

 Termite activity in the mid-Cretaceous of Australia
Termite activity in the mid-Cretaceous of Australia
Article number: 27.3.a48
https://doi.org/10.26879/1420
Copyright Palaeontological Association, September 2024
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Appendices
Submission: 13 June 2024. Acceptance: 13 September 2024.
ABSTRACT
In this paper we provide the oldest evidence of termites in Australia, based on an aggregation of several hundred morphologically distinctive faecal pellets preserved as opalized casts from the Griman Creek Formation (Albian-Cenomanian: c. 100 Ma) at Lightning Ridge, New South Wales, Australia. This trace fossil extends the record of isopterans in Australia around 40 to 50 million years earlier than previously identified termite wing impressions, and indicates that this group was an active component of the detritivorous community in eastern Gondwanan terrestrial ecosystems by the mid-Cretaceous. The distinctive prismatic faecal pellets with hexagonal cross-sections (referable to Microcarpolithes hexagonalis Vangerow) were probably produced by kalotermitid or mastotermitid termites. The associated fossil plant assemblage indicates that the producers of the faecal pellets likely fed on conifer wood. Based on the distribution of extant termites, the climate of the Lightning Ridge area (Surat Basin) was probably warm and moist during the mid-Cretaceous. Recognition that termites were well established in Australian terrestrial ecosystems by the Albian-Cenomanian implies that vicariance may have been just as influential as trans-oceanic dispersal in determining the early distribution of major termite clades. Opalization of these delicate faecal pellets highlights the potential for further discoveries of three-dimensionally preserved soft or friable body and trace fossils in the Lightning Ridge opal deposits.
Stephen McLoughlin. Department of Palaeobiology, Swedish Museum of Natural History, P.O. Box 50007, S-104 05 Stockholm, Sweden. (Corresponding author) steve.mcloughlin@nrm.se
Artai A. Santos. Department of Palaeobiology, Swedish Museum of Natural History, P.O. Box 50007, S-104 05 Stockholm, Sweden. ArtaiAnton.SantosLopez@nrm.se, artaisl29@gmail.com
Sherri Donaldson. Palaeoscience Research Centre, University of New England, Armidale, NSW, 2351, Australia. sdonal24@myune.edu.au
Christian Pott. LWL-Museum für Naturkunde, Westfälisches Landesmuseum mit Planetarium, Sentruper Straße 285, D-48161 Münster, Germany. Christian.Pott@lwl.org
Matthew McCurry. Earth and Sustainability Science Research Centre, School of Biological, Earth and Environmental Sciences (BEES), University of New South Wales, Kensington, New South Wales 2052, Australia; Australian Museum Research Institute, 1 William Street, Sydney, New South Wales 2010, Australia. Matthew.McCurry@Australian.Museum
Keywords: Isoptera; coprolites; opalization; plant-insect interactions; Kalotermitidae, Mastotermitidae
Final citation: McLoughlin, Stephen, Santos, Artai A., Donaldson, Sherri, Pott, Christian, and McCurry, Matthew. 2024. Termite activity in the mid-Cretaceous of Australia. Palaeontologia Electronica, 27(3):a48.
https://doi.org/10.26879/1420
palaeo-electronica.org/content/2024/5339-opalized-termite-coprolites
Copyright: October 2024 Palaeontological Association.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
Termites (infraorder Isoptera) are detritivorous insects that play a major role in nutrient cycling in various terrestrial ecosystems on all continents except Antarctica (Eggleton, 2011). They are eusocial insects, known for constructing some of the most complex colonial habitats in the animal kingdom (Noirot, 1970). In turn, numerous ancient and modern invertebrates and vertebrates have adapted to feed specifically or predominantly on termites (Sheppe, 1970; Redford, 1987; Luo and Wible, 2005). Termites and termitophagous animals are prominent components of northern Australian ecosystems; yet, despite their key ecological roles in the landscape, very little is known about the origins of Australasian representatives of this group.
On a global basis, the sedimentary record has yielded numerous occurrences of termite body fossils, coprolites, nests, and borings structures from a broad array of Cenozoic and Cretaceous deposits (e.g., Colin et al., 2011; Engel et al., 2011; Jouault et al., 2021), indicating that the importance of Isoptera in terrestrial ecosystems extends far back in time. Nevertheless, the fossil record highlights some important palaeogeographical and temporal gaps that inhibit reconstruction of the evolutionary history of this group. The origin of termites remains controversial. Classically, a Late Jurassic origin has been proposed for the divergence of Isoptera from their cryptocercid cockroach sister group (Engel et al., 2009; Bourguignon et al. 2014; Legendre et al., 2015). Some controversial features interpreted as termite nests from Upper Triassic and Lower Jurassic deposits have been used to suggest a more ancient origin (Hasiotis and Dubiel, 1995; Bordy et al., 2004, 2009, 2010) but the attribution of these structures to Isoptera is disputed (Genise et al. 2005; Genise, 2017) and some may represent megarhizoliths (Alonso-Zarza et al., 2008; Genise, 2017). An Early Jurassic age for the stem-termite origin and a Late Jurassic age for the crown-termite origin have been proposed based on combined molecular and fossil data (Legendre et al., 2015). However, other molecular and fossil data have suggested the possibility of a crown isopteran origin around the Triassic-Jurassic boundary (c. 200 Ma) or even older (Davis et al., 2009; Ware et al., 2010; Jouault et al., 2021). The disparity in results regarding the origin and diversification of termites makes the fossil record crucial for understanding the evolutionary history of this important group of insects. Thus far, the oldest convincing body fossils of termites were recovered from Lower Cretaceous deposits of Europe (see Appendix 1), specifically from the Berriasian Doronino Formation (Vršanský and Aristov, 2014) and Zaza Formation (Engel et al., 2007), Russia. The earliest evidence of isopteran coprolites derives from the Berriasian Missão Velha Formation of Brazil (Pires and Sommer, 2009), and over 40 other examples of fossil termite faecal pellets have been documented (see Appendix 2).
Cretaceous body fossils or coprolites of termites are relatively common in Europe (Jarzembowski, 1981; Lacasa-Ruiz and Martínez-Delclós, 1986; Schlüter, 1989; Engel et al., 2007, 2011; Engel and Delclòs, 2010; Colin et al., 2011; Engel, 2014; Sánchez-García et al., 2020), Asia (Ren, 1995; Krishna and Grimaldi, 2003; Engel et al., 2007, 2011, 2016; Colin et al, 2011; Zhao et al., 2020; Jouault et al., 2021, 2022a, 2022b; Jouault and Nam, 2023; Engel and Jouault, 2024: Jiang et al., 2024), and the Americas (Krishna, 1990; Fontes and Vulcano, 1998; Krishna and Grimaldi, 2000; Martins-Neto et al., 2006; Bechly, 2007; Grimaldi et al., 2008). There is also evidence of hexagonal isopteran coprolites in various Cretaceous deposits across Africa (Colin et al., 2011 and references therein). By contrast, the Australasian fossil record of Isoptera is depauperate. Previously, the oldest fossil isopteran from this region was a single fragmentary termite forewing described by Riek (1952) from the Paleocene-Eocene (58.5-55 Ma: Langford et al., 1995) Redbank Plains Formation near Ipswich, Queensland, and assigned to Blattotermes neoxenus Riek (Mastotermitidae). Kaulfuss et al. (2010) described another solitary forewing from Lower Miocene deposits of Foulden Maar, Otago, New Zealand, assigning it to Stolotermes kupe Kaulfuss, Harris et Lee (Stolotermitidae). McCurry et al. (2022) reported two additional termite (Mastotermitidae) wings from a thinly laminated goethitic siltstone of latest Early to earliest Late Miocene age (c. 16-11 Ma) near Talbragar in central New South Wales. Thus far, these are the only body fossil records of Isoptera from Australasia. In the absence of a rich body fossil record, trace fossils (coprolites, tunnels in wood, and nest structures) offer scope for elucidating the ancient history of termites in this region.
Australia has an extensive record of Cretaceous woods, albeit that few of these materials have been studied and most investigations have focused on the taxonomy and palaeoclimatic implications of these remains. Silicified conifer wood is abundant in eastern Australian Cretaceous basins (White, 1961; Frakes and Francis, 1990; Dettmann et al., 1992; Philippe et al., 2004; Fletcher et al., 2014, 2015; Tosolini et al., 2018). This abundance is probably linked to the release of permineralizing silica from the weathering of volcanic ash derived from the Whitsunday Igneous Province that was emplaced during the breakup of Australia and New Zealand.
In some marine deposits, the wood is entombed in calcareous concretions (Frakes and Francis, 1990) and, in others, (e.g., Western Australian marginal basins) the wood is preserved by silicification and phosphatization via the decomposition of glauconite and apatite in greensands (Simpson, 1912; McLoughlin et al., 1995b; McLoughlin, 1996; McLoughlin and McNamara, 2001; Mory and Hocking, 2017). Mummified, coalified, and charcoalified woods are also represented in various Australian basins but this material has been understudied (Backhouse et al., 1995; McLoughlin et al., 2002; Carpenter et al., 2016; Tosolini et al., 2018). Thus, there is considerable potential for investigation of termite traces in Australian fossil wood preserved in multiple styles.
