not semitectate microreticulate like the Linum pollen type from the Cuayuca Formation, and has a thinner exine.
Other occurrences. This is the first mention of the genus in the Mexican fossil record. This taxon is also recorded from the European middle Eocene to early Oligocene, Ebro Basin, Spain (Cavagnetto and Anadón, 1996).
Family MALPIGHIACEAE Jussieu, 1789
Genus MALPIGHIACEOIDITES Takahashi and Jux, 1989
Type species. Malpighiaceoidites periporifer Takahashi and Jux, 1989
Malpighiaceoidites sp.
Figures 5.6, 5.10
Material. Sample Pb-9334, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, apolar, radiosymmetric, spheroidal. Perisyncolporate, six to eight pores distributed irregularly and joined with colpi. Exine atectate, 1.5-2 µm thick, supraverrucate, verrucae irregular supratectal, 0.5-4 µm wide.
Dimensions. Diameter 25 (29.9) 33.6 µm, five specimens measured.
Comparisons. The Malpighiaceoidites sp. has pores connected by thin colpal bands, like the Type A described by Martin (2002); nevertheless, the Malpighiaceoidites sp. exhibits verrucae bigger than those presented by Type A of Martin (2002). Malpighiaceae specimens recovered in Central America by Graham (1985, 1988, 1989, 1990b) are different from those of the Cuayuca Formation. Perisyncolporites pokornyi reported from the Miocene of Venezuela (Lorente, 1986) and from the Río Turbio Formation (Eocene) from Argentina (Fernández et al., 2012) has a thicker exine without the distinctive verrucae present in the Cuayuca Formation specimen.
Other occurrences. The genus Malpighiaceoidites has been recovered only from the Miocene of Venezuela (Lorente, 1986), Paleocene of São Paulo, Brazil (Lima et al., 1991), Eocene of Argentina (Fernández et al., 2012), and Tertiary rocks from southeastern Australia (Martin, 2002).
Genus PERISYNCOLPORITES Germeraad et al., 1968
Type species. Perisyncolporites pokornyi Germeraad et al., 1968
Perisyncolporites sp.
Figure 5.5
Material. Sample Pb-9334, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, apolar, radiosymmetric, spheroidal. Perisyncolporate, colpi with one or two endoapertures with irregular distribution, ora 2.5-4 µm in diameter, with costae. Exine tectate, 0.8-1 µm thick, suprarrugulate.
Dimensions. Diameter 34.4 µm x 32 µm, one specimen measured.
Comparisons. This taxon resembles Perisyncolporitespokornyi from the middle Eocene of Florida, U.S.A. (Jarzen et al., 2010), although P. pokornyi has a thicker exine (Jarzen et al., 2010: 2.5 - 3 µm) and a smaller size (23-26 µm).
Other occurrences. U.S.A. - Middle Miocene, Florida (Jarzen and Klug, 2010). This genus has also been reported from the Paleogene of the Caribbean region and Nigeria (Germeraad et al. , 1968; Graham, 1990b) and the Paleogene of South America (Jaramillo and Dilcher, 2001).
Order MALVALES Lindley, 1833
Family MALVACEAE Jussieu, 1789
Genus BOMBACACIDITES Couper, 1960
Type species. Bombacaciditesbombaxoides Couper, 1960
Bombacacidites sp.
Figure 3.23
Material. Samples Pb-9334 and Pb-9340, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, amb tringular obtuse. Tricolporate, apertures interangular with exinal thickenings. Exine semitectate, columellate, 0.8 μm thick, reticulate, heterobochate, apocolpial lumina of 1.6 μm are larger than mesocolpial lumina <0.8 μm.
Dimensions. Equatorial diameter 21.2 µm, based on one measured specimen.
Comparisons. The Bombacacidites sp. (in this contribution) is similar in size (16-20 µm) to Bombacacidites sp. 2 in Frederiksen (1988), although the Bombacacidites sp. from the Cuayuca Formation has wider brochi (0.8 - 1.6 µm) than Bombacacidites sp. 2 from the eastern Gulf Coast (Frederiksen, 1988: 0.5 - 1 µm). The specimen herein studied is also similar in the thickness of the exine and the width of the luminae at apocolpia to Bombacacidites sp. 2 described by Jaramillo and Dilcher (2001) from the middle Paleogene of Colombia, but differs in having a smaller equatorial diameter (Bombacacidites sp. of Cuayuca: 21.2 µm; Bombacacidites sp. 2: 33 to 40 µm). Another similar specimen to the Bombacacidites sp. from the Cuayuca Formation was described as middle Eocene at the Kissinger Lakes locality (Leopold, 1974, plate 45, figures 2-3).
Other occurrences. This genus has been recorded in Mexico from the late Eocene to early Oligocene Pie de Vaca Formation, Puebla (Martínez-Hernández and Ramírez-Arriaga, 1999). U.S.A. - middle Eocene, San Diego, California (Frederiksen et al. , 1983); Eocene and Eocene-Oligocene of the Gulf Coast (Elsik, 1974; Frederiksen, 1988); middle Eocene, Kisinger, Wyoming (Leopold, 1974); middle Eocene from Pine Island, Florida, U.S.A. (Jarzen and Klug, 2010).
Order MYRTALES Lindley, 1833
Family ONAGRACEAE Jussieu, 1789
Genus CORSINIPOLLENITES Nakoman, 1965
Type species. Corsinipollenites oculus-noctis (Thiergart, 1940) Nakoman, 1965
Corsinipollenites sp. 1
Figure 4.1
Material. Samples Pb-9343, Pb-9340, and Pb-9334, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, amb triangular. Triporate, annulate, annulus 7 μm thick, pores aspidate with granulate structure. Exine tectate, 1-1.6 μm thick, psilate, with microreticular endopattern.
Dimensions. Equatorial diameter 24-38.4 µm, four specimens measured.
Comparisons. Corsinipollenites. sp. 1 (in this paper) shows smaller dimensions and more aspidate pores compared to Corsinipollenites sp. 2 and Corsinipollenites sp. 3 of the Cuayuca assemblage. Corsinipollenites sp.1 has a more protruding pore and annuli thicker than those reported by Jaramillo and Dilcher (2001) which have annulus 2 μm thick, as well as that recovered from middle Eocene of Florida, with annulus 2-4 μm wide (Jarzen and Klug, 2010).
Other occurrences. In Mexico, the genus is reported to be from late Eocene to early Oligocene Pie de Vaca Formation (Martínez-Hernández and Ramírez-Arriaga, 1999); Oligocene San Gregorio Formation (Martínez-Hernández and Ramírez-Arriaga, 2006). Jaramillo and Dilcher (2001) reported Corsinipollenites in the Paleocene of Colombia; it is also mentioned in the Paleocene of Merida Andes, western Venezuela (Pocknall and Jarzen, 2009) and in the early to middle Eocene from Pine Island, Florida (Jarzen and Klug, 2010) and Oligocene of Germany (Thiergart, 1940). More data on the distribution of this genus in the U.S.A. and northern South America is in Jarzen and Klug (2010).
Corsinipollenites sp. 2
Figure 4.3
Material. Sample Pb-9334, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, amb triangular.Triporate, pores aspidate. Exine tectate, 0.8 μm thick, psilate.
Dimensions. Equatorial diameter 32.8 x 71.2 µm, one specimen measured.
Comparisons. Corsinipollenites sp. 2 is different from Corsinipollenites sp. 1 and Corsinipollenites sp. 3 of the same association, because it is larger, with a thinner exine and pores that are not as aspidate as in the other species of Corsinipollenites described in this contribution. This taxon is larger and has a thinner exine than Corsinipollenitesatlantica from Argentina (Barreda and Palamarczuk, 2000b) and Corsinipollenites cf. psilatus from the Melot Basin, southeast Sudan (Eisawi and Schrank, 2008).
Corsinipollenites sp. 3
Figure 4.2
Material. Sample Pb-9334, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, amb triangular. Triporate, pores aspidate. Exine tectate, granulate, 0.8-1 μm thick, psilate.
