

 Alternating stripmining and sequestration in deep-sea sediments: The trace fossil Polykampton–an ecologic and ichnotaxonomic evaluation
Alternating stripmining and sequestration in deep-sea sediments: The trace fossil Polykampton–an ecologic and ichnotaxonomic evaluation
Article number: 22.2.21
https://doi.org/10.26879/930
Copyright Paleontological Society, May 2019
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 6 October 2018. Acceptance: 22 March 2019
{flike id=2491}
ABSTRACT
Polykampton Ooster, 1869, comprises deep-sea trace fossils composed of a horizontal median cylindrical tunnel and lateral spreite lobes that alternate on either side of the tunnel and are slightly inclined to bedding. The lobes in P. alpinum Ooster, 1869, are closely spaced and partly shingled. In P. guberanum isp. nov. they are asymmetric and widely spaced. In both ichnospecies, the lobes are generally below the median tunnel. In P. multiflabellatum isp. nov., the lobes extend from outer bents of the median tunnel that runs below the lobes. Polykampton is a structure produced by a “worm”-like organism occupying a deep endobenthic tier. The tracemaker evidently sequestered organic-rich sediment from the seafloor, stored it in the lobes and processed it later by stripmining for feeding, especially in times of low food availability on the seafloor. Polykampton occurs in upper Albian to Oligocene deep-sea, mainly turbiditic deposits.
Alfred Uchman. Institute of Geological Sciences, Jagiellonian University, Gronostajowa 3a, PL 30-376 Kraków, Poland; alfred.uchman@uj.edu.pl
Andreas Wetzel. Geologisches Institut, Universität Basel, Bernoullistrasse 32, CH-4056 Basel, Switzerland; andreas.wetzel@unibas.ch
Bruno Rattazzi. Museo Paleontologico di Crocefieschi, Via alla Chiesa 12, 16010 Crocefieschi (Genova), Italy; museocrocefieschi@libero.it
Keywords: fossil behavior; sequestration; ichnotaxonomy; new ichnospecies; flysch; Italy; Switzerland
Uchman, Alfred, Wetzel, Andreas, and Rattazzi, Bruno. 2019. Alternating stripmining and sequestration in deep-sea sediments: The trace fossil Polykampton – an ecologic and ichnotaxonomic evaluation. Palaeontologia Electronica 22.2.21A 1-18. https://doi.org/10.26879/930
palaeo-electronica.org/content/2019/2491-trace-fossil-polykampton
Copyright: May 2019 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
http://zoobank.org/A7C616AE-6225-41AA-BD15-FD91CDE0BB7C
INTRODUCTION
The trace fossil Polykampton alpinum Ooster, 1869, the type ichnospecies of Polykampton Ooster, 1869, resembles a feather-stitched garland and was originally classified as a fossil medusae. Previously, it has mostly been shown in drawings (Ooster, 1869; Häntzschel, 1975; Seilacher, 2007). Surprisingly, photographs of the type material have not been published nor has its morphology been analyzed in detail. In the meantime, Caulerpa eseri Unger, 1850, was transferred to Polykampton as P. eseri (Unger, 1850) by Wetzel and Uchman (1997). Later, P. cabellae Uchman and Rattazzi, 2017 was established and a new feeding behavior was proposed for this ichnospecies, which can be extended to other representatives of this ichnogenus. Moreover, two new ichnospecies of Polykampton are here recognized: P. guberanum in Switzerland, and P. multiflabellatum in Upper Cretaceous to Paleocene sediments of Italy, Austria and Spain. Their description and interpretation is an integral part of this study. Therefore, P. alpinum becomes increasingly intriguing with respect to its morphology, the ethology of its tracemaker, and ecology. Fortunately, the type material is preserved in the Natural History Museum of the Citizens’ Community of Bern, Switzerland, and additional material was found within the same lithostratigraphic unit farther to the east. These specimens form a good basis for revisiting Polykampton alpinum. It is the purpose of the present study to provide an overview of the ichnogenus Polykampton and the so far recognized ichnospecies, to show these trace fossils on photographs (including the type specimen), to interpret the behavior of the producers, and to decipher the environmental context of Polykampton.
GEOLOGICAL SETTING
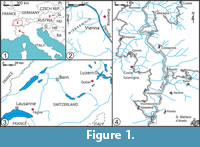 The key specimens studied derive from different parts of the Alpine system in Europe (Figure 1.1), in Austria (Figure 1.2), Switzerland (Fig. 1.3) and Italy (Figure 1.4). The type material of Polykampton alpinum was found at the Fégire locality (Switzerland; coordinates N 46°30'03''; E 06°56'46'') (Figure 1.3) in Paleocene-Eocene turbiditic sandstones (originally considered as Rhaetian), which belong to the Gurnigel Flysch Unit. Polykampton guberanum was found in scree in the Guber quarry (Switzerland; coordinates N 46°55'40''; E 08°14'04'') (Figure 1.3), where turbiditic sandstones of the Schlieren Flysch Unit are exposed.
The key specimens studied derive from different parts of the Alpine system in Europe (Figure 1.1), in Austria (Figure 1.2), Switzerland (Fig. 1.3) and Italy (Figure 1.4). The type material of Polykampton alpinum was found at the Fégire locality (Switzerland; coordinates N 46°30'03''; E 06°56'46'') (Figure 1.3) in Paleocene-Eocene turbiditic sandstones (originally considered as Rhaetian), which belong to the Gurnigel Flysch Unit. Polykampton guberanum was found in scree in the Guber quarry (Switzerland; coordinates N 46°55'40''; E 08°14'04'') (Figure 1.3), where turbiditic sandstones of the Schlieren Flysch Unit are exposed.
The Gurnigel Flysch and the closely related Schlieren Flysch show remarkable lateral continuity in facies. Composite sections more than 1000 m in thickness imply deposition in a basin-plain setting during Maastrichtian to Danian times, while the overlying sand-dominated upper Paleocene to lower Eocene sediments are interpreted to have accumulated on prograding deep-sea fans, in which younger, more proximal depositional lobes are channelized (van Stuijvenberg, 1979; Matter et al., 1980; Crimes et al., 1981; Winkler, 1983). All Polykampton specimens from Switzerland addressed in this study were collected from scree. The massive appearance of the supposedly rather thick sandstone beds containing Polykampton implies that they formed in a channelized depositional setting. All localities were revisited, but unfortunately no new specimens were discovered and, hence, the study is based on already existing material.
Polykampton multiflabellatum was found in the locality Costa Gramigna 2 on the western slope of the Aveto Valley in Liguria, Italy (GPS: N44º35.201'; E009º23.358') (Figure 1.4), where a ~20 m thick section is exposed in an escarpment along a road. The section contains thin-and medium-bedded, fine-grained, calcareous, turbiditic sandstones, which alternate with dark-gray and gray shales. They belong to the Ottone Formation (Flysch di Ottone), which forms part of the succession (Caio/Ottone sequence) of the External Ligurian Unit and is dated to the early to middle Campanian (Marroni et al., 1992, in press; Catanzariti and Perilli, 2006). The Ottone Formation is about 200 m thick. In the lower part, it contains ophiolitic debrites, which pass into calcareous flysch.