Woody axes can have long taphonomic trajectories, acquiring biogenic (bacterial, fungal, arthropod, and molluscan) damage while the tree is alive, as deadwood in the form of stumps and logs on land, during transport in freshwater systems, and as driftwood or submerged log grounds after being washed into marine or lacustrine environments (Philippe et al., 2022). Although some of the published Australian Cretaceous woods contain evidence of fungal attack, especially in the form of pocket rot (McLoughlin, 1996), and other examples host borings from teredinid bivalves (McLoughlin et al., 1995b) and oribatid mites (Fletcher and Salisbury, 2014), none of the Cretaceous axes has yet revealed termite damage.
Australasian Cenozoic woods have yielded at least two examples of termite faecal pellets within tunnels excavated in wood that is now permineralized. Well-preserved examples include Neotermes -like fossil frass documented from mid-Cenozoic permineralized plant litter in central Queensland, Australia (Rozefelds and De Baar, 1991), and Kalotermes faecal pellets recorded in Miocene Avicennia wood from North Island New Zealand (Sutherland, 2003). Sub-fossil (Pleistocene) termite mounds have also been documented from stabilized sand dune systems in the Simpson Desert, central Australia (Miller, 1989).
There are over 360 extant species of termites in Australia within about five families (Watson and Abbey, 1993). Many Australian species are endemic. They are important ecosystem engineers as various groups have selective feeding patterns and abilities to consume and degrade leaf litter, grass, wood, fungi and diffuse organic matter in soil (Clement et al., 2021), liberating nitrogen, phosphorus and potassium that plants can exploit (Griffiths et al., 2021) and, in the process, contributing to c. 1-3% of the atmospheric CH4 budget (Sanderson, 1996; Nauer et al., 2018). Extant wood and humic soil-feeding termites are roughly constrained to latitudes between 45° N and 45° S (Hellemans et al., 2022). They are particularly abundant and diverse in moist, warm (tropical) climates, where they constitute c. 10% of animal biomass and are responsible for >55% of organic matter decomposition (Bignell, 2006; Jones and Eggleton, 2011; Griffiths et al., 2019, 2021). They do not live in cool or cold habitats (Sanderson, 1996; Atlas of Living Australia, 2024).
Here, we document opalized casts of clusters of prismatic coprolites from the Albian-Cenomanian Griman Creek Formation at Lightning Ridge that are interpreted to represent the oldest evidence for termites in Australia. We evaluate the probable producers of the coprolites, their importance for interpreting the early radiation of termites, and their role in nutrient recycling in mid-Cretaceous ecosystems.
GEOLOGICAL SETTING
Opal is Australia’s official national gemstone, with a significant percentage of commercial extraction occurring at Lightning Ridge in northern New South Wales (Department of Industry, Science and Resources, Commonwealth of Australia, 2022). Both precious and non-precious opal has been exploited from this area for over 100 years, with many examles representing replacements of the hard parts of Cretaceous organisms.
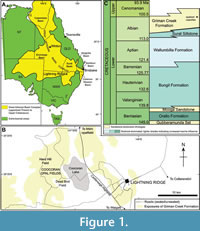 The area around Lightning Ridge (centred on 29°25′39″S, 147°58′44″E) is characterized by a low-lying NW-SE trending mesa system in the southern part of the Surat Basin (Herrmann and Maas, 2022; Figure 1A, B). The Griman Creek Formation (Rolling Downs Group) is a sedimentary succession up to 400 m thick, comprising generally thin-bedded, interlaminated, fine- to medium-grained sandstone, siltstone, mudstone, and minor coals, intercalated with thicker (c. 1-1.5 m thick) cross-bedded sandstones and conglomerates (Green et al., 1997; Payenberg and Reilly, 2004; Bell et al., 2019). The Griman Creek Formation at Lightning Ridge has been divided into two members: the Wallangulla Sandstone Member, comprising chiefly fine-grained, laminated, clay-rich sandstone with discontinuous claystone lenses of the informal ‘Finch clay facies’; and the overlying Coocoran Claystone Member, a claystone unit ≤10 m thick (Moore, 2002; Bell et al., 2019). The formation was deposited in low-energy freshwater fluvial and lacustrine systems (coastal plain environments) draining into the epeiric Eromanga Sea to the west (Bell et al., 2019). Laser Ablation Multicollector Inductively Coupled Plasma Mass Spectrometry U-Pb-Th radioisotopic dating of detrital zircons recovered from a volcanogenic bentonitic clay bed overlying the ‘Finch clay facies’ at the Hard Hill locality, yielded a range of Cretaceous depositional ages, with opalization of the formation occurring much later, during the Neogene (Bell et al., 2019; Mustoe and Smith, 2023; Figure 1C). More specifically, the suite of clear euhedral to elongate prismatic zircon grains with oscillatory magmatic zoning yielded 238U/206 Pb ages of 117±1.75 to 96.9±1.49 Ma (mid-Aptian to mid-Cenomanian), with the youngest coherent set of grains dated as 98.08+1.1/−2 Ma (early Cenomanian: at 95% confidence) and the youngest graphical peak in the population being 101 Ma (latest Albian: Bell et al., 2019). Thus, we interpret the age of the underlying ‘Finch clay facies’ of the Wallangulla Sandstone Member to approximate that of the Albian-Cenomanian boundary (hereafter, informally designated mid-Cretaceous; Figure 1CFigure 1C).
The area around Lightning Ridge (centred on 29°25′39″S, 147°58′44″E) is characterized by a low-lying NW-SE trending mesa system in the southern part of the Surat Basin (Herrmann and Maas, 2022; Figure 1A, B). The Griman Creek Formation (Rolling Downs Group) is a sedimentary succession up to 400 m thick, comprising generally thin-bedded, interlaminated, fine- to medium-grained sandstone, siltstone, mudstone, and minor coals, intercalated with thicker (c. 1-1.5 m thick) cross-bedded sandstones and conglomerates (Green et al., 1997; Payenberg and Reilly, 2004; Bell et al., 2019). The Griman Creek Formation at Lightning Ridge has been divided into two members: the Wallangulla Sandstone Member, comprising chiefly fine-grained, laminated, clay-rich sandstone with discontinuous claystone lenses of the informal ‘Finch clay facies’; and the overlying Coocoran Claystone Member, a claystone unit ≤10 m thick (Moore, 2002; Bell et al., 2019). The formation was deposited in low-energy freshwater fluvial and lacustrine systems (coastal plain environments) draining into the epeiric Eromanga Sea to the west (Bell et al., 2019). Laser Ablation Multicollector Inductively Coupled Plasma Mass Spectrometry U-Pb-Th radioisotopic dating of detrital zircons recovered from a volcanogenic bentonitic clay bed overlying the ‘Finch clay facies’ at the Hard Hill locality, yielded a range of Cretaceous depositional ages, with opalization of the formation occurring much later, during the Neogene (Bell et al., 2019; Mustoe and Smith, 2023; Figure 1C). More specifically, the suite of clear euhedral to elongate prismatic zircon grains with oscillatory magmatic zoning yielded 238U/206 Pb ages of 117±1.75 to 96.9±1.49 Ma (mid-Aptian to mid-Cenomanian), with the youngest coherent set of grains dated as 98.08+1.1/−2 Ma (early Cenomanian: at 95% confidence) and the youngest graphical peak in the population being 101 Ma (latest Albian: Bell et al., 2019). Thus, we interpret the age of the underlying ‘Finch clay facies’ of the Wallangulla Sandstone Member to approximate that of the Albian-Cenomanian boundary (hereafter, informally designated mid-Cretaceous; Figure 1CFigure 1C).
Although there is very little surface expression of the Griman Creek Formation in this region, fossils have been discovered in lenses of the ‘Finch clay facies’, extracted from small-scale subsurface mining activities since the early 1900s (Meakin, 2011). Many of the opal occurrences fill tectonic or dissolution voids in the ‘Finch clay facies’ but some represent casts of invertebrate shells vertebrate bones or robust plant organs. These do not preserve the calcium carbonate, calcium phosphate, or cellulose/lignin of the original animal and plant remains but represent infilling of voids left by dissolution of the organisms’ hard parts. Dissolution and subsequent infilling of the voids by opaline silica is inferred to be primarily a Neogene phenomenon associated with the development of deep weathering profiles across the region (Herrmann and Maas, 2022; Mustoe and Smith, 2023).
The opalized biota of the Griman Creek Formation is considered to represent one of the most important Cretaceous terrestrial fossil assemblages in Australia, containing a diverse array of vertebrate, invertebrate, and plant fossils preserved as opalized casts (Bell et al., 2019). The rich array of vertebrate remains includes both freshwater and amphibious animals: amiid fishes, dipnoans, lamniform chondrichthyans, chelids, crocodylomorphs, and elasmosaurid plesiosaurs (Kemp and Molnar, 1981; Kear, 2006; Smith, 2010; Smith and Kear, 2013; Hart et al., 2019; Kemp and Berrell, 2020; Berrell et al., 2023). Terrestrial vertebrate fossils include theropods (Bell et al., 2016; Birch et al., 2019, 2020), sauropods (Frauenfelder et al., 2021), ornithopods (Molnar and Galton, 1986; Bell et al., 2018a, 2018b; Kitchener et al., 2019), enantiornithine birds (Molnar, 1999), and monotreme mammals (Archer et al., 1985; Clemens et al., 2003; Flannery et al., 2024). Freshwater invertebrate remains include bivalve and gastropod molluscs, and a single clade of decapod crustaceans (Newton, 1915; Hocknull, 2000; Hamilton-Bruce et al., 2002, 2004; Hamilton-Bruce and Kear, 2006, 2010; Kear and Godthelp, 2008; Bell et al., 2020). The fossil plant suite from the Griman Creek Formation is currently under study by the authors and includes various equisetaleans, ferns, and cupressacean and araucarian conifers, but lacks obvious angiosperm remains (Smith and Smith, 1999).