Dimensions. Equatorial diameter 41-50 µm, three specimens measured.
Comparisons. This species of Corsinipollenites exhibits a characteristic pore with evident granulate structure. Although granulate structure is present in Corsinipollenites sp. 1 and Corsinipollenites sp. 2, more aspidate pores are present in Corsinipollenites sp. 1; Corsinipollenites sp. 2 presents less aspidate pores than the other two species described. Corsinipollenites sp. 3 has similar dimensions and annulus thickness to Corsinipollenitesatlantica described from the Argentinian Oligocene-Miocene by Barreda and Palamarczuk (2000b); nevertheless, the exine of C. atlantica is thicker. This taxon closely resembles Corsinipolleites cf. psilatus from the Melot Basin, southeast Sudan (Eisawi and Schrank, 2008).
Order RANUNCULALES Lindley, 1833
Genus RANUNCULACIDITES Van der Hammen, 1956 ex Van der Hammen and Wijmstra, 1964
Type species. Ranunculacidites operculatus (Van der Hammen and Wijmstra, 1964) Jaramillo and Dilcher, 2001
Ranunculacidites cf. communis Sah, 1967
Figure 5.13
Material. Samples Pb-9334 and Pb-9340, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, oblate spheroidal. Tricolporate, pantoperculate. Exine semitectate, columellate, 0.8 μm thick, microreticulate, brochi 0.8 μm.
Dimensions. Equatorial diameter 23 µm, polar diameter 23 µm, one specimen measured.
Comparisons. Ranunculaciditescommunis from the late Neogene of Africa (Sah, 1967) differs from R. operculatus from the middle Miocene of Florida (U.S.A.) in having a fine reticulum instead of a psilate exine (Jarzen and Klug, 2010). Ranunculaciditescommunis (Sah, 1967) differs from R. cf. communis by having shorter colpi and a thick plug pontoperculum.
Other occurrences. There are no previous records of this taxon in Mexico. Ranunculaciditesoperculatus has been recovered from the middle Miocene of Florida (Jarzen and Klug, 2010) and from the Paleogene of central Colombia (Jaramillo and Dilcher, 2001).
Ranunculacidites operculatus
Figure 5.21
Material. Sample Pb-9334(4), Palynology Laboratory, IGLUNAM.
Description. Monad, isopolar, radiosymmetric, amb oval triangular. Tricolpate, pontoperculate. Exine tectate, 0.8 μm thick, psilate.
Dimensions. Equatorial diameter 20 µm based on one measured specimen.
Other occurrences. Mexico - Oligocene to Miocene of Chiapas (Biaggi, 1978); Pliocene of Veracruz (Graham, 1975). Central America - early Miocene La Culebra Formation (Graham, 1988); early Miocene of Uscari, Costa Rica (Graham, 1987); Pleistocene of Panamá (Bartlett and Barghoorn, 1973); Pleistocene of El Salvador and Guatemala (Tsukada and Deevey, 1967); early Miocene La Boca Formation, Panamá (Graham, 1989). Carribean islands - Oligocene of Puerto Rico (Graham and Jarzen, 1969); late Miocene to Pliocene of Haiti (Graham, 1990b). South America - Eocene and Miocene of Venezuela (Lorente, 1986); Oligocene to early Miocene of Venezuela (Rull, 2001, 2003); middle Eocene of Tibu, Colombia (Gonzalez-Guzman, 1967); Cenozoic of Brazil (Regali et al., 1974); late Oligocene to Miocene of Santa Cruz Province, Argentina (Barreda and Palamarczuk, 2000a); Tertiary of Colombia and British Guyana (Van der Hammen and Wymstra, 1964).
Order ROSALES Lindley, 1833
Genus RHAMNACEAEPOLLENITES Thielle-Pfeiffer, 1980
Type species. Rhamnaceaepollenites triquetrus Thielle-Pfeiffer, 1980
Rhamnaceaepollenites sp.
Figure 5.22
Material. Sample Pb-9334, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, oblate spheroidal. Tricolporate, endoapertures 5 x 4 µm with margo. Exine semitectate, columellate, 0.8 µm thick, microreticulate to tectate perforate, heterobrochate, brochi less than 1 μm.
Dimensions. Equatorial diameter 13 µm, polar diameter 13-19 µm, two specimens measured.
Comparisons. Rhamnaceaepollenites sp. has dimensions similar to the type species R. triquetrus (Jansonius and Hills, 1976: 18-20 µm), and presents a faint to distinct reticulate exine (Jansonius and Hills, 1976: card number 3904) which is considered heterobrochate.
Other occurrences. There are no previous records of this taxon in Mexico. This genus has been reported from the Miocene of Turkey (Kayseri and Akgün, 2008).
Family ULMACEAE Mierbel, 1815
Genus ULMIPOLLENITES Wolff, 1934
Type species. Ulmipollenites undulosus Wolff, 1934
Ulmipollenites sp.
Figure 5.18
Material. Samples Pb-9137, Pb-9343, Pb-8896, Pb-8872, Pb-9141, Pb-9138, Pb-8898, Pb-9339, Pb-9340, and Pb-9147, Palynology Laboratory, IGLUNAM.
Description. Monad pollen, isopolar, radiosymmetric, amb subcircular. Tetraporate or pentaporate. Exine tectate, 1 µm thick, suprarugulate.
Dimensions. Equatorial diameter 17-33 µm, five specimens measured.
Comparisons. Considering dimensions and number of pores, Ulmipollenites sp. is similar to U. krempii from the Eocene of California, U.S.A. (Frederiksen et al., 1983) and U. undulosus (Jansonius and Hills, 1976: card number 3121); but the rugulae are less conspicuous in our specimens.
Other occurrences. Mexico - Ulmipollenites has been recovered from the late Eocene to early Oligocene Pie de Vaca Formation (Martíne--Hernández and Ramírez-Arriaga, 1999); Oligocene San Gregorio Formation (Martínez-Hernández and Ramírez, 2006); Oligo-Miocene of Chiapas (Biaggi, 1978); Neogene (Palacios and Rzedowski, 1993); Pliocene, Paraje Solo, Veracruz (Graham, 1975). U.S.A. - late Miocene Valentine Formation, Nebraska (MacGinitie, 1962); late Eocene, Jackson Group, Mississippi and Alabama (Frederiksen, 1980a); Eocene of California (Frederiksen, 1989) and middle Miocene of Florida (Jarzen and Klug, 2010). Initially found in the Tertiary of Germany by Wolff (1934) and Thomson and Plug (1953).
DISCUSSION
The Cuayuca Formation yields rich and diverse palynoflora composed of 97 taxa. Spores are represented by 13.4% (n=13) and gymnosperms 7.2% (n= 7) of the taxa, whereas angiosperms are the most diverse plant group, represented by 76.3% (n= 74) with dominance of Dicotyledonae (66%; n=64) over the Monocotyledonae (10.3%; n=10). Finally, algae are also present with 3.1% (n=3). The present contribution deals with the taxonomy and systematics of selected angiosperm pollen grains (Monocotyledonae and Dicotyledonae) recovered from the Cuayuca Formation (Table). The Monocotyledonae pollen grains are represented by monosulcate and monoporate aperture types; on the other hand, Dicotyledonae pollen grains include colpate, colporate, colporoidate, porate and inaperturate pollen types.
Vegetation types represented in the Cuayuca Formation palynoflora are suggested as a result of comparison between botanical affinities of fossil taxa and the most representative plants reported for current vegetation (Rzedowsky, 2006; Fernández-Nava et al., 1998), as well as comparison between percentages from fossil assemblages against current pollen rain (Velázquez, 2008; Palacios and Rzedowsky, 1993). In general, the palynoflora from the Cuayuca Formation includes taxa belonging to semiarid vegetation such as tropical deciduous forest, chaparral, grassland, and arid tropical scrub, in which angiosperms are one of the main representatives (Table); besides, two temperate vegetation types have been suggested: Pinus forest and cloud forest.