The specimen stored in the Naturhistorisches Hofmuseum in Vienna derives from a quarry located to the north of the so-called “Kellergasse” of Langenzersdorf, close to the Bisamberg locality, north of Vienna (Figure 1.2) where the Kahlenberger Schichten (upper Campanian to lowermost Maastrichtian) of the Rhenodanubian Flysch are exposed in an abandoned quarry and in the surrounding area. The approximately 500 m thick Kahlenberger Schichten (Kahlenberg Beds) is composed of thin-bedded, carbonate-rich turbidites with numerous intercalated horizons of variegated shales (Faupl, 1975, 1976; Uchman, 1999).
METHODS
This study is based on visual observations, digital photography and application of some image-processing software to enhance contrast (Photoshop ®). Furthermore, organic carbon and carbonate content of four samples were measured. Organic carbon was calculated as the difference between total carbon measured on a Erba Science NA 1500 Elemental Analyzer and carbonate carbon measured on a Woesthoff Carmograph 6 using calcite standards for calibration and 2N H3 PO4 for sample acidification.
SYSTEMATIC ICHNOLOGY
Ichnogenus POLYKAMPTON Ooster, 1869
Type ichnospecies. Polykampton alpinum Ooster, 1869.
Diagnosis. Horizontal, ribbon structure composed of a median cylindrical tunnel and complex side lobes (modified from Uchman and Rattazzi, 2017).
Remarks. Ooster (1869) did not provide a diagnosis. In the modified diagnosis presented herein, the word “tunnel” replaces “stem” from the diagnosis by Uchman and Rattazzi (2017) as it better describes the morphology and the non-vegetable origin.
Polykampton alpinum Ooster, 1869
Figure 2, Figure 3, Figure 4.1-5
List of records and synonyms
 *v 1869 Polykampton alpinum – Ooster, p. 23, pl. 4, figures 1-4. [Figure 2.5, 2.6; Fégire, Switzerland.]
*v 1869 Polykampton alpinum – Ooster, p. 23, pl. 4, figures 1-4. [Figure 2.5, 2.6; Fégire, Switzerland.]
1895 Polycampton alpinum-Fuchs, 433, pl. 8, figures 1, 2. [nom. null.]
? 1918 Ślad pełzania (?) [trace of creeping (?)] – Zuber, p. 116, figure 76. [copied by Vialov and Zenkevich, 1961, p. 57, figure 3; Vialov, 1993, p. 22, figure 1; Jamna Sandstone, Upper Paleogene, Jamna, Carpathians, Ukraine; sketch in Figure 4.4.]
1959 Polykampton-Seilacher, p. 1070, figure 26.
1965 Polykampton alpinum Ooster – Häntzschel, p. 73.
1973 bioglif – Makarenko, p. 220, fig. on p. 221. [Sketch in Figure 4.3; Roztoki, Sheshorskiy Horizon, Lower Oligocene (see Vialov, 1993), Carpathians, Ukraine.]
1975 Polykampton alpinum – Häntzschel, p. W97, figure 60.5.
? 1980 Sled neizvestnokho organizma [Trace of unknown organism]-Fedchenco and Tatoli, p. 87, figure 1. [Cenozoic, Carpathians, Yaremche, Ukraine.]
1981 Polykampton alpinum Ooster – Crimes et al., p. 975, plate 4, figures 5, 6.
v 1986 Flyschichnium plickai n. ichnosp. – Novák and Peslová, p. 101, plates 19, 20. [sketch in Figure 4.5; Bzová quarry, Eocene, Carpathians, Czech Republic.]
1991 Polykampton-Fu, p. 47.
? 1993 Carpatichnus zuberi Vialov gen. et sp. n.-Vialov, p. 22, figure 1.
non v 1996 Polykampton alpinum Ooster – Rykart, p. 505, figure 4b. [P. guberanum in this study; Gubersandstein in the Schlieren Flysch, Upper Paleocene (Thanetian).]
2007 Polykampton – Seilacher, p. 138, plate 48 [part].
?non 2011 Polykampton – Seilacher and Mrinjek, p. 160, figure 4a.
Diagnosis. Horizontal, straight to winding structure composed of a median tunnel and lateral, inclined, crescentic lobes. The lobes alternate on both sides of the tunnel and can show a shingled and closely spaced arrangement. The lobes usually extend downward below the level of the median tunnel. The upper part of the lobes is commonly traversed by the median tunnel.
Remarks. Ooster (1869) did not provide a diagnosis.
Types. Two slabs, nos. 5017075, 5017076, housed in the Naturhistorisches Museum der Burgergemeinde Bern [Natural History Museum of the Citizens' Community of Bern], but Ooster (1869) did not designate a holotype, so they are syntypes. Therefore, slab 5017075 (Figure 2.1-4, Figure 4.1) is designated as the lectotype and 5017076 (Figure 2.2) as the paralectotype.
 Other specimens. One specimen (Figure 3.1) in the Naturhistorisches Museum, Vienna (specimen NHMW 2018/0158/0001; from “the Rhaetian Zoophycos layers” in Switzerland, identified herein as the Gurnigel Flysch, donated by S. v. Fellenberg, the Head of the Natural History Museum in Bern in 1897).
Other specimens. One specimen (Figure 3.1) in the Naturhistorisches Museum, Vienna (specimen NHMW 2018/0158/0001; from “the Rhaetian Zoophycos layers” in Switzerland, identified herein as the Gurnigel Flysch, donated by S. v. Fellenberg, the Head of the Natural History Museum in Bern in 1897).
Type locality. Banks of a steep-sloped valley along the highly erosive Fégire creek, where it transects the beds of the Gurnigel-Schlieren Flysch Unit for a few hundreds of meters (Swiss coordinates 562'203; 150'068; Geological Atlas of Switzerland, Sheet 1244 Châtel-St-Denis).
Description of type material. Ooster (1869, pl. 4 reproduced in Figure 2.6) illustrated four specimens preserved on one slab in full relief, exposed on a split surface (while the counterpart is missing), and another specimen is present on another sandstone slab. The specimen from his figure 2 is a hypichnion on slab no. 5017075 (Figure 2.1, 2.3, 2.4, original label in Figure 2.5). The specimen illustrated on his figure 1 is an epichnion, and those in figures 3 and 4 are hypichnia on slab 5017076 (Figure 2.2).
The trace fossil on slab no. 5017075 (Figure 2.1) is composed of two rows of crescentic mound-like lobes. They are tilted by 3°-8° (on average 5°). They are partly shingled, gradually descending from an elevated apex towards the slightly convex, indistinct margin of the lobes. The elevated, slightly convex margins are 14-20 mm apart. The lobes are up to 40-45 mm long and perpendicularly oriented to the median, low-amplitude zigzag depression, which is 3-4 mm wide and turns at 140°±20°. The lobes are oriented concordantly in the same row. The opposing lobes alternate in position with a slight offset. All lobes are covered by thin ridges, less than 1 mm wide, which run concordantly to the slightly convex margin of the mound and become increasingly indistinct outward (Figure 2.1, 2.4). The ridges are a surface expression of the internal lamination of the lobes that consists of alternating layers of dominantly muddy and sandy material. The laminae are inclined. The course of the rows and the central depression, visible for a distance of 150 mm, ends at both sides by the edges of the slab. One side of one row is also broken by the margin of the slab. The slab is 25 mm thick and composed of medium-grained, quartz-dominated sandstone with parallel lamination (Bouma Tb interval) followed by slightly inclined, subparallel lamination in finer sandstone above (transition to Tc).