MATERIAL AND METHODS
The single available specimen was photographed using an Apple iPhone 12 dual camera system with primary 12 megapixel sensor with 1.4 µm pixels and 26 mm equivalent f/1.6 lens. SEM micrographs were acquired of the uncoated specimen using a JEOL NeoScope JCM-7000 Benchtop SEM at the Australian Museum, Sydney, employing an acceleration voltage of 15.0 kv under low-vacuum mode, with EDS acquisition under high-vacuum mode. Tomography was undertaken using a Nikon XT H 225 ST X-ray CT system at 130 kV and 70 µA with a resulting voxel size of 6.069 µm that was reconstructed in VGstudio Max V2023.3. The studied specimen is registered as AM F 128064 and stored at the Australian Museum, Sydney, where it was received as a donation by E. Smith in 2004.
SYSTEMATIC PALEONTOLOGY
IchnogenusMICROCARPOLITHES Vangerow, 1954
Type ichnospecies. Microcarpolithes hexagonalis Vangerow, 1954; by subsequent designation of Hall (1963).
Microcarpolithes hexagonalis Vangerow, 1954
Figure 2A-J
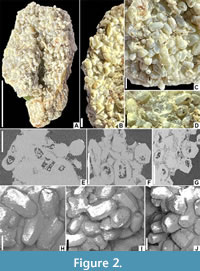 Description. The specimen consists of an aggregation of several hundred hexagonally prismatic opaline pellets; the block is 27 mm long, 16 mm wide, and 10 mm thick (Figure 2A-D). Individual pellets are oblong, 0.97-(1.33)-1.62 mm long (n=40), 0.59-(0.76)-1.00 mm in transverse diameter (n=40), and conspicuously hexagonal in cross-section (Figure 2E-G), with smoothly faceted surfaces (Figure 2H-J), and truncate to broadly rounded ends. Tomographic analysis reveals that individual coprolites have either a uniform finely granular internal composition (Figure 2E) or have an equivalent uniform outer rind and an internal cavity or porous region constituting one- to two-thirds of the pellet diameter (Figure 2F-G). The cavities locally contain irregularly arranged minute flecks of detritus. The coprolites lack any obvious regular organization or common orientation (Figure 2C). They are bound together by porous interstitial clays or opaline silica (Figure 2D, I, Figure 3).
Description. The specimen consists of an aggregation of several hundred hexagonally prismatic opaline pellets; the block is 27 mm long, 16 mm wide, and 10 mm thick (Figure 2A-D). Individual pellets are oblong, 0.97-(1.33)-1.62 mm long (n=40), 0.59-(0.76)-1.00 mm in transverse diameter (n=40), and conspicuously hexagonal in cross-section (Figure 2E-G), with smoothly faceted surfaces (Figure 2H-J), and truncate to broadly rounded ends. Tomographic analysis reveals that individual coprolites have either a uniform finely granular internal composition (Figure 2E) or have an equivalent uniform outer rind and an internal cavity or porous region constituting one- to two-thirds of the pellet diameter (Figure 2F-G). The cavities locally contain irregularly arranged minute flecks of detritus. The coprolites lack any obvious regular organization or common orientation (Figure 2C). They are bound together by porous interstitial clays or opaline silica (Figure 2D, I, Figure 3).
 Remarks. The structures described in this study differ from replacements of euhedral quartz crystals by their bluntly rounded termini (at both ends of the pellets) and irregular arrangement. These mid-Cretaceous structures are interpreted to be termite coprolites based on their equivalency in shape and cross-section to termite coprolites assigned to Microcarpolithes hexagonalis Vangerow, from the Late Cretaceous of the Netherlands (Vangerow, 1954; Colin et al., 2011). Microcarpolithes hexagonalis was originally considered to represent angiosperm seeds but later reinterpreted by Kovach and Dilcher (1988) as termite coprolites (Colin et al., 2011; Moreau et al., 2019). Here we follow the criteria of Colin et al. (2011) who proposed that Microcarpolithes hexagonalis “must be used to uniquely designate termite coprolites that are cylindrical in shape and with a conspicuous hexagonal section, making it clearly distinct from some coprolites of curculionid weevil larvae that occasionally produce subhexagonal pellets.” Although the Australian Cretaceous coprolites are slightly longer than the type specimen of M. hexagonalis from Europe, we do not regard this small difference (less than 1 mm) to be taxonomically significant for a trace fossil, which is in line with the proposal that size should not be considered primary criteria for differentiating ichnotaxa (Bertling et al., 2022).
Remarks. The structures described in this study differ from replacements of euhedral quartz crystals by their bluntly rounded termini (at both ends of the pellets) and irregular arrangement. These mid-Cretaceous structures are interpreted to be termite coprolites based on their equivalency in shape and cross-section to termite coprolites assigned to Microcarpolithes hexagonalis Vangerow, from the Late Cretaceous of the Netherlands (Vangerow, 1954; Colin et al., 2011). Microcarpolithes hexagonalis was originally considered to represent angiosperm seeds but later reinterpreted by Kovach and Dilcher (1988) as termite coprolites (Colin et al., 2011; Moreau et al., 2019). Here we follow the criteria of Colin et al. (2011) who proposed that Microcarpolithes hexagonalis “must be used to uniquely designate termite coprolites that are cylindrical in shape and with a conspicuous hexagonal section, making it clearly distinct from some coprolites of curculionid weevil larvae that occasionally produce subhexagonal pellets.” Although the Australian Cretaceous coprolites are slightly longer than the type specimen of M. hexagonalis from Europe, we do not regard this small difference (less than 1 mm) to be taxonomically significant for a trace fossil, which is in line with the proposal that size should not be considered primary criteria for differentiating ichnotaxa (Bertling et al., 2022).
DISCUSSION
Prismatic faecal pellets with sharply hexagonal cross-sections are atypical among insects, being produced by some groups of termites and some anobiid beetles, although those produced by coleopterans are typically much larger and less regular than pellets produced by isopterans (Weidner, 1956; Sutherland, 2003). Termopsidae, Kalotermitidae Archotermopsidae, and Mastotermitidae termites generate oblong pellets (Rohr et al., 1986; Colin et al., 2011; Moreau et al., 2019; Dong et al., 2022), whereas some other termite clades (e.g., Rhinotermitidae) produce amorphous, liquid, or gelatinous faeces (Arquette and Rodriguez, 2011; Colin et al., 2011). Termopsid pellets are generally larger and less regular than those of kalotermitids and mastotermitids (Light, 1934), and they lack surficial striate. Kalotermitids and mastotermitids are the most common producers of prismatic/faceted (hexagonal in cross-section) faecal pellets (Colin et al., 2011; Lewis et al., 2014) and, consequently, are the most likely producers of the Australian Cretaceous coprolites. The fecal pellets of some extant Kalotermitidae ( Kalotermes ) are smaller than those produced by the single extant species of Mastotermitidae ( Mastotermes darwinensis Froggatt). Although the mid-Cretaceous coprolites described here are more consistent with the size of the pellets produced by M. darwinensis (1-1.2 mm long according to Colin et al. 2011), the dimensional range and shapes of the Lightning Ridge coprolites are not sufficiently distinctive to exclude production by either mastotermitid or kalotermitid termites.
The hexagonal cross-section of termite dung is generated via pressure from six muscle bands that squeeze moisture out of the faecal mass in the rectum in order to collect water in the rectal grooves for retention by the insect (Child, 1934). The precise shape of the faecal pellet may also be influenced by the diet items and their moisture content (Lance, 1946). The cavities within some of the Lightning Ridge coprolites (Figure 2E-G) probably reflect incomplete opal casting of the pellet rather than constituting a primary textural or compositional character.
We interpret the dense arrangement of the Lightning Ridge coprolites to be the result of packing within a confined space, e.g., in a gallery formerly excavated within wood or within a termite mound or subterranean tunnel. The kalotermitid faecal pellets described from mid-Cenozoic (?Oligocene) deposits of central Queensland occur as masses in galleries within conifer wood or as free masses preserved in a silicified rainforest leaf litter assemblage (Rozefelds and De Baar, 1991). Those pellets are similar to the Lightning Ridge coprolites in being oblong and hexagonal in cross-section, but they are slightly larger (2.0-2.6 mm long and 0.8-1.0 mm wide). However, pellet size may be influenced by the availability and type of food or other ecological constraints (Rozefelds and De Baar, 1991). Apart from the slightly smaller size and more sharply defined hexagonally prismatic form of the Lightning Ridge coprolites, there is little to differentiate these from the central Queensland material, which supports a kalotermitid affinity for the new material and denotes a long (>100-million-year) history for this isopteran family in Australia.