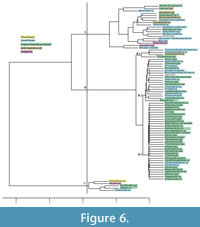 It is important to mention that in the analysis of fossil pollen association, several taphonomic factors must be considered. Since pollen assemblages represent the admixture of a number of plant communities in accordance with their distribution, type of pollen dispersal, biological habit, and proximity to the depositional area, their representativeness will vary greatly in any given space. Despite the previous consideration, hierarchical clustering analysis supports the hypothesis that several plant communities existed at the time of deposition (Figure 6). This analysis was carried out considering the total count of each taxon in order to know whether plant communities were grouped independently. Cluster analysis shows highlighted taxa according to the vegetation type in which they are usually found (Rzedowsky, 2006). In general, three groups are shown in the cladogram. First of all, node A groups four types of vegetation, of which most taxa belong to cloud forest, although elements of tropical deciduous forest, arid tropical scrub, and chaparral are also included. On the other hand, even though most of the taxa included in node B are currently common in tropical deciduous forest (subnodes B-2 and B-3), there are also representatives of cloud forest grouped principally in subnodes B-1 and B2, besides two taxa in B-3. Finally, node C contains Pinus forest and scarce taxa from chaparral, cloud forest, and tropical deciduous forest (Figure 6).
It is important to mention that in the analysis of fossil pollen association, several taphonomic factors must be considered. Since pollen assemblages represent the admixture of a number of plant communities in accordance with their distribution, type of pollen dispersal, biological habit, and proximity to the depositional area, their representativeness will vary greatly in any given space. Despite the previous consideration, hierarchical clustering analysis supports the hypothesis that several plant communities existed at the time of deposition (Figure 6). This analysis was carried out considering the total count of each taxon in order to know whether plant communities were grouped independently. Cluster analysis shows highlighted taxa according to the vegetation type in which they are usually found (Rzedowsky, 2006). In general, three groups are shown in the cladogram. First of all, node A groups four types of vegetation, of which most taxa belong to cloud forest, although elements of tropical deciduous forest, arid tropical scrub, and chaparral are also included. On the other hand, even though most of the taxa included in node B are currently common in tropical deciduous forest (subnodes B-2 and B-3), there are also representatives of cloud forest grouped principally in subnodes B-1 and B2, besides two taxa in B-3. Finally, node C contains Pinus forest and scarce taxa from chaparral, cloud forest, and tropical deciduous forest (Figure 6).
The existence of Pinus forest ( Pinuspollenites: 43.6 - 55.4%) and cloud forest with abundance of Engelhardia - Alfaroa - Oreomunnea ( Momipites: 10.5 - 16.7%) during late Eocene - early Oligocene have also been corroborated by means of comparison between percentages in Cuayuca Formation pollen fossil assemblages and current pollen rain (Velázquez, 2008; Palacios and Rzedowsky, 1993).
The palynoflora is interpreted to have been deposited in a shallow lacustrine environment under semiarid conditions. The local semiarid vegetation surrounding the lake included grassland, arid tropical scrub, and tropical deciduous forests. Upland plant communities are represented by Pinus forest and cloud forest (Ramírez-Arriaga et al., 2006, 2008).
PAE Analysis between Cuayuca Formation Sections
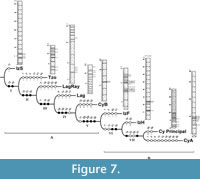 PAE analysis included the presence/absence of 44 palynomorphs (Appendix 3) and resulted in one cladogram in which L= 89, CI = 0.50 and RI = 0.54 (Figure 7). Upon analysis, the cladogram shows that the base of the tree -node I- groups Tzompahuacan (Tzo) with all other clades due to the presence of Liliacidites (13) and Tubulifloridites (42). Level II clusters Lagunillas de Rayón (LagRay) with node III sections based on the presence of Alnus vera (15), Quercoidites (31) and Rhoipites aff. cryptoporus (32).
PAE analysis included the presence/absence of 44 palynomorphs (Appendix 3) and resulted in one cladogram in which L= 89, CI = 0.50 and RI = 0.54 (Figure 7). Upon analysis, the cladogram shows that the base of the tree -node I- groups Tzompahuacan (Tzo) with all other clades due to the presence of Liliacidites (13) and Tubulifloridites (42). Level II clusters Lagunillas de Rayón (LagRay) with node III sections based on the presence of Alnus vera (15), Quercoidites (31) and Rhoipites aff. cryptoporus (32).
Cupressacites/Taxodiaceaepollenites (7), Monoporopollenites (12) and Chenopodipollis (20) provide support for node III to group Lagunillas (Lag) with node IV. Clade IV relates the Cuayuca “B” section with node V because they share Momipites microcoryphaeus (25), Myrtaceidites (28) and Ulmipollenites (43). Node V associates the Cuayuca B section with node VI based on the presence of Momipites tenuipolus (26), Platanoidites (29), Rugulitriporites (33) and Armeria type (38). Izúcar de Matamoros “F” section and node VII share Eucommia (21) and Striatricolporites (35). Finally, the derived state -node VII- shows cladistic relationship between the Cuayuca Principal section and Cuayuca Section A (CyA) due to the presence of Aglaoreidia (10) and Aeschynomene (37).
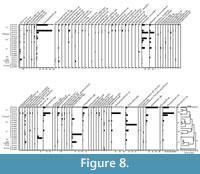 PAE analysis applied to Cuayuca sections expresses historical links, related by space-time (Figure 7). According to field observations, sections grouped at the base of the cladogram correspond to the base of the Cuayuca Mcy Member (Figure 7: group A); in contrast, the most derived sections shown in the cladogram are located within the upper layers (Figure 7: group B) of the Cuayuca Formation. On the other hand, basal sections were less diverse, while more derived sections are the most diverse and yield a great abundance of palynomorphs (Figure 8).
PAE analysis applied to Cuayuca sections expresses historical links, related by space-time (Figure 7). According to field observations, sections grouped at the base of the cladogram correspond to the base of the Cuayuca Mcy Member (Figure 7: group A); in contrast, the most derived sections shown in the cladogram are located within the upper layers (Figure 7: group B) of the Cuayuca Formation. On the other hand, basal sections were less diverse, while more derived sections are the most diverse and yield a great abundance of palynomorphs (Figure 8).
Finally, index taxa such as Aglaoreidia (10), Eucommia (21), Momipites tenuipolus (26), and Armeria (38) are found among the taxa defining the most derived hierarchic levels. On the other hand, two index taxa are located at different hierarchic levels: Ranunculacidites operculatus (30) and Thomsonipollissabinetownensis (36).
CONISS Analysis
Finally, considering that the hierarchic levels of sections in the PAE analysis were concordant with their lithological position in the field, pollen assemblages were plotted and a CONISS analysis was applied to recognize paleopalynological changes throughout Cuayuca Formation Mcy Member. The graph (Figure 8) shows four paleopalynological groups. Samples taken from the base of the Cuayuca Formation, group A, are characterized by the dominance of Alnusvera, Rhoipites and Tubulifloridites; group B shows dominance of the Momipites group and Tubulifloridites; Momipites group and Liliacidites characterize group C; finally, group D shows abundance of temperate elements such as Momipites group, Ulmipollenites and Pinuspollenites, as well as semiarid taxa, for instance, Brosipollis, Chenopodipollis, Monoporopollenites, Liliacidites, and Ephedripites.
In summary, within the upper sections of the Cuayuca Formation, tropical deciduous forest, and arid tropical scrub, as well as cloud forest and Pinus forest are well represented, contrasting with the lower sections, where Rhoipites and Tubulifloridites elements from tropical deciduous forest are dominant.