Specimens on slab 5017076 (Figure 2.2) are similar and generally poorly visible. They show a nearly straight or slightly winding to curved course. The specimen shown and labelled as (1) by Ooster (1869) on his plate 4, in fact, represents two specimens that cross each other (Figure 2.6). The most complete specimen is a winding ridge, obtusely triangular in cross-section, having ribs running slightly oblique from the ridge crest. The ribs are up to 13 mm long and located in alternating position on both sides of the ridge, up to 8 mm apart on one side. The ridge is up to 28 mm wide and at least 100 mm long. Specimens (3) and (4) shown by Ooster on plate 4 occur on the lower side of the same slab and are badly preserved; there another Polykampton specimen is present, but it was not illustrated by Ooster. The lower side is a rather smooth parting surface (Tb? division) and exhibits only badly preserved specimens due to the coarse grain size. Furthermore, this side was covered by plaster-of-Paris when the large specimens on the other side were shown in an exhibition. Unfortunately, the plaster-of-Paris was not completely removed afterward. The median tunnel is filled mainly by mudstone and it is surrounded by sandstone.
 Remarks. Because of the internal lamination of the lobes, they can be classified as spreite structures. Overlapping spreite laminae suggest that their formation started proximally in prolongation of the last curve of the median tunnel and continued to the other distal side. Multiple overlapping probes in the direction of movement, resulting in a spreite (-like) structure have also been observed for the trace fossil Hillichnus Bromley et al., 2003. However, the main difference to that ichnogenus is that, in Polykampton, no vertical tubes run to the seafloor. Ooster's sample 5017076 contains specimens on both sides (1, 3 and 4), but does not show cross-sections of such tubes on either side, nor on the vertical surfaces of the slab.
Remarks. Because of the internal lamination of the lobes, they can be classified as spreite structures. Overlapping spreite laminae suggest that their formation started proximally in prolongation of the last curve of the median tunnel and continued to the other distal side. Multiple overlapping probes in the direction of movement, resulting in a spreite (-like) structure have also been observed for the trace fossil Hillichnus Bromley et al., 2003. However, the main difference to that ichnogenus is that, in Polykampton, no vertical tubes run to the seafloor. Ooster's sample 5017076 contains specimens on both sides (1, 3 and 4), but does not show cross-sections of such tubes on either side, nor on the vertical surfaces of the slab.
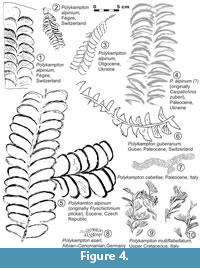
Some other ichnotaxa known from deep-sea sediments having the same morphological features are included in Polykampton alpinum (see the list of records, including synonyms), for instance Flyschichnium plickai Novák and Pelsová, 1983 (Figure 4.5) from the Eocene flysch of the Czech Carpathians. Carpatichnus zuberi Vialov, 1993 (Figure 4.4) from the Paleogene of the Carpathians in Ukraine probably belongs to P. alpinum. In addition, the unnamed trace fossil from the Oligocene Carpathian flysch in Ukraine (Makarenko, 1973; see Figure 4.3) belongs to P. alpinum. Fu (1991) mentioned that Polykampton occurs in the Upper Cretaceous-Paleogene of the Austrian Alps and Apennines, and the Carpathians should be added to that list.
Polykampton guberanum isp. nov.
Figure 4.6, Figure 5.1-5
zoobank.org/7374F14E-1A71-4AE5-B321-36AE1D7C23CF
List of records
v 1996 Polykampton alpinum Ooster – Rykart, p. 505, figure 4b. [Gubersandstein in the Schlierenflysch, upper Paleocene (Thanetian).]
 Derivation of name. From Guber quarry, the type locality.
Derivation of name. From Guber quarry, the type locality.
Diagnosis. Horizontal, straight, curved or winding structure composed of a median tunnel and lateral, crescentic, wing-like lobes arranged widely spaced in a row. The tips of lobes appear in alternating positions on both sides of the median tunnel that overlaps the lobes. Most of the lobes show an arcuate, concave, sharply outlined margin on one side and a convex, less distinct margin on the opposite side.
Holotype and paratypes. Specimen (Figure 5.2) in the middle of the slab NML 1995-11a (Figure 5.1) is defined as the holotype and is stored in the repository of the Naturmuseum Luzern, Switzerland. Five other specimens on this slab and their counterparts on the slab NML 1995-11b (shown in exhibition), as well as one specimen on slab O155 (Figure 5.5) housed in the Naturhistorisches Museum Basel are paratypes.
Type locality and horizon. Guber quarry (Switzerland), Schlieren Flysch Unit (upper Paleocene: Thanetian: nannoplankton NP8 Zone).
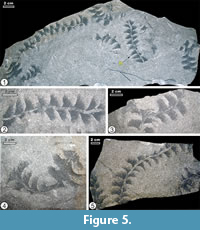
Description. The trace fossil was observed on parting surfaces of light-gray or grayish-brown, fine-to medium-grained sandstone slabs (Figure 6). It is a horizontal, straight, slightly bent to winding structure, 42-60 mm wide and up to 250 mm long. A tunnel is traversing the lobes nearly in the middle. The median tunnel is 2.0-5.5 mm wide. It is filled with sandstone and locally shows a thin mud lining and poorly expressed, sparse menisci due to higher content of clay. Rarely, the tunnel is branched (Figure 5.3). The branching shows lateral lobes with the same features as the other parts of the burrow. The lobes show a crescentic, wing-like shape. They are up to 11-18 mm wide and up to 28 mm long. From the proximal tip the lobes widen outward. The proximal part is gently arcuate, having at both sides concave, sharply bounded margins, while the distal part appears convex and has a less distinct, fuzzy margin. The lobes are filled with clayey to silty, dark-gray material and show a delicate spreite structure. The spreite laminae were produced first at the concave margin that is close to the median tunnel. Each lamina runs at a small deviating angle to the previous lamina to constitute the lobe. The inclination of the laminae increases distally in a fanlike mode. The tips of lobes appear in alternate position on both sides of the median tunnel. The lobes are asymmetrical and widely spaced, being about 14-22 mm apart. The traversing median tunnel divides a lobe into a large and a small part that alternate on each side of the median tunnel. In some burrow fragments, the tunnel is not seen at all or is only partly visible (Figure 5.4). The sandstone in vicinity of the lobes does not differ in composition from the sand farther away from the lobes.
