| |
ISOCRINID CRAWLING
Finger-Tip Pull
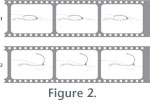 While indirect evidence of isocrinid motility had been gathered through submersible
observations since at least the mid-1980s (Messing
1985; Messing
et al. 1988; Baumiller
et al. 1991), direct evidence of crawling has until now consisted
of one in situ report (Messing et al. 1988) and two sets
of laboratory observations (Baumiller
et al. 1991; Birenheide
and Motokawa 1994). These direct observations revealed that
during crawling the crinoid is prone on the substrate with most
of the stalk horizontal. Only the proximal portion bent sharply
away from the substrate, such that the oral-aboral axis of the calyx
orients sub-vertically with the oral surface pointing up (Figure
2.1). The arms, arranged radially around the calyx, have their long
axes sub-parallel to the substrate with ambulacra facing up. Crawling
involves a repeating sequence of movements by the leading arms,
that is, those directed away from the stalk, consisting of a power
and a recovery stroke by each of the leading arms: 1) with the distal
tip strongly flexed and pressing against the substrate, the rest
of the arm curls slightly aborally, pulling the animal forward (power
phase); 2) the arm tip lifts off the substrate and the arm straightens
by extending orally (recovery phase); and 3) the tip lowers against
the substrate and the sequence is repeated. This sequence is somewhat
analogous to crawling using one's fingertips, and we refer to it
as the "finger-tip pull" mode of crawling. The movement
of the leading arms is out of phase; while some arms flex and pull,
others straighten. The non-leading arms, that is, those on the side
and closer to the stalk, are slightly flexed aborally and, together
with the stalk and cirri, are pulled passively by the leading arms.
In some instances, these arms have been observed to push the individual
forward by anchoring the tip in the substrate, and extending orally
(Birenheide
and Motokawa 1994). While indirect evidence of isocrinid motility had been gathered through submersible
observations since at least the mid-1980s (Messing
1985; Messing
et al. 1988; Baumiller
et al. 1991), direct evidence of crawling has until now consisted
of one in situ report (Messing et al. 1988) and two sets
of laboratory observations (Baumiller
et al. 1991; Birenheide
and Motokawa 1994). These direct observations revealed that
during crawling the crinoid is prone on the substrate with most
of the stalk horizontal. Only the proximal portion bent sharply
away from the substrate, such that the oral-aboral axis of the calyx
orients sub-vertically with the oral surface pointing up (Figure
2.1). The arms, arranged radially around the calyx, have their long
axes sub-parallel to the substrate with ambulacra facing up. Crawling
involves a repeating sequence of movements by the leading arms,
that is, those directed away from the stalk, consisting of a power
and a recovery stroke by each of the leading arms: 1) with the distal
tip strongly flexed and pressing against the substrate, the rest
of the arm curls slightly aborally, pulling the animal forward (power
phase); 2) the arm tip lifts off the substrate and the arm straightens
by extending orally (recovery phase); and 3) the tip lowers against
the substrate and the sequence is repeated. This sequence is somewhat
analogous to crawling using one's fingertips, and we refer to it
as the "finger-tip pull" mode of crawling. The movement
of the leading arms is out of phase; while some arms flex and pull,
others straighten. The non-leading arms, that is, those on the side
and closer to the stalk, are slightly flexed aborally and, together
with the stalk and cirri, are pulled passively by the leading arms.
In some instances, these arms have been observed to push the individual
forward by anchoring the tip in the substrate, and extending orally
(Birenheide
and Motokawa 1994).
Elbow-Crawl
 While finger-tip pull may characterize isocrinid locomotion under some circumstances,
we recorded a very different crawling movement by a specimen of
Neocrinus decorus on a ~5 minute video sequence during dive
JSL 3479 (Figure 3). This behavior may be described as "elbow-crawl" (Figure 2.2). With the crinoid prone, the entire stalk
is nearly straight, such that the stalk and the oral-aboral axis
of the calyx are nearly parallel to the substrate. Strong aboral
flexure curves the radiating arms so that their tips point toward
the stalk. Those arms adjacent to the substrate undergo a sequence
of power and recovery strokes, while the rest of the arms, elevated
above the substrate, remain strongly flexed aborally, but virtually
static. As in finger-tip pull crawling, the power stroke consists
of aboral flexure of the arm, while the recovery involves oral straightening.