Index Taxa
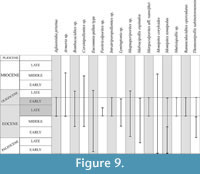 Pollen grain morphology of the best preserved angiosperm specimens is described. Several pollen types found in the Cuayuca Formation are useful age-diagnostic taxa. Angiosperms of chronostratigraphic importance in south central Mexico, such as Aglaoreidia pristina, Armeria, Bombacacidites, Corsinipollenites, Eucommia, Favitricolporites, Intratriporopollenites, Lymingtonia, Magnaperiporites, Malvacipollis spinulosa, Margocolporites aff. vanwijhei, Momipites coryloides, Momipites tenuipolus, Mutisiapollis, Ranunculaciditesoperculatus, and Thomsonipollis sabinetownensis all suggest a late Eocene - early Oligocene age for the Cuayuca Formation (Figure 9). The index taxa have been useful for correlating outcrops from Izúcar de Matamoros sections with the Cuayuca Formation stratotypes (Figure 10), thus allowing for the conclusion that the Izúcar de Matamoros outcrops are part of the Evaporitic Member of the Cuayuca Formation (Ramírez-Arriaga et al., 2008).
Pollen grain morphology of the best preserved angiosperm specimens is described. Several pollen types found in the Cuayuca Formation are useful age-diagnostic taxa. Angiosperms of chronostratigraphic importance in south central Mexico, such as Aglaoreidia pristina, Armeria, Bombacacidites, Corsinipollenites, Eucommia, Favitricolporites, Intratriporopollenites, Lymingtonia, Magnaperiporites, Malvacipollis spinulosa, Margocolporites aff. vanwijhei, Momipites coryloides, Momipites tenuipolus, Mutisiapollis, Ranunculaciditesoperculatus, and Thomsonipollis sabinetownensis all suggest a late Eocene - early Oligocene age for the Cuayuca Formation (Figure 9). The index taxa have been useful for correlating outcrops from Izúcar de Matamoros sections with the Cuayuca Formation stratotypes (Figure 10), thus allowing for the conclusion that the Izúcar de Matamoros outcrops are part of the Evaporitic Member of the Cuayuca Formation (Ramírez-Arriaga et al., 2008).
CONCLUSION
 This contribution is the first to include morphological descriptions of 48 angiosperm pollen grain taxa from the Cuayuca Formation, south-central Mexico, identified to genus level and in some cases to species level. It complements a previous systematic study describing pteridophyte spores and gymnosperm pollen grains (Ramírez-Arriaga et al., 2005).
This contribution is the first to include morphological descriptions of 48 angiosperm pollen grain taxa from the Cuayuca Formation, south-central Mexico, identified to genus level and in some cases to species level. It complements a previous systematic study describing pteridophyte spores and gymnosperm pollen grains (Ramírez-Arriaga et al., 2005).
In the palynoflora recovered from the Cuayuca Formation we have recognized representatives of semiarid vegetation such as tropical deciduous forest, chaparral and arid tropical scrub; nevertheless, temperate taxa from Pinus forest and cloud forest were also present.
A total of 16 age-diagnostic index taxa have been useful in assigning a late Eocene - early Oligocene age for the Cuayuca Formation: Aglaoreidia pristina, Armeria, Bombacacidites, Corsinipollenites, Eucommia, Favitricolporites, Intratriporopollenites, Lymingtonia, Magnaperiporites, Malvacipollis spinulosa, Margocolporites aff. vanwijhei, Momipites coryloides, Momipites tenuipolus, Mutisiapollis, Ranunculacidites operculatus, and Thomsonipollis sabinetownensis.
PAE analysis showed space-time relationships between sections. Upper sections from the Cuayuca Formation were characterized by dominance of semiarid taxa such as Chenopodipollis, Monoporopollenites, and Ephedripites, suggesting the presence of arid tropical scrub. Additionally, tropical deciduous forest, cloud forest, and Pinus forest are well represented. In the lower sections, Rhoipites and Tubulifloridites elements from tropical deciduous forest were dominant.
Finally, the analysis of angiosperm pollen from the Cuayuca Formation is a valuable tool for understanding the evolution of Mexican plant diversity. Since many of the plant families represented in the Cuayuca Formation palynoflora are part of the current vegetation (Appendix 2) growing in the Balsas River Basin (Fernández-Nava et al., 1998), most of the angiosperm families identified from the Cuayuca Formation may have been present within the Cuayuca region since the Paleogene.
ACKNOWLEDGEMENTS
This research was supported by the DGAPA-PAPIIT-IN105411 and DGAPA- PAPIIT-IN114914-3 projects, Universidad Nacional Autónoma de Mexico (UNAM). The authors are grateful to Magdalena Alcayde for the first draft revision and S.H. Ponce Wainer for revising and proofreading the final English version of the manuscript. We thank doctoral candidate I.G. Escobedo for her invaluable support in the statistical analysis, and the corrections of two PE reviewers that greatly enriched the present manuscript.
REFERENCES
AAPG III. 2009. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Botanical Journal of the Linnean Society, 161:105-121.
Anderson, R.Y. 1960. Cretaceous-Tertiary palynology, eastern side of the San Juan basin, New Mexico. New Mexico Bur. Mines and Mineral Resources Mem., 6:1-58.
Aublet, J.B.C.F. 1775. Histoire des plantes de la Guyane Française. Didot, Londres et Paris. Tome 2:622-976.
Baksi, S.K. and Deb, U. 1976. On Mulleripollis gen. nov. a pollen tetrad from the upper Cretaceous of the Bengal Basin, West Bengal, India. Review of Palaeobotany and Palynology, 22:73-77.
Barreda, V.D. 1997a. Palynomorph assemblage of the Chenque Formation, Late Oligocene?-Miocene from Golfo San Jorge Basin, Patagonia, Argentina. Part 1: Terrestrial algae, trilete and monolete spores. Ameghiniana, 34:69-80.
Barreda, B. 1997b. Palynomorph assemblage of the Chenque Formation, Late Oligocene?-Miocene from Golfo San Jorge basin, Patagonia, Argentina. Part 3. Polycolpate and tricolporate pollen. Ameghiniana, 34:131-144.
Barreda, B. 1997c. Palynomorph assemblage of the Chenque Formation, Late Oligocene?-Miocene from Golfo San Jorge basin, Patagonia, Argentina. Part 4. Polycolporate and porate pollen. Ameghiniana, 34:145-154.
Barreda, V. and Palamarczuk, S. 2000a. Palinoestratigrafía de depósitos del Oligoceno tardío - Mioceno en el área sur del Golfo San Jorge, Provincia de Santa Cruz, Argentina. Ameghiniana, 37:103-117.
Barreda, V.D. and Palamarczuk, S. 2000b. Estudio palinoestratigráfico del Oligoceno tardío - Mioceno en secciones de la costa Patagónica y plataforma continental Argentina, p. 103-138. In Aceñolaza, F.G. and Herbst, R. (eds.). El Neógeno de Argentina. INSUGEO, Serie Correlación Geológica , 14.
Barreda, V.D., Gutiérrez, P.R., and Limarino, C.O. 1998. Edad y paleoambiente de la "Serie del Yeso", Valle del Cura, Provincia de San Juan: Evidencias palinológicas. Ameghiniana, 35:321-335.
Bartlett, A.S. and Barghoorn, E.S. 1973. Phytogeographic history of the Isthmus of Panama during the past 12,000 years (a history of vegetation, climate and sea-level change), p. 203-209. In Graham A. (ed.) Vegetation and vegetational history of northern Latin America. Elsevier, Amsterdam.
Bebout, J.W. 1980. Observed stratigraphic distribution of spores, pollen and incertaesedis palynomorphs in the Tertiary section of the COST No. B-2 well, Baltimore Canyon, Atlantic outer Continental shelf. Palynology, 4:181-196.
Bentham, G. and Hooker, J.D. 1862. Genera plantarum. Hookerian herbarium. Kew, voluminis primi Paris II.
Berchtold, F. and Presl, J.S. 1820. O Přirozenosti Rostlin. Prir. Rostlin, Praha, Czech Republic.
Biaggi, R.E. 1978. Palynology and paleoecology of some Oligo-Miocene sediments from Chiapas, Mexico. MSc Thesis, Walla Walla College, WA, USA.