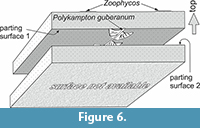 On the upper surface of slab NML 1995-11a in the Naturmuseum Luzern there are two spreite lobes of Zoophycos, which are up to 43 mm wide and up to 135 mm long. Polykampton guberanum occurs 30-55 mm below the surface of Zoophycos (surfaces 1, 2 in Figure 6). Unfortunately, the base of the sample (base of slab NML1995-11b) is not available for inspection as it is fixed to the exhibition wall. The slab O155 from the Naturhistorisches Museum Basel is 73 mm thick. Polykampton guberanum occurs on the upper surface and Ophiomorpha annulata (Książkiewicz) on the lower one.
On the upper surface of slab NML 1995-11a in the Naturmuseum Luzern there are two spreite lobes of Zoophycos, which are up to 43 mm wide and up to 135 mm long. Polykampton guberanum occurs 30-55 mm below the surface of Zoophycos (surfaces 1, 2 in Figure 6). Unfortunately, the base of the sample (base of slab NML1995-11b) is not available for inspection as it is fixed to the exhibition wall. The slab O155 from the Naturhistorisches Museum Basel is 73 mm thick. Polykampton guberanum occurs on the upper surface and Ophiomorpha annulata (Książkiewicz) on the lower one.
Remarks. Polykampton guberanum from the Guber quarry was first illustrated by Rykart (1996) and determined as P. alpinum, but it differs from the latter and P. cabellae by the widely spaced lobes and their triangular tips (Figure 4.6). Furthermore, P. guberanum is the only ichnospecies of Polykampton that exhibits only one lobe that is traversed by the median tunnel dividing it into two parts. In P. alpinum and P. cabellae (Figure 4.7), terminations of the lobes are semielliptical. Polykampton eseri (Unger, 1850), is much smaller and shows perpendicular, narrowly isolated, slightly arcuate lobes, with semielliptical terminations (Figure 4.8).
The fuzzy margin of the lobes of P. guberanum may partly result from the oblique cut of the spreite by the parting surface and the divergence of the probes in distal position. The parts of the burrow without visible median tunnel suggest that tunnel and lobes were produced at slightly different levels.
Slab NML 1995-11a in the Naturmuseum Luzern provides clear evidence that P. guberanum was produced at least 5.5 cm below the top of the sandstone bed (Figure 6), which was probably covered by the mud that now fills the lobes. Therefore, the tier of P. guberanum was probably located at least a decimeter below the sediment-water interface. It is not clear if it was formed below the tier occupied by Zoophycos, which occurs at the top of the slab, because it is also possible that Zoophycos was produced later originating from a bed above.
Geochemical data. The dark spreite-fill material of Polykampton guberanum is enriched in organic carbon (Corg: 3.05 %) and depleted in carbonate (CaCO3: 1.5 %) compared to the host sediment (Corg: 1.05 %; CaCO3: 28.8 %).
Polykampton multiflabellatum isp. nov.
Figure 4.9-10, Figure 7, Figure 8
zoobank.org/B286E14C-2839-4C45-B0D1-2F75FCD292F4
List of records
2016 Polykampton – Rodríguez-Tovar et al., p. 59, figure 6D. [Bolonia Unit, lower Paleocene, Campo de Gibraltar Complex, Cadiz Province, southern Spain.]
2017 Polykampton – Pujalte et al., p. 193, figure 6f. [Olivares Formation, Paleocene, southern Spain.]
Derivation of name. A combination of Latin multi-many, and flabellatum-fanlike.
 Diagnosis. Horizontal structure composed of a winding, meandering or rarely straight median tunnel and lateral fanlike lobes, which extend from the outer bends of the median tunnel. The lobes can extend upward above the level of the median tunnel.
Diagnosis. Horizontal structure composed of a winding, meandering or rarely straight median tunnel and lateral fanlike lobes, which extend from the outer bends of the median tunnel. The lobes can extend upward above the level of the median tunnel.
Holotype and paratypes. Specimen 7408 is the holotype. Paratypes 1 and 2 are specimens 6851 and 6852, respectively. All specimens are housed in the Museo Paleontologico di Crocefieschi, Italy.
Other specimens. Specimens 6853, 7061, 7061, 7331, 7332, 7386, 7449 in the Museo Paleontologico di Crocefieschi. Moreover, specimen NHMW 2018/0157/0001 in the Naturhistorisches Museum in Vienna (collected by Ernst Anton Leopold Kittl in 1888), which derived from the Kahlenberg Beds, Austria.
Type locality and horizon. Costa Gramigna 2 (GPS coordinates: N44º35.201'; E009º23.358'), Ottone Formation, Italy.
Description. Hypichnial structures on a sandstone bedding plane. The sinuous course of the meandering median tunnel is 2-5 mm wide. The meanders have a wavelength of 20-37 mm, and their amplitude ranges mainly from 2 to 6 mm, rarely up to 17 mm. The turns commonly exhibit angles of 130º-145º, rarely 100º. The median tunnel is either smooth or covered by longitudinal wrinkles. The lateral lobes are fan-shaped, 27-53 mm long, up to 40 mm wide. The fanlike lobes diverge at the base at an angle of 81°-155°, but in particular specimens the angle is 124°-134°, 97°-155°, 81°-95° or 82°-144°, respectively. Narrow, straight, gently curved sinuous ridges and grooves less than 1 mm wide cover the lobes. Both run concordantly parallel or subparallel along the lobe. The ridges and grooves can be arranged in narrow fanlike bundles (Figure 8.2) that may diverge distally. Most of the bundles are less than 5 mm wide. The lobes may expand slightly above the level of the median tunnel beyond the next meander. The median tunnel can be terminated by a lobe. The elevated edge of the lobe is gently curved backward. The direction of the elevated edge of the lobe and its internal groove-and-ridge arrangement may increasingly diverge with the distance to the edge. Locally, 1-2 mm long and about 0.5 mm wide oblique ridges cover the lobes and the median tunnel. These small ridges are most visible in some washed-out specimens (Figure 7.6, Figure 8.2). In some specimens, the median tunnel is clearly preserved, while only the proximal part of lobes is visible (Figure 7.4, 7.5). Aside from these features, polishing of lobes did not reveal their internal structure. On the upper side of the sandstone slabs, there is no evidence that burrow parts related to P. multiflabellatum occur underneath.
 Some of the lobes are covered with parallel/subparallel short, straight to slightly curved ridges, which are up to 0.3 mm wide and up to 3 mm long. They are oriented parallel or obliquely to the spreite laminae. Similar small ridges cover some segments of the tunnels. They are oriented obliquely to the tunnel on either side where they form a chevron pattern (Figure 7.6, Figure 8.1).
Some of the lobes are covered with parallel/subparallel short, straight to slightly curved ridges, which are up to 0.3 mm wide and up to 3 mm long. They are oriented parallel or obliquely to the spreite laminae. Similar small ridges cover some segments of the tunnels. They are oriented obliquely to the tunnel on either side where they form a chevron pattern (Figure 7.6, Figure 8.1).