The fossil flora of the Lightning Ridge opal fields is currently under study by the authors. Preliminary results indicate that the Albian-Cenomanian woody vegetation of this area was dominated by cupressacean and araucariacean conifers. The Lightning Ridge plant assemblage shares several taxa with the slightly younger Winton Formation flora of the Eromanga Basin to the north, in western Queensland (McLoughlin et al., 1995a, 2010). The former differs mainly in the apparent absence of angiosperms; hence, we infer that, like the Oligocene examples from Queensland (Rozefelds and De Baar, 1991), the Lightning Ridge mid-Cretaceous termites were probably feeding primarily on coniferous wood.
The Lightning Ridge specimen is not the only example of an opalized mass of termite coprolites. Rogers (1928, 1938) documented termite coprolites in Pliocene opalized wood from Santa Barbara, California. They are of similar size and shape to the Lightning Ridge material but more loosely aggregated and bound by interstitial opal and colloform lussatite.
Given that extant termites do not live in cool or cold habitats, it is likely that the climate experienced at Lightning Ridge during the late Early Cretaceous was fairly warm, despite the region’s relatively high (c. 60° S) palaeolatitude (Burgener et al., 2023). In addition, permineralized woods preserved at Lightning Ridge have marked growth ring boundaries suggesting strong seasonality in productivity. A relatively warm climate is supported by the presence of crocodilians and the diverse range of fossil organisms found in the opalized assemblages at Lightning Ridge (Smith and Smith, 1999; Bell et al., 2019). This is consistent with inferred global stepwise warming through the Albian-Turonian interval (Dettmann et al., 1992; Forster et al., 2007; Bottini and Erba, 2018; Huber et al., 2018; Burgener et al., 2023).
In North America, at least one archaic mammal ( Fruitafossor windscheffeli Luo and Wible, 2005) had, by the Late Jurassic, developed forelimb adaptations for digging and specialized dentition consistent with feeding on termites, other insects or soft plant matter. However, similar adaptations can develop among vermivorous mammals (Charles et al., 2013). Among extant Australian animals, the numbat ( Myrmecobius fasciatus : Marsupialia), short-beaked echidna ( Tachglossus aculeatus : Monotremata), and at least 16 species of lizards specialize in feeding on termites (Abensperg-Traun and Steven, 1997). As yet, no lizard or marsupial fossils have been recovered from the Griman Creek Formation at Lightning Ridge. Recent work has uncovered six species of monotremes from this deposit (Flannery et al., 2024). Among these, Opalios splendens Flannery et al. 2024 has some features reminiscent of adaptations to termitivory (e.g., long, slender jaws, reduced teeth with simplified crown shape, and inter-molar or -premolar diastemata) although these might, alternatively, have been adaptations to feeding on vermiform invertebrates. Although isopterans were clearly widespread by this time, whether any vertebrates targeted termites as a food source as early as the mid-Cretaceous in Australia remains speculative.
Mastotermitids and kalotermitids are relatively basal in at least some phylogenies of Isoptera, suggesting divergence early in the history of this order (Legendre et al., 2008; Lo and Eggleton, 2011). Owing to the relatively poor body fossil record of termites and recent discoveries of isopteran body and trace fossils extending back to, at latest, the Early Cretaceous on multiple continents (Jarzembowski, 1981; Rohr et al., 1986; Martínez-Delclòs and Martinell, 1995; Francis and Harland, 2006; Pires and Sommer, 2009; Colin et al., 2011; Dong et al., 2022; Greppi et al., 2023), we suggest that the ancient distribution of many termite clades remains inadequately constrained and poorly dated. Thus, biogeographic hypotheses invoking long-distance dispersal for the distribution of many termite groups (Emerson, 1955, 1969; Bourguignon et al., 2016; Buček et al., 2022) are probably premature. It is clear that isopterans have a fossil record extending back to the mid-Mesozoic, before the major phase of Pangean breakup (McLoughlin, 2001) and, on that basis, vicariance related to continental separation may have played an important role in the distribution of termite groups through time.
CONCLUSIONS
The distinctive opal-replaced, clustered, hexagonally prismatic coprolites with blunt termini from the Griman Creek Formation at Lightning Ridge, Surat Basin, New South Wales, are tentatively assigned to kalotermitid or mastotermitid termites. Being dated to c. 100 Ma, they represent the oldest evidence of termites in Australasia, pre-dating previous body and trace fossil records by c. 40-50 million years. These coprolites demonstrate the presence of termite activity in mid-Cretaceous continental ecosystems of eastern Australia, suggesting that these organisms were already important detritivores of dead, probably coniferous wood before the diversification of angiosperms. Mineral replacement of this delicate frass cluster, presumably representing faeces backfilling a bored cavity within wood, highlights the possibility of discovering additional opalized delicate body and trace fossils at Lightning Ridge that will provide a more complete understanding of the structure, biotic interactions and energy flow within the mid-Cretaceous coastal plain ecosystems of eastern Australia. Based on the modern climatic preferences of kalotermitid and mastotermitid termites, along with co-preserved crocodilians, a relatively warm climate is invoked for the high-latitude southern Surat Basin around the Albian-Cenomanian transition. The growing number of Mesozoic body and trace fossil records from multiple continents suggests that isopterans were widely established around the world before the major phase of Pangean breakup.
ACKNOWLEDGEMENTS
This research was funded by a Swedish Research Council VR grant (number 2022-03920) to SM. Specimen AM F 128064 was donated to the Australian Museum by E. Smith. We thank the editor and reviewers for their constructive comments on the manuscript.
REFERENCES
Abensperg-Traun, M. and Steven, D. 1997. Ant- and termite-eating in Australian mammals and lizards: a comparison. Australian Journal of Ecology, 22:9-17.
https://doi.org/10.1111/j.1442-9993.1997.tb00637.x
Alonso-Zarza, A.M., Genise, J.F., Cabrera, M.C., Mangas, J., Martín-Pérez, A., Valdeolmillos, A., and Dorado-Valiño, M. 2008. Megarhizoliths in Pleistocene aeolian deposits from Gran Canaria (Spain): Ichnological and palaeoenvironmental significance. Palaeogeography, Palaeoclimatology, Palaeoecology, 265:39-51.
https://doi.org/10.1016/j.palaeo.2008.04.020
Archer, M., Flannery, T.F., Ritchie, A., and Molnar, R.E. 1985. First Mesozoic mammal from Australia - an Early Cretaceous monotreme. Nature, 318:363-366.
https://doi.org/10.1038/318363a0
Arquette, T.J. and Rodriguez, J.M. 2011. Examination of methods for Formosan subterranean termite (Isoptera: Rhinotermitidae) feces recovery. Florida Entomologist, 94:109-111.
https://doi.org/10.1653/024.094.0115
Atlas of Living Australia website, 2024. Species page:
https://bie.ala.org.au/species/
https://biodiversity.org.au/afd/taxa/bf0614d9-26df-43f0-bfe8-b6d16d5d640a
Accessed 20 April 2024.
Backhouse, J., Godfrey, I., and McLoughlin, S. 1995. Cretaceous fossil wood from Collie. Geological Survey of Western Australia Annual Review, 1994-995:51-54.
Bechly, G. 2007. Isoptera, termites, p. 249-262. In Martill, D.M., Bechly, G., and Loveridge, R.F. (eds), The Crato fossil beds of Brazil: Chap. 11.9. Cambridge University Press, New York.
Bell, P.R., Cau, A., Fanti, F., and Smith, E.T. 2016. A large clawed theropod (Dinosauria: Tetanurae) from the Lower Cretaceous of Australia and the Gondwanan origin of megaraptorid theropods. Gondwana Research, 36:473-487.
https://doi.org/10.1016/j.gr.2015.08.004
Bell, P.R., Burns, M.E., and Smith, E.T. 2018a. A probable ankylosaurian (Dinosauria, Thyreophora) from the Early Cretaceous of New South Wales, Australia. Alcheringa, 42:120-124.
https://doi.org/10.1080/03115518.2017.1384851
Bell, P.R., Herne, M.C., Brougham, T., and Smith, E.T., 2018b. Ornithopod diversity in the Griman Creek Formation (Cenomanian), New South Wales, Australia. PeerJ, 6:e6008.
https://doi.org/10.7717/peerj.6008
Bell, P.R., Fanti, F., Hart, L.J., Milan, L.A., Craven, S.J., and Smith, E. 2019. Revised geology, age, and vertebrate diversity of the dinosaur-bearing Griman Creek Formation (Cenomanian), Lightning Ridge, New South Wales, Australia. Palaeogeography, Palaeoclimatology, Palaeoecology, 514:655-671.
https://doi.org/10.1016/j.palaeo.2018.11.020
Bell, P.R., Bicknell, R.D.C., and Smith, E.T. 2020. Crayfish bio-gastroliths from eastern Australia and the middle Cretaceous distribution of Parastacidae. Geological Magazine, 157:1023-1030.