Bromhead, E.F. 1838. Edinburgh New Philosophical Journal. Quarterly Journal of Science, 25:126.
Brown, R. 1814. General Remarks, Geographical and Systematical, on the Botany of Terra Australis, p. 533-613. In Flinders, M. (ed.), A Voyage to Terra Australis(Vol 2). G. and W. Nicol, London.
Cavagnetto, C., and Anadón, A. 1996. Preliminary palynological data on floristic and climatic changes during the middle Eocene-early Oligocene of the eastern Ebro Basin, northeast Spain. Review of Palaeobotany and Palynology, 92:281-305.
Cavagnetto, C. and Guinet, P. 1994. Pollen fossile de Leguminosae - Mimosoideae dans l’Oligocene inférieur du basin de l’Ebre (Espagne) - implications paléoclimatiques et paléogeographiques. Review of Palaeobotany and Palynology, 81:327-335.
Christopher, R.A., Powell, D.C., Reinhardt, J., and Markewich, H.W. 1980. The stratigraphic and structural significance of Paleocene pollen from Warm Springs, Georgia. Palynology, 4:105-124.
Collinson, M.E., Fowler, K., and Boulter, M.C. 1981. Floristic changes indicate a cooling climate in the Eocene of southern England. Nature, 291:315-317.
Cookson, I.C. 1947. Plant microfossils from the lignites of Kerguelen Archipelago, B.A.N.Z. Antarctic Research Expedition, 2:129-142.
Cookson, I.C. and Pike, K. 1954. Some dicotyledonous pollen types from Cainozoic deposits in the Australian region. Australian Journal of Botany, 2:197-219.
Couper, R.A. 1953. Upper Mesozoic and Cainozoic spores and pollen grains from New Zealand. New Zealand Geological Survey, Palaeontological Bulletin, 22:1-77.
Couper, R.A. 1960. New Zealand Mesozoic and Cainozoic plant microfossils. New Zealand Geological Survey, Palaeontological Bulletin, 32:1-87.
Cronquist, A. 1981. An integrated system of classification of flowering plants. Columbia University Press, New York.
Cronquist, A., Takhtajan, A., and Zimmermann, W. 1966. On the higher taxa of Embryobionta. Taxon, 15:129-134.
Cross, A.T. and Martínez-Hernández, E. 1980. Compositae pollen in early Tertiary rocks, Baja California, Mexico. 5th. International, Palynological Conference, (Cambridge), Abstracts, p. 97.
Doyle, J.A. and Robbins, E.J.I. 1975. Angiosperm pollen zonation of the continental Cretaceous of the Atlantic coastal plain and its application to deep wells in the Salisbury Embayment. Proceedings of the Eight Annual Meeting (Houston, Texas). Palynology, 1:43-78.
Ducke, A. 1922. Plantes nouvelles ou peu connues de la Région Amazonienne (IIe partie). Archivos do Jardim Botanico do Rio de Janeiro, 3:3-281.
Eisawi, A. and Schrank, E. 2008. Upper Cretaceous to Neogene palynology of the Melut Basin, southeast Sudan. Palynology, 32:101-129.
El Ghazali, G.E.B. 1990. An illustrated key to endoaperture morphology. Review of Palaeobotany and Palynology, 63:227-231.
Elsik, W.C. 1968. Palynology of a Paleocene Rockdale lignite, Milan County, Texas. II. Morphology and taxonomy. Pollen et Spores, 10:16-44.
Elsik, W.C. 1974. Characteristic Eocene palynomorphs in the Gulf Coast, U.S.A. Palaeontographica AbteilungB, 149:90-111.
Elsik, W.C.H. and Dilcher, D.L. 1974. Palynology and age of clays exposed in the Lawrence Clay Pit, Henry County, Tennessee . Palaeontographica Abteilung B, 146:65-87.
Engler, A. 1892. Syllabus der Vorlesungen über specielle und medicinisch-pharmaceutische Botanik: eine Übersicht über das Pflanzensystem mit Berücksichtigung der Medicinal- und Nutzpflanzen. Gebrüder Borntraeger Verlag, Berlin.
Engler, H.G.A. 1907. Syllabus der Pflanzenfamilien (ed. 5) 139.
Erdtman, G. 1960. On three new genera from the L. Headon Beds . Botaniska Notiser, 113:46-48.
Erdtman, G. 1972. AnIntroduction to Palynology I. Pollen morphology and plant taxonomy: Angiosperms. Almqvist and Viksell, Stockholm.
Fernández, D.A., Panti, C., Palazzesi, L., and Barreda, V.B. 2012. La presencia de una familia neotropical (Malpighiaceae) en el extremo más austral de Sudamérica durante el Eoceno. Revista Brasileira de Paleontologia, 15:386-391.
Fernández-Nava, R., Rodríguez, J.C., Arreguín, S.M.L., and Rodríguez, J.A. 1998. Listado florístico de la cuenca del Río Balsas, México. Polibotánica, 9:1-151.
Fowler, K. 1971. A new species of Aglaoreidia Erdtm. from the Eocene of southern England. Pollen et Spores, 13:135-147.
Frederiksen, N.O. 1979. Paleogene sporomorph biostratigraphy, northeastern Virginia. Palynology, 3:129-167.
Frederiksen, N.O. 1980a. Sporomorphs from the Jackson Group (upper Eocene) and adjacent strata of Mississippi and western Alabama. U.S. Geological Survey Professional Paper, 1084:1-75.
Frederiksen, N.O. 1980b. Paleogene sporomorphs from South Carolina and quantitative correlations with the Gulf Coast. Palynology, 4:125-179.
Frederiksen, N.O. 1983. Middle Eocene palynomorphs from San Diego, California. Part II. Angiosperm pollen and miscellanea. American Association of Stratigraphic Palynologists Contribution Series, 12:32-155.
Frederiksen, N.O. 1984. Stratigraphic, paleoclimatic and paleobiogeographic significance of Tertiary sporomorphs from Massachusetts . U.S. Geological Survey Professional Paper, 1308:1-25.
Frederiksen, N.O. 1988. Sporomorph biostratigraphy, floral changes, and paleoclimatology, Eocene and early Oligocene of the eastern Gulf Coast. U.S. Geological Survey Professional Paper, 1448:1-101.
Frederiksen, N.O. 1989. Eocene sporomorph biostratigraphy of southern California. Palaeontographica, 211:135-179.
Frederiksen, N.O. 1998. Upper Paleocene and lowermost Eocene angiosperm pollen biostratigraphy of the eastern Gulf Coast and Virginia. Micropaleontology, 44:45-68.
Frederiksen, N.O., Carr, D.R., Lowe, G.D., and Wosika, E.P. 1983. Middle Eocene palynomorphs from San Diego, California. American Association of Stratigraphic Palynologists, Contribution Series, 12:1-157.
Fries, C. Jr. 1960. Geología del Estado de Morelos y de partes adyacentes de México y Guerrero, región central meridional de México. Universidad Nacional Autónoma de México, Instituto de GeologíaBoletín, 60:1-236.
Fries, C. Jr. 1966. Hoja Cuernavaca 14Q-h(8). Resumen de la geología de la Hoja Cuernavaca, Estado de Morelos. Universidad Nacional Autónoma de México, Instituto de Geología, Carta Geológica de México. Serie 1: 100, 000, mapa con texto.
Gaponoff, S.L. 1984. Palynology of the Silverado Formation (late Paleocene), Riverside and Orange counties, California. Palynology, 8:71-106.
Germeraad, J.H., Hopping, C.A., and Muller, J. 1968. Palynology of Tertiary sediments from tropical areas. Review of Palaeobotany and Palynology, 6:189-348.
Goloboff, P.A. 1993. NONA version 2.0 Program and documentation. Computer program distributed by J.M. Carpenter, Dept. of Entomology. American Museum of Natural History, New York.
González-Guzmán, A.E. 1967. A palynological study on the upper Los Cuervos and Mirador formations ( lower and middle Eocene; Tribú Area, Colombia ). E.J. Brill, Leiden.