Remarks. No vertical tubes have been observed in association with P. multiflabellatum and, hence, it cannot belong to Hillichnus Bromley et al., 2003. Polykampton multiflabellatum differs from P. alpinum by the large, horizontal, brush-like lobes, while P. eseri (Unger, 1850) is much smaller and shows perpendicular isolated lobes (Figure 8.8). Polykampton cabellae Uchman and Rattazzi, 2017 is also much smaller than P. multiflabellatum and shows closely spaced, shingled lobes (Figure 8.5). The small ridges that cover some lobes and ridges (Figure 7.6, Figure 8.1) can be interpreted as scratch traces produced in firmground by the tracemaker during feeding and locomotion.
 Polykampton multiflabellatum shows some similarities with Zonarides striatus Squinabol, 1888 from the Monte Antola Formation (late Campanian-Maastrichtian) in Liguria, Italy. Later it was transferred to Saportia Squinabol, 1891 as Saportia striata by Squinabol (1888) (Figure 9; see also Häntzschel, 1975, p. W102, figure 64.4). This trace fossil is only known from one very fragmentary and one larger specimen (= lectotype). Saportia is a hypichnial full relief, which shows a winding median tunnel with backward curved branches and proximal part of lateral lobes. The lobes are slightly inclined up (in stratigraphic sense), with a corrugate surface, and welded with the sandstone bedding surface. This trace fossil differs from Phycodes, which is composed of a common stem and bundles of tubes converging in the proximal part and diverging in the distal part (Häntzschel, 1975). Saportia shows some similarities to Polykampton multiflabellatum, but its median tunnel is 12-15 mm wide, i.e., 3-4 times wider than in P. multiflabellatum. This excludes synonymization of these ichnospecies. However, Saportia could be probably synonymized with Polykampton at the ichnogenus level, but more complete specimens than the lectotype studied here would be needed. The visibility of lobes depends on the level of intersection of the burrow system with the bedding plane.
Polykampton multiflabellatum shows some similarities with Zonarides striatus Squinabol, 1888 from the Monte Antola Formation (late Campanian-Maastrichtian) in Liguria, Italy. Later it was transferred to Saportia Squinabol, 1891 as Saportia striata by Squinabol (1888) (Figure 9; see also Häntzschel, 1975, p. W102, figure 64.4). This trace fossil is only known from one very fragmentary and one larger specimen (= lectotype). Saportia is a hypichnial full relief, which shows a winding median tunnel with backward curved branches and proximal part of lateral lobes. The lobes are slightly inclined up (in stratigraphic sense), with a corrugate surface, and welded with the sandstone bedding surface. This trace fossil differs from Phycodes, which is composed of a common stem and bundles of tubes converging in the proximal part and diverging in the distal part (Häntzschel, 1975). Saportia shows some similarities to Polykampton multiflabellatum, but its median tunnel is 12-15 mm wide, i.e., 3-4 times wider than in P. multiflabellatum. This excludes synonymization of these ichnospecies. However, Saportia could be probably synonymized with Polykampton at the ichnogenus level, but more complete specimens than the lectotype studied here would be needed. The visibility of lobes depends on the level of intersection of the burrow system with the bedding plane.
Stratigraphically, P. multiflabellatum ranges from Campanian (Ottone Formation; Italy) to Paleocene (occurrences in Spain).
DISCUSSION
The ichnogenus Polykampton comprises mainly horizontal trace fossils composed of a cylindrical median tunnel and complex, slightly inclined lateral lobes. The fanlike lobes of P. alpinum, P. guberanum and P. multiflabellatum result from closely spaced, partly overlapping probings, which led to the formation of spreite. The diameter of the median tunnel appears to be wider than that of the probings. The median tunnel cross-cuts the lobes.
Functional Morphology and Feeding Strategy
Polykampton alpinum, P. guberanum and P. multiflabellatum are deep-tier burrows formed below the tier containing Chondrites and maybe even below Zoophycos. The tracemaker was not restricted to a specific lithology because the trace fossils occur in sandstone, but also can cross the sandstone-mudstone interface. Therefore, the tracemakers appear to prefer a particular depth interval below the seafloor having specific geochemical conditions.
The spreite structures observed in the lobes of Polykampton alpinum, P. guberanum and P. multiflabellatum provide evidence of stripmining in the sand and in the muddy top of the turbidite bed underneath. The stripmining is seen as an expression of the systematic utilization of the sediment. The dark color of the fanlike lobes documents enrichment of fine-grained material that was intentionally introduced into the sand, while no sediment sorting is evident adjacent to the lobes. Organic carbon and carbonate data support the interpretation of temporary surface feeding of the Polykampton producer. The high carbonate content of the host sediment is due to the presence of bioclasts and carbonate cement within the introduced turbidite sand, while the low carbonate content of the mud implies its accumulation close or below the calcite compensation depth (e.g., Winkler, 1983). The high organic content of the spreite fill results from sequestration of nutritious material by the Polykampton producer transferred from the seafloor downward as there is no evidence of sorting of turbidite sand adjacent to the lobes. These findings rule out the assumption that organic matter buried in the sand was the feeding target. In fact, the sand is not depleted, but distinctly enriched in dark gray mud particles within the lobes. Therefore, it is interpreted that the producer collected organic-rich, fine-grained material from the sediment surface and stored it in the burrow, particularly in the lobes. The mud-enriched marginal parts of the median tunnel support this view. Later, during times when little food was available on the seafloor, the sequestered material was utilized by re-probing the lobes. Combining these findings implies that Polykampton alpinum, P. guberanum and P. multiflabellatum represent sequestration structures. Polykampton cabellae has already been interpreted as a sequestrichnion (Uchman and Rattazzi, 2018).
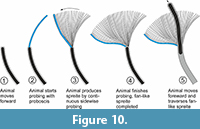 The tracemaker occupied the median tunnel, produced partly overlapping probes at varying angles to the course of the median tunnel, and finally formed the lobes (Figure 10). The diameter of the probings appears to be narrower than that of the median tunnel because the probes overlap and the median tunnel is lined and filled with sand after burrow abandonment. However, when preserved, the last-produced probe displays a similar width to that of the median tunnel. Nonetheless, it is suggested that the tracemaker very likely used an extendable, proboscis-like body appendage for probing while occupying the median tunnel at its end. After finishing a lobe and some forward movement, a new lobe was formed in an alternate position at the opposite side of the tunnel. The median tunnel was incidentally connected to the seafloor as the sediment from there was introduced somehow to the lobes, but repeated upward-directed tunnels have not been encountered. In some places, the median tunnel shows a laminated wall structure, which implies that the producers moved forward and backward while occupying the burrow.
The tracemaker occupied the median tunnel, produced partly overlapping probes at varying angles to the course of the median tunnel, and finally formed the lobes (Figure 10). The diameter of the probings appears to be narrower than that of the median tunnel because the probes overlap and the median tunnel is lined and filled with sand after burrow abandonment. However, when preserved, the last-produced probe displays a similar width to that of the median tunnel. Nonetheless, it is suggested that the tracemaker very likely used an extendable, proboscis-like body appendage for probing while occupying the median tunnel at its end. After finishing a lobe and some forward movement, a new lobe was formed in an alternate position at the opposite side of the tunnel. The median tunnel was incidentally connected to the seafloor as the sediment from there was introduced somehow to the lobes, but repeated upward-directed tunnels have not been encountered. In some places, the median tunnel shows a laminated wall structure, which implies that the producers moved forward and backward while occupying the burrow.