https://doi.org/10.1017/S0016756819001092
Berrell, R.W., Cavin, L., Trinajstic, K., Boisvert, C., and Smith, E.T. 2023. The first record of amiid fishes (Halecomorphi, Amiiformes, Amiidae, Calamopleurini?) from Eastern Gondwana. Cretaceous Research, 148:105538.
https://doi.org/10.1016/j.cretres.2023.105538
Bertling, M., Buatois, L.A., Knaust, D., Laing, B., Mángano, M.G., Meyer, N., Mikuláš, R., Minter, N.J., Neumann, C., Rindsberg, A.K., and Uchman, A. 2022. Names for trace fossils 2.0: theory and practice in ichnotaxonomy. Lethaia, 55:1-19.
https://doi.org/10.18261/let.55.3.3
Bignell, D.E. 2006. Termites as soil engineers and soil processors, p. 183-220. In König, H. and Varma, A. (eds.), Intestinal Microoganisms of Termites and Other Invertebrates. Springer, Berlin.
https://doi.org/10.1007/3-540-28185-1_8
Birch, S.A., Smith, E.T., and Bell, P.R. 2019. New theropod (Tetanurae: Avetheropoda) material from the ‘mid’-Cretaceous Griman Greek Formation at Lightning Ridge, New South Wales, Australia. Royal Society Open Science, 6:180826.
https://doi.org/10.1098/rsos.180826
Birch, S.A., Smith, E.T., and Bell, P.R. 2020. Noasaurids are a component of the Australian ‘mid’-Cretaceous theropod fauna. Scientific Reports, 10:1428.
https://doi.org/10.1038/s41598-020-57667-7
Bordy, E.M., Bumby, A.J., Catuneanu, O., and Eriksson, P.G. 2004. Advanced Early Jurassic termite (Insecta: Isoptera) nests: evidence from the Clarens Formation in the Tuli Basin, southern Africa. Palaios, 19:68-78.
https://doi.org/10.1669/0883-1351(2004)019<0068:AEJTII>2.0.CO;2
Bordy, E.M., Bumby, A.J., Catuneanu, O., and Eriksson, P.G. 2009. Possible trace fossils of putative termite origin in the Lower Jurassic (Karoo Supergroup) of South Africa and Lesotho. South African Journal of Science, 105:356-362.
https://doi.org/10.4102/sajs.v105i9/10.110
Bordy, E.M., Knoll, F., and Bumby, A. 2010. New data on the palaeontology and sedimentology of the Lower Jurassic Lisbon Formation (Karoo Supergroup), Ellisras Basin, South Africa. Neues Jahrbuch fur Geologie und Palaontologie-Abhandlungen, 258:145-155.
https://doi.org/10.1127/0077-7749/2010/0091
Bottini, C. and Erba, E. 2018. Mid-Cretaceous paleoenvironmental changes in the western Tethys. Climate of the Past, 14:1147-1163.
https://doi.org/10.5194/cp-14-1147-2018
Bourguignon, T., Lo, N., Cameron, S.L., Šobotník, J., Hayashi, Y., Shigenobu, S., Watanabe, D., Roisin, Y., Miura, T., and Evans, T.A. 2014. The evolutionary history of termites as inferred from 66 mitochondrial genomes. Molecular Biology and Evolution, 32:406-421.
https://doi.org/10.1093/molbev/msu308
Bourguignon, T., Lo, N., Šobotník, J., Sillam-Dussès, D., Roisin, Y., and Evans, T.A. 2016. Oceanic dispersal, vicariance and human introduction shaped the modern distribution of the termites Reticulitermes, Heterotermes and Coptotermes . Proceedings of the Royal Society B, 283:20160179.
https://doi.org/10.1098/rspb.2016.0179
Buček, A., Wang, M., Šobotník, J., Hellemans, S., Sillam-Dussès, D., Mizumoto, N., Stiblík, P., Clitheroe, C., Lu, T., González Plaza, J.J., Mohagan, A., Rafanomezantsoa, J.J., Fisher, B., Engel, M., Roisin, Y., Evans, T., Scheffrahn, R., and Bourguignon, T. 2022. Molecular phylogeny reveals the past transoceanic voyages of drywood termites (Isoptera, Kalotermitidae). Molecular Biology and Evolution 39:msac093.
https://doi.org/10.1093/molbev/msac093
Burgener, L., Hyland, E., Reich, B.J., and Scotese, C., 2023. Cretaceous climates: Mapping paleo-Köppen climatic zones using a Bayesian statistical analysis of lithologic, paleontologic, and geochemical proxies. Palaeogeography, Palaeoclimatology, Palaeoecology, 613:111373.
https://doi.org/10.1016/j.palaeo.2022.111373
Carpenter, R.J., Holman, A.I., Abell, A.D., and Grice, K. 2016. Cretaceous fire in Australia: a review with new geochemical evidence, and relevance to the rise of the angiosperms. Australian Journal of Botany, 64:564-578.
https://doi.org/10.1071/BT16109
Charles, C., Solé, F., Rodrigues, H.G., and Viriot, L. 2013. Under pressure? Dental adaptations to termitophagy and vermivory among mammals. Evolution, 67:1792-1804.
https://doi.org/10.1111/evo.12051
Child, H.J. 1934. The internal anatomy of termites and the histology of the digestive tract, p. 58-88. In Kofoid, C.A. (ed.), Termites and Termite Control. University of California Press, Berkeley, California.
Clemens, W.A., Wilson, G.P., and Molnar, R.E. 2003. An enigmatic (synapsid?) tooth from the Early Cretaceous of New South Wales, Australia. Journal of Vertebrate Paleontology, 23:232-237.
https://doi.org/10.1671/0272-4634(2003)23[232:AESTFT]2.0.CO;2
Clement, R.A., Flores-Moreno, H., Cernusak, L.A., Cheesman, A.W., Yatsko, A.R., Allison, S.D., Eggleton, P., and Zanne, A.E. 2021. Assessing the Australian termite diversity anomaly: How habitat and rainfall affect termite assemblages. Frontiers in Ecology and Evolution, 9:657444.
https://doi.org/10.3389/fevo.2021.657444
Colin, J.-P., Néraudeau, D., Nel, A., and Perrichot, V. 2011. Termite coprolites (Insecta: Isoptera) from the Cretaceous of western France: A palaeoecological insight. Revue de micropaléontologie, 54:129-139.
https://doi.org/10.1016/j.revmic.2011.06.001
Davis, R.B., Baldauf, S.L., and Mayhew, P.J. 2009. Eusociality and the success of the termites: insights from a supertree of dictyopteran families. Journal of Evolutionary Biology, 22:1750-1761.
https://doi.org/10.1111/j.1420-9101.2009.01789.x
Department of Industry, Science and Resources, Commonwealth of Australia. 2022. Resources and energy quarterly, December 2022.
https://www.industry.gov.au/publications/resources-and-energy-quarterly-december-2022
Accessed 03 June 2024.
Dettmann, M.E., Molnar, R.E., Douglas, J.G., Burger, D., Fielding, C., Clifford, H.T., Francis, J., Jell, P., Rich, T., Wade, M., Rich, P.V., Pledge, N., Kemp, A., and Rozefelds, A. 1992. Australian Cretaceous terrestrial faunas and floras: biostratigraphic and biogeographic implications. Cretaceous Research, 13:207-262.
https://doi.org/10.1016/0195-6671(92)90001-7
Dong, C., Shi, G.-L., Wang, Z.-X., and Huang, D.-Y. 2022. Termite coprolites (Blattodea: Isoptera) from the Early Cretaceous of eastern Inner Mongolia, Northeast China. Palaeoentomology, 5:006-019.
https://doi.org/10.11646/palaeoentomology.5.1.2
Eggleton, P. 2011. An introduction to termites: biology, taxonomy and functional morphology, p.1-26. In Bignell, D.E., Roisin, Y., and Lo, N. (eds), Biology of termites: a modern synthesis. Sringer, Cham.
https://doi.org/10.1007/978-90-481-3977-4_1
Emerson, A.E. 1955. Geographical origins and dispersions of termite genera. Fieldiana (Zoology), 37:465-521.
https://doi.org/10.5962/bhl.title.2783
Emerson, A.E. 1969. A revision of the Tertiary fossil species of the Kalotermitidae (Isoptera). American Museum Novitates, 2359:1-57.
Engel, M.S. 2014. A termite (Isoptera) in Late Cretaceous amber from Vendée, northwestern France. Paleontological Contributions, 2014(10E):21-24.
https://doi.org/10.17161/PC.1808.15985
Engel, M.S., Grimaldi, D.A., and Krishna, K. 2007. Primitive termites from the Early Cretaceous of Asia (Isoptera). Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie), 371:1-32.
Engel, M.S., Grimaldi, D.A., and Krishna, K. 2009. Termites (Isoptera): Their phylogeny, classification, and rise to ecological dominance. American Museum Novitates, 3650:1-27.
Engel, M.S. and Delclòs, X. 2010. Primitive termites in Cretaceous amber from Spain and Canada (Isoptera). Journal of the Kansas Entomological Society, 83:111-128.
https://doi.org/10.2317/JKES0908.06.1
Engel, M.S., Nel, A., Azar, D., Soriano, C., Tafforeau, P., Néraudeau, D., Colin J.-P., and Perrichot, V. 2011. New, primitive termites (Isoptera) from Early Cretaceous ambers of France and Lebanon. Palaeodiversity, 4:39–49.