Graham, A. 1975. Late Cenozoic evolution of tropical lowland vegetation in Veracruz, Mexico. Evolution, 29:723-735.
Graham, A. 1976. Studies in Neotropical Paleobotany.II. The Miocene communities of Veracruz, Mexico. Annals of the Missouri Botanical Garden, 63:782-842.
Graham, A. 1985. Studies in Neotropical Paleobotany. IV. The Eocene communities of Panama. Annals of the Missouri Botanical Garden, 72:504-534.
Graham, A. 1987. Miocene communities and paleoenvironments of southern Costa Rica. American Journal of Botany, 74:1501-1518.
Graham, A. 1988. Studies in Neotropical Paleobotany. V. The lower Miocene communities of Panama - La Culebra Formation. Annals of the Missouri Botanical Garden, 75:1440-1466.
Graham, A. 1989. Studies in Neotropical Paleobotany. VII. The lower Miocene communities of Panama - the La Boca Formation. Annals of the Missouri Botanical Garden, 76:50-66.
Graham, A. 1990a. New angiosperm records from the Caribbean Tertiary. American Journal of Botany, 77:897-910.
Graham, A. 1990b. Late Tertiary microfossil flora from the Republic of Haiti. American Journal of Botany, 77:911-926.
Graham, A. 1991. Studies in Neotropical Paleobotany. VII. The Pliocene communities of Panama - Introduction and ferns, gymnosperms, angiosperms (monocots). Annals of the Missouri Botanical Garden, 78:190-200.
Graham, A. 1999. Studies in Neotropical Paleobotany. XIII. An Oligo-Miocene palynoflora from Simojovel (Chiapas, Mexico). American Journal of Botany, 86:17-31.
Graham, A. 2000. Studies in Neotropical Palaeobotany. IV. A palynoflora from the middle Eocene Saramaguacán Formation of Cuba. American Journal of Botany, 87:1526-1539.
Graham, A. 2010. Late Cretaceous and Cenozoic History of Latin American Vegetation and Terrestrial Environments. Missouri Botanical Garden Press, USA.
Graham, A., Cozadd, D., Areces-Mallea, A., and Frederiksen, N.O. 2000. Studies in Neotropical Paleobotany XIV. A palynoflora from the middle Eocene Saramaguacán Formation of Cuba. American Journal of Botany, 87:1526-1539.
Graham, A. and Jarzen, D.M. 1969. Studies in Neotropical Paleobotany. I. The Oligocene communities of Puerto Rico. Annals of the Missouri Botanical Garden, 56:308-357.
Grimm, E.C. 1987. CONISS: A Fortran 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Computers & Geosciences, 13:13-35.
Haeckel, E. 1866. Generelle Morphologie der Organismen: allgemeine Grundzüge der organischen Formen-Wissenschaft, mechanisch begründet durch die von C. Darwin reformirte Decendenz-Theorie. Berlin.
Harris, W.K. 1965. Basal Tertiary microfloras from the Princetown area, Victoria, Australia. Palaeontographica AbteilungB, 115:75-106.
Jansonius, J. and Hills, L.V. 1976. Genera file of fossil spores and pollen. Special Publication Department of Geology. University of Calgary, Canada (card numbers: 1562, 2338, 2441, 3110, 2086, and 3121).
Jaramillo, C.A. and Dilcher, D.L. 2001. Middle Paleogene palynology of central Colombia, South America: a study of pollen and spores from tropical latitudes. Paleontographica AbteilungB, 258:87-213.
Jardine, P.E. and Harrington, G.J. 2008. The Red Hills Mine palynoflora: A diverse swamp assemblage from the late Paleocene of Mississippi, U.S.A. Palynology, 32:183-204.
Jarzen, D.M. and Klug, C. 2010. A preliminary investigation of the lower to middle Eocene palynoflora from Pine Island, Florida, U.S.A. Palynology, 34:164-179.
Jarzen, D.M., Corbett, S.L., and Manchester, S.R. 2010. Palynology and paleoecology of the middle Miocene Alum Bluff flora, Liberty County, Florida, U.S.A. Palynology, 34:261-286.
Jaramillo, C.A. and Dilcher, D.L. 2001. Middle Paleogene palynology of central Colombia, South America: a study of pollen and spores from tropical latitudes. PalaeontographicaB, 258:87-213.
Johnson, D.E. 2000. Métodos Multivariados Aplicados al Análisis de Datos. International Thomson Editores, México.
Jussieu de, A.L. 1789. Genera Plantarum secundum ordines naturales disposita. Viduam Herissant et Theophilum Barrois, 1-498.
Kayseri, M.S. and Akgün, F. 2008. Palynostratigraphic, palaeovegetational and palaeoclimatic investigations on the Miocene deposits in central Anatolia (Çorum Region and Sivas Basin). Turkish Journal of Earth Sciences, 17:361-403.
Krutzsch, W. 1960. Über Thomsonipollis magnificus (Th. and Pf. 1953) n. fgen. n. comb. und Bemerkungen zur regionalen Verbreitung einiger Pollengruppen im älteren Paläogen. Freiburger Forschungshefte, C86:54-65.
Krutsch, W. 1963. Atlas der mittel-und jungtertiären dispersen Sporen-und Pollen-sowie der Mikroplanktonformen des nördlichen Mitteleuropas. Lieferung II. VEB Deutscher Verlag der Wissenschaften, Berlin.
Krutzsch, W. 1966. Die sporenstratigraphische Gliderung des Älteren Tertiär in nördlichen Mitteleuropa (Paläozän - Mitteloligozän). Metodische grundlagen und gegenwärtiger stand der untersuchungen. Abhandlungen des Zentralen Geologischen Institut, 8:112-149.
Krutzsch, W. 1968. Brosipollis und Labrapollis, zwei neue Pollengenera aus dem Tertiär Mitteleuropas: Review of Palaeobotany and Palynology, 6:61-70.
Langenheim, J.H., Hackner, B.L., and Bartlett, A. 1967. Mangrove pollen at the depositional site of Oligo-Miocene amber from Chiapas, Mexico. Botanical Museum Leaflets, Harvard University, 21:289-324.
Leidelmeyer, P. 1966. The Paleocene and lower Eocene pollen flora of Guyana. Leidse Geologische Mededelingen, 38:49-70.
Leopold, E.B. 1974. Pollen and spores of the Kisinger Lakes fossil leaf locality. In Mac Ginitie, H.D. An early Middle Eocene flora fron the Yellowstone-Absaroka Volcanic Province, northwestern Wind River Basin, Wyoming. University of California, Publications in Geological Science.
Leopold, E.B. and Scott, C.P. 2001. Florissant leaf and pollen floras of Colorado compared: climatic implications. Denver Museum of Nature and Science, 4:17-69.
Lima, M.R., Melo, M.S., and Coimbra, A.M. 1991. Palinologia de sedimentos da Bacia de São Paulo, Terciario do Estado de São Paulo, Brasil. Rev. IG, São Paulo, 12:7-20.
Lindley, J. 1833. Nixus Plantarum. Bradbury and Evans Printers, London.
Lindley, J. 1836. A natural system of botany. Bradbury and Evans Printers, London.
Lindley, J. 1846. The Vegetable Kingdom. Bradbury and Evans Printers, London.
Linnaeus, C. 1753. Species Plantarum 1. Laurentius Salvius. Stockholm.
Lorente, A.M.A. 1986. Palynology and palynofacies of the upper Tertiary in Venezuela. Dissertationes Botanicae, Band 99. J.Cramer, Venezuela.
Love, J.D., Leopold, E.B., and Love, D.W. 1978. Eocene rocks, fossils, and geologic history: Teton Range, northwestern Wyoming. Washington, DC. U.S. Geological Survey Professional Paper, 932-B:1-40.
MacGinitie, H.D. 1962. The Kilgore flora. A late Miocene flora fron northern Nebraska. University of California Publications in Geological Sciences, 35:67-158.