However, whereas in finger-tip pull, only the flexed tip of the
nearly straight arm presses against the substrate, and the ambulacral
pinnule-bearing surface of the arm orients upward, in elbow-crawl,
the arms remain flexed aborally throughout the stroke cycle so that
it is the pinnule-covered oral side of the middle third to distal
half of the arm that creates traction with the substrate. In the
recovery stroke, the middle of the arm uncurls slightly while the
base of the arm flexes toward the mouth, bringing the entire curved
middle and distal portions of the arm up and forward one or a few
centimeters. In the power stroke, the arm base bends away from the
mouth, pushing the more strongly flexed arm down and backward against
the substrate. The pinnules may also aid crawling via a ratchet-like
mechanism. During the recovery stroke, the pinnules are lifted off
the substrate and spread out at an angle from the arm axis. In the
power stroke, as they touch the substrate, they flex at an acute
angle opposite the direction of motion, perhaps functioning like
oars. While finger-tip pull may characterize isocrinid locomotion under some circumstances,
we recorded a very different crawling movement by a specimen of
Neocrinus decorus on a ~5 minute video sequence during dive
JSL 3479 (Figure 3). This behavior may be described as "elbow-crawl" (Figure 2.2). With the crinoid prone, the entire stalk
is nearly straight, such that the stalk and the oral-aboral axis
of the calyx are nearly parallel to the substrate. Strong aboral
flexure curves the radiating arms so that their tips point toward
the stalk. Those arms adjacent to the substrate undergo a sequence
of power and recovery strokes, while the rest of the arms, elevated
above the substrate, remain strongly flexed aborally, but virtually
static. As in finger-tip pull crawling, the power stroke consists
of aboral flexure of the arm, while the recovery involves oral straightening.
However, whereas in finger-tip pull, only the flexed tip of the
nearly straight arm presses against the substrate, and the ambulacral
pinnule-bearing surface of the arm orients upward, in elbow-crawl,
the arms remain flexed aborally throughout the stroke cycle so that
it is the pinnule-covered oral side of the middle third to distal
half of the arm that creates traction with the substrate. In the
recovery stroke, the middle of the arm uncurls slightly while the
base of the arm flexes toward the mouth, bringing the entire curved
middle and distal portions of the arm up and forward one or a few
centimeters. In the power stroke, the arm base bends away from the
mouth, pushing the more strongly flexed arm down and backward against
the substrate. The pinnules may also aid crawling via a ratchet-like
mechanism. During the recovery stroke, the pinnules are lifted off
the substrate and spread out at an angle from the arm axis. In the
power stroke, as they touch the substrate, they flex at an acute
angle opposite the direction of motion, perhaps functioning like
oars.
In both modes of crawling, there is no support for the claim that crinoids "'walk
about' on [cirri]" (Clarkson
1998, p. 297). However, as has been previously noted (Baumiller
et al. 1991), there is incontrovertible evidence for some motility
of cirri during locomotion, which suggests that these appendages
might be involved in locomotion, perhaps as ratchets or hooks.
Speeds
While isocrinids observed in the laboratory (Baumiller
et al. 1991; Birenheide
and Motokawa 1994) covered distances up to a meter using the
finger-tip pull mode, their speeds were so slow that movement was
nearly undetectable with the naked eye; for example, Birenheide
and Motokawa (1994) reported speeds of 0.5 m h-1
(~0.1 mm sec-1). In this context, the most striking feature
of the elbow-crawl mode recorded in situ is the much higher
speed; Neocrinus decorus covered more than 3 m in just under
5 min, corresponding to an average speed of 36 m h-1
(~10 mm sec-1). In fact, the 5 min of recorded behavior
consisted of spurts of even more rapid movement (~30 mm sec-1)
interspersed with intervals of slower crawling or no movement. In this context, the most striking feature
of the elbow-crawl mode recorded in situ is the much higher
speed; Neocrinus decorus covered more than 3 m in just under
5 min, corresponding to an average speed of 36 m h-1
(~10 mm sec-1). In fact, the 5 min of recorded behavior
consisted of spurts of even more rapid movement (~30 mm sec-1)
interspersed with intervals of slower crawling or no movement.
Traces
The arms and stalk of crawling crinoids interact with the substrate, and this
activity ought to produce traces. Previously, Messing
et al. (1988) recognized what they considered to be two types
of traces left by a crawling isocrinid: "a drag mark over 1
m long..." left by the stalk (p. 481); and
"short radiating scratch marks" on the substrate around
the crown and on the sediment surface behind an isocrinid made by
the arms involved in crawling. Unfortunately, no photos of either
trace are available.
 Our observations provide further proof for
at least one such trace: in the video footage, grains of sediment
are displaced as the stalk is pulled behind the crawling N. decorus
producing a drag mark (Figure 4). While the low angle at which the
video footage was shot makes it difficult to recognize any of the
fine traces that would be produced by the power strokes of the arms,
experiments with comatulids crawling on fine-grained substrate reveal
the types of traces that such behavior is likely to produce (Figure
5). Our observations provide further proof for
at least one such trace: in the video footage, grains of sediment
are displaced as the stalk is pulled behind the crawling N. decorus
producing a drag mark (Figure 4). While the low angle at which the
video footage was shot makes it difficult to recognize any of the
fine traces that would be produced by the power strokes of the arms,
experiments with comatulids crawling on fine-grained substrate reveal
the types of traces that such behavior is likely to produce (Figure
5).
|