The bundles of spreite in P. multiflabellatum (Figure 8.2) may reflect multiphase stages of the lobe formation. A single bundle is produced during rather continuous activity of the tracemaker. Succession of the bundles records a lateral shift of the tracemaker activity. Similar width of the bundles indicates that more or less the same portion of sediment was mined and that the activity occurred rhythmically.
The observed pattern of P. guberanum suggests that the producers reworked the sediment within a lobe from one side to the other (Figure 10). Consecutive lobes were produced alternately on either side of the median tunnel. The fanlike spreite lobes and the undisturbed curvature of the probings in the connection to the median tunnel suggest that they were constructed while the tracemaker resided within the median tunnel.
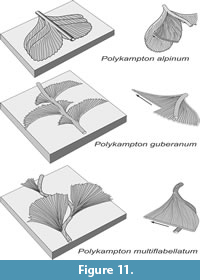 Polykampton alpinum and P. guberanum exhibit lobes that were formed slightly inclined downward from the level of the median tunnel. In P. alpinum, lobes are more or less of the same shape and size on both sides of the median tunnel, while in P. guberanum, each lobe is divided asymmetrically by the median tunnel. In P. multiflabellatum, the lobes were produced horizontally instead, and a new segment of the median tunnel was partly formed just below the level of the lobe (Figure 11). A new lobe was produced on the other side of the median tunnel. Wrinkles on the wall of the median tunnel suggest pressure from inside, which might have resulted from multiple utilization or passage of the animal, and possibly from its growth.
Polykampton alpinum and P. guberanum exhibit lobes that were formed slightly inclined downward from the level of the median tunnel. In P. alpinum, lobes are more or less of the same shape and size on both sides of the median tunnel, while in P. guberanum, each lobe is divided asymmetrically by the median tunnel. In P. multiflabellatum, the lobes were produced horizontally instead, and a new segment of the median tunnel was partly formed just below the level of the lobe (Figure 11). A new lobe was produced on the other side of the median tunnel. Wrinkles on the wall of the median tunnel suggest pressure from inside, which might have resulted from multiple utilization or passage of the animal, and possibly from its growth.
Similar to Polykampton, other deep-or middle-tier trace fossils are actively filled with organic-rich material from above and represent sequestrichnia. They include Zoophycos (e.g., Bromley, 1991; Löwemark et al., 2004 and references therein), Phymatoderma (Izumi, 2012, 2015), Cladichnus (Wetzel and Uchman, 2013), Rhizocorallium hamatum (Uchman and Rattazzi, 2017) or Tubulichnium rectum (Uchman and Wetzel, 2017). Due to such a nutrition strategy, the usually rapid microbial decomposition of organic-rich mud collected from the sea floor (e.g., Boetius and Lochte, 1994) is hindered and the nutritious mud is available for a prolonged period of time.
Tracemaker
Polykampton alpinum has been considered by Ooster (1869) as a hydrozoan, spawn ribbons of gastropods by Fuchs (1895), a possible trace fossil by Ehrenberg (1942) and finally as a feeding burrow by Seilacher (1959, 2007; also Häntzschel, 1975). The tracemakers are not known, but Makarenko (1973) considered certain beetles in sand in a continental setting or mollusks in a marine environment. Vialov (1993) supposed that such traces could be produced by the parapodia of polychaetes or the appendages of decapods. Fedchenko and Tatoli (1980) compared the form described by Makarenko (1973) to Mystichnis pacificus Vialov and Zenkevich, 1961. Vialov (1993) discussed similarities of this trace fossil to the so-called Pinsdorfer Versteinerung, which he named as Pinsdorfichnis abeli. Bivalves could be excluded because shell and siphon considerably differ in size that is not evident in Polykampton spreite lobes and median tunnel. Taking into account the fine stripmining, gentle curvature of the median tunnel and laminae, a flexible body with expandable proboscis is the very suitable for forming such structures. Wrinkles on the median tunnel in P. multiflabellatum are suggestive of a pressure from the interior. This can be seen as evidence that the body is laterally expandable. Therefore, a “worm”, probably a polychaete, is the most probable trace maker. Polychaetes show a large variety and plasticity in feeding (Fauchald and Jumars, 1979). Scratch traces in general are traditionally ascribed to crustaceans, but more recently they are also referred to “worms” (Knaust, 2013). Therefore, the scratch marks in P. multiflabellatum do not contradict the interpretation presented.
Paleoenvironment
Polykampton alpinum and P. guberanum found in Switzerland occur in rather thick-bedded turbiditic sandstones interpreted to have accumulated in a channelized part of middle-to upper-fan setting (van Stuijvenberg, 1979; Winkler, 1983). The associated trace fossils are typical of the deep-sea Nereites ichnofacies (Crimes et al., 1981). Polykampton eseri and P. cabellae have also been found only in turbiditic sediments; P. eseri preferably occurs in distal deep-sea fan settings in marly deposits (Wetzel and Uchman, 1997; Uchman, 1999), while P. cabellae was found 1.5-3.5 cm below the top of thick turbiditic marlstone beds, which are covered by a few centimeters of background mudstones (Uchman and Rattazzi, 2017).
In the same outcrop, P. multiflabellatum, Helminthopsis isp., Planolites isp. and ?Thalassinoides isp. are present. In a nearby outcrop of the Ottone Formation, Costa Gramigna 1 (GPS: N44º35.316'; E009º23.497'), Chondrites targionii, Ch. intricatus, Planolites isp., Urohelminthoida isp., Paleodictyon isp., Helminthorhaphe isp., Rotundusichnium zumayense, Scolicia isp. and Phycosiphon incertum have been found. This trace fossil assemblage is typical of the deep-sea Nereites ichnofacies (e.g., Uchman and Wetzel, 2012). The Kahlenberg Formation in Austria, wherein P. multiflabellatum also occurs, contains a typical deep-sea trace fossil assemblage (Uchman, 1999). The same is valid for the Spanish occurrences of this ichnospecies (Rodríguez-Tovar et al., 2016; Pujalte et al., 2017).
The environmental situation in channelized parts of modern deep-sea fans supports the interpretation of the remarkable fluctuations in availability of benthic food. Intermittent episodic currents carry fine-grained material and organic matter downslope, for instance, within the Nazaré and Congo deep-sea fan canyon-channel system (e.g., Oliveira et al., 2007; Vangriesheim et al., 2009).