Engel, M.S., Barden, P., Riccio, M.L., and Grimaldi, D.A. 2016. Morphologically specialized termite castes and advanced sociality in the Early Cretaceous. Current Biology, 26:522-530.
https://doi.org/10.1016/j.cub.2015.12.061
Engel, M.S. and Joault, C. 2024. Hodotermopsid termites from the mid-Cretaceous Hkamti and Kachin ambers (Isoptera: Hodotermopsidae). Palaeoentomology, 7:80-91.
https://doi.org/10.11646/palaeoentomology.7.1.5
Flannery, T.F., McCurry, M.R., Rich, T.H., Vickers-Rich, P., Smith, E.T., and Helgen, K.M. 2024. A diverse assemblage of monotremes (Monotremata) from the Cenomanian Lightning Ridge fauna of New South Wales, Australia. Alcheringa, 48:319-337.
https://doi.org/10.1080/03115518.2024.2348753
Fletcher, T.L., Cantrill, D.J., Moss, P.T., and Salisbury, S.W. 2014. A new species of Protophyllocladoxylon from the Upper Cretaceous (Cenomanian-Turonian) portion of the Winton Formation, central-western Queensland, Australia. Review of Palaeobotany and Palynology, 208:43-49.
https://doi.org/10.1016/j.revpalbo.2014.05.004
Fletcher, T.L. and Salisbury, S.W. 2014. Probable oribatid mite (Acari: Oribatida) tunnels and faecal pellets in silicified conifer wood from the Upper Cretaceous (Cenomanian-Turonian) portion of the Winton Formation, central-western Queensland, Australia. Alcheringa, 38:541-545.
https://doi.org/10.1080/03115518.2014.912557
Fletcher, T.L., Moss, P.T., and Salisbury, S.W. 2015. Wood growth indices as climate indicators from the Upper Cretaceous (Cenomanian-Turonian) portion of the Winton Formation, Australia. Palaeogeography, Palaeoclimatology, Palaeoecology, 417:35-43.
https://doi.org/10.1016/j.palaeo.2014.10.012
Fontes, L.R. and Vulcano, M.A. 1998. Cupins fosseis do Novo Mundo. In Fontes, L.R. and Filho, E.B. (eds.), Cupins: o desafio do conhecimento: 243-295. Piracicaba, Brazil: FEALZ, 512 pp.
Forster, A., Schouten, S., Baas, M., and Sinninghe Damsté, J.S. 2007. Mid-Cretaceous (Albian-Santonian) sea surface temperature record of the tropical Atlantic Ocean. Geology, 35:919-922.
https://doi.org/10.1130/G23874A.1
Frakes, L.A. and Francis, J.E. 1990. Cretaceous palaeoclimates, p. 273-287. In Ginsburg, R.N. and Beaudoin, B. (eds.), Cretaceous Resources, Events and Rhythms. Kluwer Academic Publishers, Dordrecht.
https://doi.org/10.1007/978-94-015-6861-6_14
Francis, J.E. and Harland, B.M. 2006. Termite borings in Early Cretaceous fossil wood, Isle of Wight, UK. Cretaceous Research, 27:773-777.
https://doi.org/10.1016/j.cretres.2006.02.001
Frauenfelder, T.G., Campione, N.E., Smith, E.T., and Bell, P.R. 2021. Diversity and palaeoecology of Australia’s southern-most sauropods, Griman Creek Formation (Cenomanian), New South Wales, Australia. Lethaia, 54:354-367.
https://doi.org/10.1111/let.12407
Genise, J.F. 2017. The trace fossil record of eusociality in ants and termites, p. 285-312. In Genise, J.F. (ed.), Ichnoentomology: Insect Traces in Soils and Paleosols. Springer, Cham.
Genise, J.F., Bellosi, E.S., Melchor, R.N., and Cosarinsky, M.I. 2005. Comment: Advanced early Jurassic termite (Insecta: Isoptera) nests: evidence from the Clarens Formation in the Tuli Basin, southern Africa (Bordy et al., 2004). Palaios, 20:303-308.
https://doi.org/10.2110/palo.2004.p05-C01
Green, P.M., Hoffmann, T.J.B., and Gray, A.R.G. 1997. The Surat and Bowen Basins South-East Queensland. Queensland Department of Mines and Energy, Geological Survey Division, Brisbane, 238 pp.
Greppi, C.D., García Massini, J.L., Rombola, C.F., and Pujana, R.R. 2023. Borings and coprolites of termites in fossil woods from the Lower Cretaceous (Kachaike Formation) of Argentinean Patagonia. Review of Palaeobotany and Palynology, 308:104800.
https://doi.org/10.1016/j.revpalbo.2022.104800
Griffiths, H.M., Ashton, L.A., Evans, T.A., Parr, C.L., and Eggleton, P. 2019. Termites can decompose more than half of deadwood in tropical rainforest. Current Biology, 29:R118-R119.
https://doi.org/10.1016/j.cub.2019.01.012
Griffiths, H.M., Ashton, L.A., Parr, C.L., and Eggleton, P. 2021. The impact of invertebrate decomposers on plants and soil. New Phytologist, 231:2142-2149.
https://doi.org/10.1111/nph.17553
Grimaldi, D.A., Engel, M.S., and Krishna, K. 2008. The species of Isoptera (Insecta) from the Early Cretaceous Crato Formation: a revision. American Museum Novitates, 2008:1-30.
https://doi.org/10.1206/616.1
Hall, J.W. 1963. Megaspores and other fossils in the Dakota Formation (Cenomanian) of Iowa (U.S.A.). Pollen et Spores 5:425-443.
Hamilton-Bruce, R.J., Smith, B.J., and Gowlett-Holmes, K.L. 2002. Descriptions of a new genus and two new species of viviparid snails (Mollusca: Gastropoda: Viviparidae) from the Early Cretaceous (middle-late Albian) Griman Creek Formation of Lightning Ridge, northern New South Wales. Records of the South Australian Museum, 35:193-203.
Hamilton-Bruce, R.J., Kear, B.P., and Smith, B.J. 2004. A new non-marine Early Cretaceous gastropod species from Lightning Ridge, New South Wales. Alcheringa, 28:485-492.
https://doi.org/10.1080/03115510408619297
Hamilton-Bruce, R.J. and Kear, B.P. 2006. A new fossil non-marine snail (Gastropoda) from the Lower Cretaceous (Albian, Griman Creek Formation) of eastern Australia. Molluscan Research, 26:84-88.
https://doi.org/10.11646/mr.26.2.4
Hamilton-Bruce, R.J. and Kear, B.P. 2010. A possible succineid land snail from the Lower Cretaceous non-marine deposits of the Griman Creek Formation at Lightning Ridge, New South Wales. Alcheringa, 34:325-331.
https://doi.org/10.1080/03115511003723279
Hart, L.J., Bell, P.R., Smith, E.T., and Salisbury, S.W. 2019. Isisfordia molnari sp. nov., a new basal eusuchian from the mid-Cretaceous of Lightning Ridge, Australia. PeerJ, 7:e7166.
https://doi.org/10.7717/peerj.7166
Hasiotis, S.T. and Dubiel, R.F. 1995. Termite (Insecta: Isoptera) nest ichnofossils from the Upper Triassic Chinle Formation, petrified forest national park, Arizona. Ichnos, 4:119-130.
https://doi.org/10.1080/10420949509380119
Hellemans, S., Šobotník, J., Lepont, G., Mihaljevič, M., Roisin, Y., and Bourguignon, T. 2022. Termite dispersal is influenced by their diet. Proceedings of the Royal Society B, 289:20220246.
https://doi.org/10.1098/rspb.2022.0246
Herrmann, J. and Maas, R. 2022. Formation of sediment-hosted Opal-AG at Lightning Ridge (New SouthWales, Australia): Refining the deep-weathering model. Journal of Geology, 130:77-109.
Hocknull, S.A. 2000. Mesozoic freshwater and estuarine bivalves from Australia. Memoirs of the Queensland Museum, 45:405-426.