Macphail, M.K. 1999. Palynostratigraphy of the Murray Basin, inland southeastern Australia. Palynology, 23:197-240.
Macphail, M.K. and Hill, R.S. 1994. K-Ar dated palynofloras in Tasmania 1: Early Oligocene, Proteacidites tuberculatus Zone sediments, Wilmot Dam, northwestern Tasmania. Papers and Proceedings of the Royal Society of Tasmania, 128:1-15.
Mai, D.H. 1961. Über eine fossile Tiliaceen - Blute und tilioiden pollen aus dem deutschen Tertiär. Geologie, 32:54-93.
Martin, H.A. 2002. History of the Malpighiaceae in Australia and its biogeographic implications: evidence from pollen. Australian Journal of Botany, 50:171-182.
Martínez-Hernández, E. and Ramírez-Arriaga, E. 1996. Paleocorología de angiospermas de la flora Mexicana durante el Mesozoico y Terciario. Algunas evidencias palinológicas. Boletín de la Sociedad Botánica de México, 58:87-97.
Martínez-Hernández, E. and Ramírez-Arriaga, E. 1999. Palinoestratigrafía de la region de Tepexi de Rodríguez, Puebla, México - implicaciones cronoestratigráficas. Revista Mexicana de Ciencias Geológicas, 16:187-207.
Martínez-Hernández, E. and Ramírez-Arriaga, E. 2006. Tertiary palynofloristic correlations between Mexican basins with emphasis in dating the Balsas Group, p. 19-45. In Vega, F.V., Nyborg, T.G., Perrilliat, M.C., Montellano, M., Cevallos, S., and Quiroz, S. (eds.), Studies on Mexican Paleontology. Springer, the Netherlands.
Martínez-Hernández, E., Hernández-Campos, H., and Sánchez-López, M. 1980. Palinología del Eoceno en el noreste de México. UNAM, Instituto de Geología. Revista, 4:155-166.
Mierbel, B.C.F. 1815. Eléments de Physiologie Végétale et de Botanique. Avec supplément: De la composition chimique des végétaux par M. Chevreul. Chez Magimel, Librairie, Rue de Thionville, Nº 9. Paris.
Morrone, J.J. and Crisci, J.V. 1995. Historical Biogeography: Introduction to methods. Annu. Rev. Ecol. Syst., 26:373-401.
Muller, J. 1968. Palynology of the Pedawan and Plateau Sandstone formations (Cretaceous - Eocene) in Sarawak, Malaysia. Micropaleontology, 14:1-37.
Nakoman, E. 1965. Description d’un nouveau genre de forme: Corsinipollenites. Annales Societé Géologique du Nord, 85:155-158.
Newman, K.R. 1965. Upper Cretaceous - Paleocene guide palynomorphs from northwestern Colorado. Series in Earth Sciences No. 2, University of Colorado Press, Boulder, Colorado.
Nichols, D.J. 2010. Reevaluation of the holotypes of the Wodehouse pollen species from the Green River Formation (Eocene, Colorado and Utah). AASP Contribution Series, 44:1-97.
Nichols, D.J., Ames, H.T., and Traverse, A. 1973. On Arecipites Wodehouse, Monocolpopollenites Thomson and Pflug, and the species “ Monocolpopollenitestranquillus ”. Taxon, 22:241-256.
Nixon, K.C. 2002. WinClada ver. 1.00.08 Published by the author, Ithaca, N.Y.
Oliver, D. 1890. Eucommia ulmoides Oliv. Hooker’s Icones Plantarum, 20.
Ottone, E.G., Barreda, V.D., and Pérez, D.J. 1998. Basin evolution as reflected by Miocene palynomorphs from the Chinches Formation, frontal cordillera (32ºS), San Juan Province, Argentina. Revista Española de Micropaleontología, 30:35-47.
Palacios, Ch.R. and Rzedowski, J. 1993. Estudio palinológico de las floras fósiles del Mioceno inferior y principios del Mioceno medio de la región de Pichucalco, Chiapas. Acta Botánica Mexicana, 24:1-96.
Palisot de Beauvois, A.M.F.J. 1806. Flore d’Oware et de Benin, en Afrique. Imprimerie de Fain Jeune et Compagnie, Paris, 1:49-60.
Papanicolaou, K. and Kokkini, S. 1982. Armeria (Plumbaginaceae) in Greek mountains. Willdenowia, 12:213-219.
Perleb, K.J. 1818. Versuch über die Arzneikräfte der Pflanzen: verglichen mit den aüsseren. Vers. Arzneikr. Pfl., 1-450.
Pflug, H.D. 1953. Zur Entstehung und Entwicklung des angiospermiden Pollens in der Erdgeschichte. Palaeontographica, Abt. B., 95:60-171.
Pocknall, D.T. and Jarzen, D.M. 2009. Pollen with viscin threads from the Late Cretaceous and Paleocene, Merida Andes, western Venezuela. Palynology, 33:55-61.
Pocknall, D.T. and Mildenhall, D.C. 1984. Late Oligocene - early Miocene spores and pollen from Southland, New Zealand. New ZealandGeological Survey Paleontological Bulletin, 51:1- 66.
Potonié, R. 1931. Zur Mikroskopie der Braunkohlen. Tertiäre Blütenstaubformen. Braunkohle, 20:554-556.
Potonié, R. 1934. Zur mikrobotanik des Eocänen Humodils des Geiseltals. Preuss. Geol. Landesanstalt.Arbeiten Inst. Paläobot. Petrog. Brennsteine: 25-125.
Potonié, R. 1958. Synopsis der Gattungen der sporae dispersae. Teil 2. Beihefte zum Geologischen Jahrbuch, 31:1-114.
Potonié, R. 1960. Synopsis der Gattungen der sporae dispersae. III. Teil: Nachträge Sporites, Forsetzung Pollenites. Mit Generalregister zu Teil I - III. Beihefte zum Geologisches Jahrbuch, 39:1-189.
Punt, W., Hoen, P.P., Blackmore, S., Nilsson, S., and Le Thomas, A. 2007. Glossary of pollen and spore terminology. Review of Palaeobotany and Palynology, 143:1-81.
Ramanujam, C.G.K. 1966. Palynology of the Miocene lignite from South Arcot District, Madras, India. Pollen et Spores, 8:149-203.
Quattrocchio, M., Ruiz, L., and Volkheimer, W. 2000. Palynological zonation of the Paleogene of the Colorado and Salta Group Basins, Argentina. Revista Española de Micropaleontología, 32:61-78.
Ramírez-Arriaga, E., Martínez-Hernández, E., and Prámparo, M.B. 2005. Palynomorph assemblages of the Cuayuca Formation (Paleogene), Puebla, southern Mexico. Part 1: trilete spores, monolete spores and gymnosperm pollen. Ameghiniana, 42:567-578.
Ramírez-Arriaga, E., Martínez-Hernández, E., and Prámparo, M.B. 2006. Palynology of the Paleogene Cuayuca Formation (stratotype sections), southern Mexico: chronostratigraphical and palaeoecological implications. Review of Palaeobotany and Palynology, 141:259-275.
Ramírez-Arriaga, E., Martínez-Hernández, E., Flores-Olvera, H., Ochoterena, H., and Prámparo, M. 2008. Correlation of the late Eocene - early Oligocene Izúcar de Matamoros evaporites (Cuayuca Formation) in Mexico based on parsimony analysis of endemicity. Palynology, 32:231-252.
Ramírez-Arriaga, E., Prámparo, M.B., Nieto-Samaniego, A.F., Martínez-Hernández, E., Valiente-Banuet, A., Macías-Romo, C., and Dávalos-Álvarez, O.G. 2014. Palynological evidence for Middle Miocene vegetation in the Tehuacán Formation of Puebla, Mexico. Palynology, 38:1-27.
Regali, M.S.P., Uesugui, N., and Santos, A.S. 1974. Palinologia dos sedimentos Meso-Cenozóicos do Brasil. Parte 1. Boletin Tecnico Petrobrás, 17:177-191.