CONCLUSIONS
The ichnogenus Polykampton Ooster, 1869 groups horizontal complex burrows composed of a horizontal median cylindrical tunnel and lateral spreite lobes that alternate on either side of the tunnel. In the type ichnospecies, P. alpinum Ooster, 1869 (Upper Cretaceous-Oligocene; Alps, Apennines, Carpathians), lobes are inclined to the bedding, partly shingled and closely spaced. Lobes of P. guberanum isp. nov. (Paleocene, Swiss Alps) are asymmetric and widely spaced. In P. multiflabellatum isp. nov. (Campanian-Paleocene; Apennines, Betic Cordillera) opposite to the other ichnospecies, the lobes are located above the median tunnel and extend from its outer bents. Polykampton is a deep-tier feeding structure (sequestrichnion) of a “worm”-like organism, which sequestered organic-rich sediment from the seafloor, stored it in the lobes and processed it later by stripmining. Polykampton occurs deep-sea, mainly turbiditic deposits. The sequestration is a good adaptation to generally low food availability and to its seasonal or incidental supply on the seafloor.
ACKNOWLEDGEMENTS
The research in Italy by A.U. was supported by the Fondazione Luigi, Cesare e Liliana Bertora. Additional support was provided by the Jagiellonian University. Research of A.U. in the Naturhistorisches Museum in Vienna received support from the SYNTHESIS Project http://www.synthesys.info, which is financed by the European Community Research Infrastructure Action under the FP6 "Structuring the European Research Area" Programme. A.W. was supported by the Swiss National Science Foundation (grant No. 200021_169042/1). U. Menkveld-Gfeller (Bern, Switzerland) provided access to the Polykampton type material and additional specimens housed in the Naturhistorisches Museum der Burgergemeinde Bern. B. Hotz (Luzern, Switzerland) made available Polykampton specimens exhibited and stored in the Naturmuseum Luzern. T. Nichterl (Vienna, Austria) provided information about specimens in the Naturhistorisches Museum in Wien, and A. Lukeneder (Vienna, Austria) provided additional photographs of Polykampton alpinum housed in this museum. A. Brugiglio (University of Genova, Italy) provided information and enabled inspection of Saportia striata (Squinabol) specimen in the Department of Earth, Environmental and Resources Science of University of Genova. M. Wiesner (Hamburg, FRG) provided measurements of organic carbon and carbonate content. C.A. Meyer (Basel, Switzerland) translated the abstract to French. D. Knaust (Stavanger, Norway) and three anonymous reviewers provided helpful comments, which enabled improvement of the paper. All these contributions are gratefully acknowledged.
REFERENCES
Boetius, A. and Lochte, K. 1994. Regulation of microbial enzymatic degradation of organic matter in deep-sea sediments. Marine Ecology Progress Series, 104:299-307. https://doi.org/10.3354/meps104299
Bromley, R.G. 1991. Zoophycos: strip mine, refuse dump, cache or sewage farm? Lethaia, 24:460-462. https://doi.org/10.1111/j.1502-3931.1991.tb01501.x
Bromley, R.G., Uchman, A., Gregory, M.R., and Martin, A.J. 2003. Hillichnus lobosensis igen. et isp. nov., a complex trace fossil produced by tellinacean bivalves, Paleocene, Monterey, California, USA. Palaeogeography, Palaeoclimatology, Palaeoecology, 192:157-186. https://doi.org/10.1016/S0031-0182(02)00684-3
Catanzariti, R. and Perilli, N. 2006. Age of the Ottone Unit in the Zignago Area (External Ligurian Units, Northern Apennines): Constraints from calcareous nannofossils. Ofioliti, 31:11-24.
Crimes, T.P., Goldring, R., Homewood, P., van Stuijvenberg, J., and Winkler, W. 1981. Trace fossil assemblages of deep-sea fan deposits, Gurnigel and Schlieren flysch (Cretaceous-Eocene). Eclogae Geologicae Helvetiae, 74:953-995.
Ehrenberg, K. 1942. Über einige Lebensspuren aus dem Oberkreideflysch von Wien und Umgebung. Paleobiologica, 7(1941):282-313.
Fauchald, K. and Jumars, P.A. 1979. The diet of worms: a study of polychaete feeding guilds. Oceanography and Marine Biology-An Annual Review, 17:193-284.
Faupl, P. 1975. Schwermineralien und Strömungsrichtungen aus den Kaumberger Schichten (Oberkreide) des Wienerwald-Flysches, Niederösterreich. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, 1975:538-450.
Faupl, P. 1976. Vorkommen und Bedeutung roter Pelite in den Kaumberger Schichten (Oberkreide) des Wienerwald-Flysches, Niederösterreich. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, 1975:538-450.
Fedchenco, Y.I. and Tatoli, I.A. 1980. Slied nieizviestnoho organisma na kainozoiskom piestchanike Karpat. Paleontologicheskiy Sbornik, 17:87-89.
Fu, S. 1991. Funktion, Verhalten und Einteilung fucoider und lophoctenoider Lebensspuren. Courier Forschungs-Institut Senckenberg, 135:1-79.
Fuchs, T. 1895. Studien über Fucoiden und Hieroglyphen. Denkschriften der Kaiserlichen Akademie der Wissenschaften Wien, mathematisch-naturwissenschaftliche Classe, 62:369-448.
Häntzschel, W. 1965. Vestigia invertebratorum et problematica, p. 1-140. In Westphal, F. (ed.), Fossilium Catalogus. Gravenhage, Junk.
Häntzschel, W. 1975. Trace Fossils and Problematica. In Teichert, C. (ed.), Treatise on Invertebrate Paleontology, Part W, Miscellanea, Supplement I. Geological Society of America and University of Kansas, New York and Lawrence.
Izumi, K. 2012. Formation process of the trace fossil Phymatoderma granulata in the Lower Jurassic black shale (Posidonia Shale, southern Germany) and its paleoecological implications. Palaeogeography, Palaeoclimatology, Palaeoecology, 353-355:116-122. https://doi.org/10.1016/j.palaeo.2012.07.021
Izumi, K. 2015. Composite Phymatoderma from Neogene deep-marine deposits in Japan: implications for Phanerozoic benthic interactions between subsurface burrows and the trace-makers of Chondrites and Phycosiphon. Acta Palaentologica Polonica, 60:1009-1020. https://doi.org/10.4202/app.00060.2014
Knaust, D. 2013. The ichnogenus Rhizocorallium: Classification, trace makers, palaeoenvironments and evolution. Earth-Science Reviews, 126:1-47. https://doi.org/10.1016/j.earscirev.2013.04.007
Löwemark, L., Lin, I., Wang, C., Huh, C., Wei, K., and Vhen, C. 2004. Ethology of Zoophycos producer: arguments against the gardening model from δ13Corg evidences of the spreiten material. Terrestrial, Atmospheric and Oceanic Sciences, 15:713-725.
Makarenko, D.E. 1973. Slid zagadkovogo organismy na oligotsenovomu piskoviku (Trace of mystery organism of the Oligocenian sandstone). Dopividi Akademii Nauk Ukrainskoi RSR, Seria B, Geologia, Geofizika, Chimia ta Biologia, 3:220-222. (In Ukrainian, with English summary)
Marroni, M., Monechi, S., Perilli, N., Principi, G., and Treves, B. 1992. Late Cretaceous flysch deposits of the Northern Apennines, Italy: age of inception of orogenesis-controlled sedimentation. Cretaceous Research, 13:487-504. https://doi.org/10.1016/0195-6671(92)90013-G
Marroni, M., Ottria, G., Pandolfi, G., Catanzariti, R., Bormioli, D., Cucchi, A., and Moletta, G. In press. Note Illustrative della Carta Geologica d’Italia alla scala 1: 50 000, Foglio 196, Cabella Ligure. Servizio Geologico d’Italia, Roma.