Huber, B.T., MacLeod, K.G., Watkins, D.K., and Coffin, M.F. 2018. The rise and fall of the Cretaceous Hot Greenhouse climate. Global and Planetary Change, 167:1-23.
https://doi.org/10.1016/j.gloplacha.2018.04.004
Jarzembowski, E. 1981. An Early Cretaceous termite from southern England (Isoptera: Hodotermitidae). Systematic Entomology, 6:91-96.
https://doi.org/10.1111/j.1365-3113.1981.tb00018.x
Jiang, Y., Deng, X., Shih, C., Zhao, Y., Ren, D., and Zhao, Z. 2024. Primitive new termites (Blattodea, Termitoidae) in Cretaceous amber from Myanmar. ZooKeys, 1197:115-126.
https://doi.org/10.3897/zookeys.1197.114452
Jones, D.T. and Eggleton, P. 2011. Global biogeography of termites: a compilation of sources, p. 477-498. In Bignell, D.E., Roisin, Y., and Lo, N. (eds.), Biology of Termites: a modern synthesis. Springer, Dordrecht.
https://doi.org/10.1007/978-90- 481-3977-4_17
Jouault, C. and Nam, G.S. 2023. A new primitive termite from the Lower Cretaceous (Albian) Jinju Formation of Korea. Historical Biology, 35:1522-1527.
https://doi.org/10.1080/08912963.2022.2098490
Jouault, C., Legendre, F., Grandcolas, P., and Nel, A. 2021. Revising dating estimates and the antiquity of eusociality in termites using the fossilized birth-death process. Systematic Entomology, 46:592-610.
https://doi.org/10.1111/syen.12477
Jouault, C., Engel, M.S., Huang, D., Berger, J., Grandcolas, P., Perkovsky, E.E., Legendre, F., and Nel, A. 2022a. Termite Valkyries: soldier-like alate termites from the Cretaceous and task specialization in the early evolution of Isoptera. Frontiers in Ecology and Evolution, 10:737367.
https://doi.org/10.3389/fevo.2022.737367
Jouault, C., Engel, M.S., Legendre, F., Huang, D., Grandcolas, P., and Nel, A. 2022b. Incrementing and clarifying the diversity and early evolution of termites (Blattodea: Isoptera). Zoological Journal of the Linnean Society, 196:608-629.
https://doi.org/10.1093/zoolinnean/zlac064
Kaulfuss, U., Harris, A.C., and Lee, D.E., 2010. A new fossil termite (Isoptera, Stolotermitidae, Stolotermes ) from the Early Miocene of Otago, New Zealand. Acta Geologica Sinica, 84:705-709.
https://doi.org/10.1111/j.1755-6724.2010.00275.x
Kear, B.P. 2006. Marine reptiles from the Lower Cretaceous of South Australia: Elements of a high-latitude cold-water assemblage. The Palaeontological Association, 49:837-856.
https://doi.org/10.1111/j.1475-4983.2006.00569.x
Kear, B.P. and Godthelp, H. 2008. Inferred vertebrate bite marks on an Early Cretaceous unionoid bivalve from Lightning Ridge, New South Wales, Australia. Alcheringa, 32:65-71.
https://doi.org/10.1080/03115510701757498
Kemp, A. and Molnar, R.E. 1981. Neoceratodus forsteri from the Lower Cretaceous of New South Wales, Australia. Journal of Paleontology, 55:211-217.
Kemp, A. and Berrell, R. 2020. A new species of fossil lungfish (Osteichthyes: Dipnoi) from the Cretaceous of Australia. Journal of Vertebrate Paleontology, 40:e1822369.
https://doi.org/10.1080/02724634.2020.1822369
Kitchener, J.L., Campione, N.E., Smith, E.T., and Bell, P.R. 2019. High-latitude neonate and perinate ornithopods from the mid-Cretaceous of southeastern Australia. Scientific Reports, 9:19600.
https://doi.org/10.1038/s41598-019-56069-8
Kovach, W.L. and Dilcher D.L. 1988. Megaspores and other dispersed plant remains from the Dakota Formation (Cenomanian) of Kansas, USA. Palynology, 12:89-119.
https://doi.org/10.1080/01916122.1988.9989338
Krishna, K. 1990. Isoptera, p. 76-81. In Grimaldi, D.A. (ed.), Insects from the Santana Formation, Lower Cretaceous, of Brazil. Bulletin of the American Museum of Natural History 195.
Krishna, K. and Grimaldi, D.A. 2000. A new subfamily, genus, and species of termite (Isoptera) from New Jersey Cretaceous amber. Studies on Fossils in Amber, with Particular Reference to the Cretaceous of New Jersey. Backhuys Publishers, Leiden, The Netherlands, 133-140.
Krishna, K. and Grimaldi, D.A. 2003. The first Cretaceous Rhinotermitidae (Isoptera): a new species, genus, and subfamily in Burmese amber. American Museum Novitates, 2003:1-10.
Lacasa-Ruiz, A. and Martínez-Delclòs, X. 1986. Meiatermes: Nuevo género fosil de insecto isóptero, Hodotermitidae, de las calizas Neocomienses del Monsec, Provincía de Lérida, España. 1986, 65 pp.
Lance, J.F. 1946. Fossil arthropods of California. 9. Evidence of termites in the Pleistocene asphalt of Carpintera, California. Southern California Academy of Sciences Bulletin, 45:21-27.
https://doi.org/10.3160/0038-3872-45.1.21
Langford, R.P., Wilford, G.E., Truswell, E.M., and Isern, A.R. 1995. Palaeogeographic Atlas of Australia, Vol. 10 - Cainozoic. Australian Geological Survey Organisation, Canberra.
Legendre, F., Whiting, M.F., Bordereau, C., Cancello, E.M., Evans, T.A., and Grandcolas, P. 2008. The phylogeny of termites (Dictyoptera: Isoptera) based on mitochondrial and nuclear genes: implications for the evolution of the worker and pseudergate castes, and foraging behaviors. Molecular Phylogenetics and Evolution, 48:615-627.
https://doi.org/10.1016/j.ympev.2008.04.017
Legendre, F., Nel, A., Svenson, G.J., Robillard, T., Pellens, R., and Grandcolas, P. 2015. Phylogeny of Dictyoptera: dating the origin of cockroaches, praying mantises and termites with molecular data and controlled fossil evidence. Plos one, 10(7):e0130127.
Lewis, V.R., Sutherland, A.M., and Haverty, M.I. 2014. Drywood termites. University of California, Agriculture and Natural Resources, Pest Notes Publication, 7440:1-5.
Light, S.F. 1934. A world view of termites, p. 117-125. In Kofoili, C.A. (ed.), Termites and Termite Control. University of California Press, Berkeley, California.
Lo, N. and Eggleton, P. 2011. Termite phylogenetics and co-claenesis with symbionts, p. 27-50. In Bignell, D.E., Roisin, Y., and Lo, N. (eds.), Biology of Termites: A Modern Synthesis. Springer, Dordrecht.
https://doi.org/10.1007/978-90-481-3977-4_2
Luo, Z.X. and Wible, J.R. 2005. A Late Jurassic digging mammal and early mammalian diversification. Science, 308:103-107.
https://doi.org/10.1126/science.1108875
Martínez-Delclòs, X. and Martinell, J. 1995. The oldest known record of social insects. Journal of Paleontology, 69:594-599.
https://doi.org/10.1017/S0022336000034983
Martins-Neto, R.G., Ribeiro-Júnior, C., and Prezoto, F. 2006. New fossils (Isoptera: Hodotermitidae), from the Santana Formation (Lower Cretaceous, Araripe Basin, Northeast Brazil), with descriptions of new taxa including a new subfamily. Sociobiology, 47:125-134.
McCurry, M.R., Cantrill, D.J., Smith, P.M., Beattie, R., Dettmann, M., Baranov, V., Magee, C., Nguyen, J.M.T., Forster, M.A., Hinde, J., Pogson, R., Wang, H., Marjo, C.E., Vasconcelos, P., and Frese, M. 2022. A Lagerstätte from Australia provides insight into the nature of Miocene mesic ecosystems. Science Advances, 8:eabm1406.
https://doi.org/10.1126/sciadv.abm1406
McLoughlin, S. 1996. Early Cretaceous macrofloras of Western Australia. Records of the Western Australian Museum, 18:19-65.
McLoughlin, S. 2001. The breakup history of Gondwana and its impact on pre-Cenozoic floristic provincialism. Australian Journal of Botany, 49:271-300.
https://doi.org/10.1071/BT00023
McLoughlin, S. and McNamara, K., 2001. Ancient Floras of Western Australia. Department of Earth and Planetary Sciences, Western Australian Museum, Perth.
McLoughlin, S., Drinnan, A.N., and Rozefelds, A.C. 1995a. The Cenomanian flora from the Winton Formation, Eromanga Basin, Queensland, Australia. Memoirs of the Queensland Museum, 38:273-313.
McLoughlin, S., Haig. D.W., Backhouse, J., Holmes, M.A., Ellis, G., Long, J.A., and McNamara, K.J. 1995b. Oldest Cretaceous sequence, Giralia Anticline, Carnarvon Basin, Western Australia: late Hauterivian-Barremian. AGSO Journal of Australian Geology and Geophysics, 15:445-468.
McLoughlin, S., Tosolini, A.-M.P., Nagalingum, N.S., and Drinnan, A.N. 2002. Early Cretaceous (Neocomian) flora and Fauna of the lower Strzelecki Group, Gippsland Basin, Victoria. Association of Australasian Palaeontologists Memoir, 26:1-144.
McLoughlin, S., Pott, C., and Elliott, D. 2010. The Winton Formation flora (Albian-Cenomanian, Eromanga Basin): implications for vascular plant diversification and decline in the Australian Cretaceous. Alcheringa, 34:303-323.
https://doi.org/10.1080/03115511003669944
Meakin, S. 2011. Geodiversity of the Lightning Ridge area and implications for Geotourism. Proceedings of the Linnean Society of New South Wales, 132:71-82.
Miller, L.R. 1989. Sub-fossil termite mounds in the Simpson Desert. Northern Territory Naturalist, 11:27-30.