Roche, E. and Schuler, M. 1976. Analyse palynologique (pollen et spores) de divers gisements du Tongrien de Belgique. Interprétation paléoecologique et stratigraphique. Administration des Mines-Service Géologique de Belgique Professional Paper, 11:1-57.
Rosen, D.E. 1988. From fossils to earth history: applied historical biogeography, p. 437-481. In Myers, A.A. and Giller, P.S. (eds.), Analytical biogeography an integrated approach to the study of animal and plant distributions. Chapman, London and New York.
Rull, V. 2001. Quantitative palynological record from the early Miocene of western Venazuela, with emphasis on mangroves. Palynology, 25:109-126.
Rull, V. 2003. Contributions of quantitative ecological methods to the interpretation of stratigraphically homogeneous pre-Quaternary sediments: a palynological example from the Oligocene of Venezuela. Palynology, 27:75-98.
Rzedowsky, J. 2006. Vegetación de México. 1a. Edicióndigital.Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, México.
Sah, S.C.D. 1967. Palynology of an upper Neogene profile from Rusizi Valley (Burundi). Musée Royal de l’Afrique Centrale. Annales, Ser. 8º. Sciences Géologiques, 57:1-173.
Sah, S.C.D. and Dutta, S.K. 1966. Palyno-stratigraphy of the sedimentary formations of Assam: 1. Stratigraphical position of the Cherra Formation. Palaeobotanist, 15:72-86.
Salard-Cheboldaeff, M. 1979. Palynologie Maestrichtienne et Tertiaire du Cameroun. Etude qualitative et répartition verticale des principales espèces. Review of Palaeobotany and Palynology, 28:365-388.
Salard-Cheboldaeff, M. 1981. Palynologie Maestrichtienne et Tertiaire du Cameroun. Résultats botaniques. Review of Palaeobotany and Palynology, 32:401-439.
Silva-Romo, G., Mendoza, R.C., and Martiny, B. 2000. Acerca del origen de las cuencas Cenozoicas de la zona cortical extendida del sur de México: Un ejemplo en la región Mixteca. GEOS Boletín Informativo. Época II. Unión Geofísica Mexicana, 20:326-327.
Srivastava, S.K. 1969. Some angiosperm pollen from the Edmonton Formation (Maestrichtian), Alberta, Canada . J. Sen memorial volume: 47-67.
Srivastava, S.K. 1972. Systematic description of some spores from the Edmonton Formation (Maestrichtian), Alberta, Canada. PalaeontographicaAbteilungB, 139:1-46.
Srivastava, S.K. 1984. Palynology of the Monterey Formation (Miocene) phosphatic facies at Lions Head, Santa Maria area, California. Palynology, 8:33-49.
Stover, L.E., Elsik, W.C., and Fairchild, W.W. 1966. New genera and species of early Tertiary palynomorphs from Gulf Coast. Univ. Kansas Paleontological Contributions Paper, 5:1-10.
Takahashi, J. and Jux, U. 1989. Palynology of middle Cenozoic lacustrine deposits from the Jos Plateau, Nigeria. Bulletin of the Faculty of Liberal Arts, Nagasaki University, Natural Science 29:181-367.
Thiele-Pfeiffer, H. 1980. Die Miozäne mikroflora aus dem Braun - kohlentagebau Oder bei Wackersdorf/Oberpfalz. Palaeontographica AbteilungB, 258:95-224.
Thiergart, F. 1940. Die mikropaläontologie als pollenanalyse im Dienst der Braunkohlenforschung., Ferdinand Enke, Stuttgart. Schriften aus dem Gebiet der Brennstoff - Geologie, 13:1- 87.
Thomson, P. and Pflug, H. 1953. Pollen und sporen des mitteleuropäischen Tertiärs. Palaeontographica AbteilungB, 94:1-138.
Tomasini-Ortiz, A.C. and Martínez-Hernández, E. 1984. Palinología del Eoceno - Oligoceno de Simojovel, Chiapas. Universidad Nacional Autónoma de México , Instituto de Geología, Paleontología Mexicana, 50:1-61.
Traverse, A. 1988. Paleopalynology. Unwin Hyman, Boston.
Tschudy, R.H. 1973. Stratigraphic distribution of significant Eocene palynomorphs of the Mississippi Embayment. U.S.Geological Survey Professional Paper, 743B:1-24.
Tschudy, R.H. and van Loenen, S.D. 1970. Illustrations of plant microfossils from the Yazoo Clay (Jackson Group, Upper Eocene) Mississippi. U.S. Geological Survey Professional Paper, 643-E:1-5.
Tsukada, M. and Deevey, E.S. 1967. Pollen analyses from four lakes in the southern Maya area of Guatemala and El Salvador, p. 303-331. In Cushing, E.J. and Wright, Jr. H.E. (eds.), Quaternary paleoecology. New Haven, CT, Yale University Press.
Van der Hammen, T. 1956. Nomenclatura palinológica sistemática. Boletín Geológico del Instituto Geológico Nacional de Colombia, 4:23-62.
van der Hammen, T. and Wymstra, T.A. 1964. A palynological study on the Tertiary and upper Cretaceous of British Guiana. Leidse Geologische Mededelingen, 30:184-241.
Van Pelt, R.S., Engelhardt, D.W., Christopher, R.A., and Lucas-Clark, J. 2000. Establishing a hydrostratigraphic framework using palynology. An example fron the Savannah River Site, South Carolina. Chapter 18. In Martin, R.E. (ed.), Environmental Micropaleontology v. 15 Topics in Geobiology. Kluwer Academic/Plenum Publishers, New York.
Velázquez, S.A.L. 2008. Lluvia de polen en un gradiente altitudinal en la ladera NW del Volcán La Malinche, Tlaxcala México. MSc Thesis, Instituto Politécnico Nacional, Escuela Nacional de Ciencias Biológicas, Sección de Estudios de Posgrado e Investigación, Mexico.
von Hoffmannsegg, J.C. and Link, J.H.F. 1809. Flore Portugaise ou description de toutes les plantes qui croissent naturellement en Portugal. Fl. Portug., 1:1-458.
Von Martius, C.F.P. 1835. Conspectus Regni Vegetabilis: secundum characteres morphologicos praesertim carpicos in classes ordines et familias digesti, adjectis exemplis nominibusque plantarum usui medico technico et oeconomico inservientium - Uebersicht der Classen, Ordnungen und Familien des Gewächsreiches nach morphologischen Grundsätzen, unter besonderer Rücksicht auf den Fruchtbau, mit Angabe von Beispielen und von den in der Medicin, Technik und Oekonomie besonderswichtigen Pflanzen zunächst als Leitfaden bei seinen akademischen Vorlesungen entworfen. Joh. Leonhard Schrag, Nurnberg.
von Willdenow, C.L. 1809. Enumeratio Plantarum Horti Regii Botanici Berolinensis: continens descriptions omnium vegetabilium in horto dicto cultorum. Berolini, Taberna Libraria Scholare Realis.
Weyland, H. and Pflug, H. 1957. Die Pflanzenreste der pliozänen Braunkohle von Ptolemaïs in Nordgriechenland. Palaeontographica, B, 102:96-108.
Wilkinson G.C. and Boulter, M.C. 1980. Oligocene pollen and spores from the western part of the British Isles. PalaeontographicaAbteilungB, 175:27-83.
Wingate, F.H. 1983. Palynology and age of the Elko Formation (Eocene) near Elko, Nevada. Palynology, 7:93-132.
Wodehouse, R.P. 1933. Tertiary pollen II. The oil shales of the Eocene Green River Formation. BulletinTorrey Botanical Club, 60:479-552.
Wolff, H. 1934. Mikrofossilien des Pliozänen Humodils der Grube Freigericht bei Dettingen am Main und Vergleich mit älteren Schichten des Tertiärs sowie posttertiären Ablagerungen. Arbeitung des Institute Paläobotanische und Petrographische von Brennsteine, 5:55-101.