Matter, A., Homewood, P., Caron, C., Rigassi, D., van Stuijvenberg, J., Weidmann, M., and Winkler, W. 1980. Flysch and molasse of western and central Switzerland, p. 26 – 293. In Trümpy, R. (ed.), Geology of Switzerland -a Guide Book. Part B: Geological Excursions. Wepf, Basel.
Novák, Z. and Peslová, H. 1986. Flyschichnium plickai n. ichnogen. n. sp. – new fossil trace from the Upper Paleogene of the Carpathian flysch in Moravia (Czechoslovakia). Západne Karpaty, séria paleontológia, 11:99-107.
Oliveira, A., Santos, A.I., Rodrigues, A., and Vitorino, J. 2007. Sedimentary particle distribution and dynamics on the Nazaré canyon system and adjacent shelf (Portugal). Marine Geology, 246:105-122. https://doi.org/10.1016/j.margeo.2007.04.017
Ooster, W.-A. 1869. Die organische Reste der Zoophycos-Schichten der Schweizer Alpen, p. 15-35. In Ooster, W.-A. and von Fischer-Ooster, C. (eds.), Protozoe Helvetica. Mittheilungen aus dem Berner Museum der Naturgeschichte über merkwürdige Thier-und Pflanzenreste der schweizerischen Vorwelt, vol. 1. H. Georg, Basel.
Pujalte, V., Apellaniz, E., Caballero, F., Monechi, S., Ortiz, S., Orue-Etxebarria, X., Rodríguez-Tovar, F.J., and Schmitz, B. 2017. Integrative stratigraphy and climatic events of a new lower Paleogene reference section from the Betic Cordillera: Río Gor, Granada province, SE Spain. Spanish Journal of Palaeontology, 32:185-206.
Rodríguez-Tovar, F.J., Piñuela, L., and García-Ramos, J.C. 2016. Trace fossils assemblages from the Cenozoic “Flysch Units” of the Campo de Gibraltar Complex (southern Spain). Ichnos, 23:53-70. https://doi.org/10.1080/10420940.2015.1130708
Rykart, R. 1996. Spurenfossilien aus dem Gubersandstein, Schlierenflysch (OW). Schweizer Strahler, 10:503-507.
Seilacher, A. 1959. Zur ökologischen Charakteristik von Flysch und Molasse. Eclogae Geologicae Helvetiae, 51(1958):1062-1078.
Seilacher, A. 2007. Trace Fossil Analysis. Springer, Berlin, Heidelberg, New York.
Seilacher, A. and Mrinjek, E. 2011. Benkovac Stone (Eocene, Croatia): a deep-sea Plattenkalk? Swiss Journal of Geoscience, 104(Suppl. 1):S159-S166. https://doi.org/10.1007/s00015-011-0051-7
Squinabol, S. 1888. Contribuzioni alla flora fossile dei terreni terziari della Liguria. I. Fucoidi ed Elmintoidee. Bolletino della Societá Geológica Italiana, 6:545-561.
Squinabol, S. 1891. Contribuzioni alla flora fossile dei terreni terziari della Liguria. I. Alghe. Sordomuti, Genova.
Uchman, A. 1999. Ichnology of the Rhenodanubian Flysch (Lower Cretaceous-Eocene) in Austria and Germany. Beringeria, 25:65-171.
Uchman, A. and Rattazzi, B. 2017 (2016 online). Rhizocorallium hamatum (Fischer-Ooster, 1858), a Zoophycos-like trace fossil from deep-sea Cretaceous-Neogene sediments. Historical Biology, 29:395-410. https://doi.org/10.1080/08912963.2016.1167481
Uchman, A. and Rattazzi, B. 2018. (2017 online). The trace fossil Polykampton cabellae isp. nov. from the Pagliaro Formation (Paleocene), Northern Apennines, Italy: a record of nutritional sediment sequestration by a deep sea invertebrate. Ichnos 25:1-10. https://doi.org/10.1080/10420940.2017.1308362
Uchman, A. and Wetzel, A. 2012. Deep-sea fans, p. 643-671. In Bromley, R.G. and Knaust, D. (eds.), Trace Fossils as Indicators of Sedimentary Environments. Developments in Sedimentology, 64. Elsevier, Amsterdam.
Uchman, A. and Wetzel, A. 2017. Hidden subsurface garden on own faeces – the trace fossil Tubulichnium rectum (Fischer-Ooster, 1858) from the Cretaceous-Palaeogene deep-sea sediments. Palaeontologia Electronica, 20.2.40A:1-18. https://doi.org/10.26879/777
https://palaeo-electronica.org/content/2017/1968-tubulichnium-deep-sea-trace
Unger, F. 1850. Genera et species plantarum fossilium. Wilhelm Braumüller, Vindbonae.
Van Stuijvenberg, J. 1979. Geology of the Gurnigel area (Prealps, Switzerland). Beiträge zur Geologischen Karte der Schweiz, 151:1-111. Schweizerische Geologische Kommission, Bern.
Vangriesheim, A., Khripounoff, A., and Crassous, P. 2009. Turbidity events observed in situ along the Congo submarine channel. Deep-Sea Research II, 56:2208-2222. https://doi.org/10.1016/j.dsr2.2009.04.004
Vialov, O.S. 1993. Some interesting bilobate crawling trails, recent and fossil, p. 21-27. In Vialov, O.S. and Fedonkin, M.A. (eds.), Trace Fossils of Extinct Organisms. Nauka, Moskva. (In Russian)
Vialov, O.S. and Zenkevich, N.L. 1961. Trail of a crawling animal of the floor of the Pacific Ocean. Izvestiya Akademii Nauk SSR, Seriya Geologicheskaya, 1961(1):52-58.
Wetzel, A. and Uchman, A. 1997. Ichnology of deep-sea fan overbank deposits of the Ganei Slates (Eocene, Switzerland) – a classical flysch trace fossil locality studied first by Oswald Heer. Ichnos, 5:139-162.
Wetzel, A. and Uchman, A. 2013. Cladichnus parallelum isp. nov.-a new mid to deep-tier feeding burrow system. Ichnos, 20:120-128. https://doi.org/10.1080/10420940.2013.816301
Winkler, W. 1983. Stratigraphie, Sedimentologie und Sedimentpetrographie des Schlieren-Flysches (Zentralschweiz). Beiträge zur Geologischen Karte der Schweiz, Neue Folge, 158:1-105. Schweizerische Geologische Kommission, Bern.
Zuber, R. 1918. Flisz i nafta. Prace Naukowe, Dział II, 2:1-381. Towarzystwo dla Popierania Nauki Polskiej, Lwów.