Molnar, R.E. 1999. Avian tibiotarsi from the Early Cretaceous of Lightning Ridge, New South Wales. In Tomida, Y., Rich, P.V., and Vickers-Rich, P. (eds.), Proceedings of the Second Gondwanan Dinosaur Symposium. National Science Museum Monographs 15. pp. 197-209.
Molnar, R.E. and Galton, P.M. 1986. Hypsilophodontid dinosaurs from Lightning Ridge, New South Wales, Australia. Geobios, 19:231-243.
https://doi.org/10.1016/S0016-6995(86)80046-8
Moore, M., 2002. Geophysical Ground Surveys Over Lightning Ridge Opal Prospects. Geological Survey Report, GS2002/442:1-40.
Moreau, J.D., Trincal, V., Nel, A., Simon-Coinçon, R., Sallé, V., Couls, M.L., Néraudeau, D., and Fernandez, V. 2019. Hidden termite coprolites revealed by synchrotron microtomography inside Eocene‒Oligocene filled wood-borings from the Malzieu Basin, Lozère, southern France. Lethaia, 53:106-117.
https://doi.org/10.1111/let.12344
Mory, A.J. and Hocking, R.M. 2017. Geology of the Kennedy Range Area. Geological Survey of Western Australia, Perth.
Mustoe, G.E. and Smith, E.T., 2023. Timing of opalization at Lightning Ridge, Australia: New evidence from opalized fossils. Minerals, 13:1471.
https://doi.org/10.3390/min13121471
Nauer, P.A., Hutley, L.B., and Arndt, S.K., 2018. Termite mounds mitigate half of termite methane emissions. Proceedings of the National Academy of Sciences United States of America, 115:13306-13311.
https://doi.org/10.1073/pnas.1809790115
Newton, R.B. 1915. On some molluscan remains from the opal deposits (Upper Cretaceous) of New South Wales. Proceedings of the Malacological Society, London, 10:217-235.
https://doi.org/10.1093/oxfordjournals.mollus.a063569
Noirot, C. 1970. The nests of termites, p. 73-125. In Krishna, K. and Weesner, F.M. (eds.), Biology of Termites, 2. Academic Press, New York.
Payenberg, T.H.D. and Reilly, M.R.W. 2004. Lower Balonne Airborne Geophysical Project. Core descriptions of ten conventional cores from the lower Balonne area, southern Queensland, Australia. CRC LEME Open File Report, 166:1-15. Cooperative Research Centre for Landscape Environments and Mineral Exploration, Perth, Western Australia.
https://crcleme.org.au/Pubs/OPEN%20FILE%20REPORTS/OFR%20161-162-163-164-166-167/OFR166.pdf
Philippe, M., McLoughlin, S., Strullu-Derrien, C., Bamford, M., Kiel, S., Nel, A., and Thévenard, F. 2022. Life in the woods: taphonomic evolution of a diverse saproxylic community within fossil woods from Upper Cretaceous submarine mass flow deposits (Mzamba Formation, southeast Africa). Gondwana Research, 109:113-133.
https://doi.org/10.1016/j.gr.2022.04.008
Philippe, M., Bamford, M., McLoughlin, S., Da Rosa Alves, L.S., Falcon-Lang, H.J., Gnaedinger, S., Ottone, E., Pole, M., Rajanikanth, A., Shoemaker, R.E., Torres, T., and Zamuner, A. 2004. Biogeographic analysis of Jurassic-Early Cretaceous wood assemblages from Gondwana. Review of Palaeobotany and Palynology, 129:141-173.
https://doi.org/10.1016/j.revpalbo.2004.01.005
Pires, E.F. and Sommer, M.G. 2009. Plant-arthropod interaction in the Early Cretaceous (Berriasian) of the Araripe Basin, Brazil. Journal of South American Earth Sciences, 27:50-59.
https://doi.org/10.1016/j.jsames.2008.09.004
Redford, K.H. 1987. Ants and termites as food, p. 349-399. In Genoways, H.H. (ed.), Current Mammalogy. Springer, Boston, MA.
https://doi.org/10.1007/978-1-4757-9909-5_9
Ren, D. 1995. Isoptera Comstock, 1895. pp. 56-61. In Ren, D., Lu, L., Guo, Z., and Ji, S. (eds.), Faunae and Stratigraphy of Jurassic-Cretaceous in Beijing and the Adjacent Areas. Seismic Publishing House, Beijing.
Riek, E. 1952. Fossil insects from the Tertiary sediments at Dinmore, Queensland. Papers Department of Geology, University of Queensland, 4:17-22.
Rogers, A.F. 1928. Natural history of the silica minerals. American Mineralogist, 13:73-87.
Rogers, A.F. 1938. Fossil termite pellets in opalised wood from Santa Barbara County, California. American Journal of Science, 36:389-392.
https://doi.org/10.2475/ajs.s5-36.215.389
Rohr, O.M., Boucot, A.J., Miller, J., and Abbott, M. 1986. Oldest termite nest from the Upper Cretaceous of west Texas. Geology, 14:87-88.
https://doi.org/10.1130/0091-7613(1986)14<87:OTNFTU>2.0.CO;2
Rozefelds, A.C. and De Baar, M. 1991. Silicified Kalotermitidae (Isoptera) frass in conifer wood from a mid-Tertiary rainforest in central Queensland, Australia. Lethaia, 24:439-442.
https://doi.org/10.1111/j.1502-3931.1991.tb01498.x
Sanderson, M.G. 1996. Biomass of termites and their emissions of methane and carbon dioxide: A global database. Global Biogeochemical Cycles, 10:543-557.
https://doi.org/10.1029/96GB01893
Sánchez-García, A., Peñalver, E., Delclòs, X., and Engel, M. S. 2020. Early Cretaceous termites in amber from northern Spain (Isoptera). Cretaceous Research, 110:104385.
Schlüter, T. 1989. Neue Daten über harzkonservierte Arthropoden aus dem Cenomanium NW-Frankreichs. Documenta naturae, 56:59-70.
Sheppe, W. 1970. Invertebrate predation on termites of the African savanna. Insectes Sociaux, 17:205-218.
https://doi.org/10.1007/BF02226194
Simpson, E.S. 1912. Unusual types of petrifaction from Dandarragan. Journal of the Natural History Scientific Society of Western Australia, 4:33-37.
Smith, E.T. 2010. Early Cretaceous chelids from Lightning Ridge, New South Wales. Alcheringa, 34:375-384.
https://doi.org/10.1080/03115518.2010.488117
Smith, E.T. and Kear, B.P., 2013. Spoochelys ormondea gen. et sp. nov., an archaic meiolaniid-like turtle from the Early Cretaceous of Lightning Ridge, Australia, pp. 121-146. In Brinkman, D.B., Holroyd, P.A., Gardner, J.D. (eds.), Morphology and Evolution of Turtles, Vertebrate Paleobiology and Paleoanthropology. Springer Netherlands, Dordrecht.
https://doi.org/10.1007/978-94-007-4309-0_9
Smith, E. and Smith, R. 1999. Black Opal Fossils of Lightning Ridge. Kangaroo Press, Sydney.
Sutherland, J.I. 2003. Miocene petrified wood and associated borings and termite faecal pellets from Hukatere Peninsula, Kaipara Harbour, North Auckland, New Zealand. Journal of the Royal Society of New Zealand, 33:395-414.
https://doi.org/10.1080/03014223.2003.9517736
Tosolini, A.-M.P., Korasidis, V.A., Wagstaff, B.E., Cantrill, D.J., Gallagher, S.J., and Norvick, M.S. 2018. Palaeoenvironments and palaeocommunities from Lower Cretaceous high-latitude sites, Otway Basin, southeastern Australia. Palaeogeography, Palaeoclimatology, Palaeoecology, 496:62-84.
https://doi.org/10.1016/j.palaeo.2018.01.017
Vangerow, E.F. 1954. Megasporen und andere pflanzliche Mikrofossilien aus der Aachener Kreide. Palaeontographica B, 96:24-38.
Vršanský, P. and Aristov, D. 2014. Termites (Isoptera) from the Jurassic/Cretaceous boundary: evidence for the longevity of their earliest genera. European Journal of Entomology, 111:137-141.
https://doi.org/10.14411/eje.2014.014
Ware, J.L., Grimaldi, D.A., and Engel, M.S. 2010. The effects of fossil placement and calibration on divergence times and rates: an example from the termites (Insecta: Isoptera). Arthropod Structure & Development, 39:204-219.
https://doi.org/10.1016/j.asd.2009.11.003
Watson, J.A.L. and Abbey, H.M. 1993. Atlas of Australian Termites. CSIRO, Melbourne.
Weidner, H. 1956. Kotballen von Termiten im Bernstein. Veröffentlichungen aus dem Übersee-Museum Bremen, Reihe A, Naturwissenschaften, A2:363-364.
White, M.E. 1961. Report on 1960 collections of Mesozoic plant fossils from the Northern Territory. Bureau of Mineral Resources, Geology and Geophysics, Australia, Record, 1961/146:1-26.
Zhao, Z., Yin, X., Shih, C., Gao, T., and Ren, D. 2020. Termite colonies from mid-Cretaceous Myanmar demonstrate their early eusocial lifestyle in damp wood. National Science Review, 7:381-390.
https://doi.org/10.1093/nsr/nwz141
