|
||||
DISCUSSIONThe crawling behavior of isocrinids, especially the fast mode of elbow-crawl reported for the first time, has implications for several aspects of crinoid biology and paleobiology. First, we need to ask whether a biomechanical basis exists for the apparent differences in speed between finger-tip pull and elbow-crawl. Second, in light of the much more rapid speeds of crawling of isocrinids, we need to reconsider the biological role of crawling. Finally, from paleontological and evolutionary perspectives, crawling abilities of isocrinids lead us to ask how one might recognize crawling abilities among extinct crinoids and what is the history of this behavior? Effect of Crawling Mode on Speed: Biomechanical ConsiderationsAlthough N. decorus observed in situ was moving down a slight slope and laboratory specimens used the finger-tip pull to crawl along a flat-bottomed aquarium, this alone is unlikely to account for the >100-fold difference in speed. More probably, the differences between the two modes of crawling account for the dramatically different speeds.
Of course, other differences between elbow-crawling and finger-tip pulling may contribute to the difference in speed. For example, the above-mentioned use of the pinnule "ratchet" in the former case may be an effective strategy of increasing traction with the substrate and preventing slippage of the arm during the power stroke. This could have an impact on speed. A similar, but reversed, action occurs in swimming comatulids, in which the pinnules lie against the arm to provide much less resistance on the upward recovery stroke, and spread out at almost right angles to the arm axis on the downward power stroke, offering much greater surface area to act against the water. Biological Role of CrawlingRegardless of how the differences in speeds between the elbow-crawl and the finger-pull modes of locomotion are achieved, speeds of 10-30 mm sec-1 allow us to consider very different scenarios for its biological role than speeds of 0.1 mm sec-1. For example, the suggestion that crawling may be stimulated by unfavorable micro-environmental conditions, such as inappropriate current velocities or microturbulence (Messing et al. 1988; Baumiller et al. 1991; Birenheide and Motokawa 1994), seems reasonable for both crawling speeds, as it would allow the animal to reach an environment with even a slightly different flow regime. Rapid crawling could also allow isocrinids to respond to disturbance by another organism. For example, Messing et al. (1988) reported an interaction between the isocrinid, Cenocrinus asterius, and a large, cidaroid echinoid, Calocidaris micans, in the vicinity of Georgetown, Grand Cayman. They inferred that disturbance of the crinoid's filtration fan by the echinoid may have induced crawling. More recent observations (Baumiller et al. 2000) and analyses of gut contents of cidaroids lead us to conclude that the interaction may involve cidaroid feeding on live isocrinids. In this instance, an isocrinid crawling at 0.1 mm sec-1 could not escape a cidaroid, whereas one crawling at 10-30 mm sec-1 could. In fact, we have argued (Baumiller et al. 2000) that the active stalk shedding (autotomy) and crawling are part of a "lizard tail" strategy used by isocrinids to escape from benthic enemies, such as cidaroids. Paleobiological Implications of CrawlingSince it is now established that some extant stalked crinoids, the isocrinids, can locomote under their own power and since its role in allowing the animal to respond to physical and biological cues by relocating is likely to be ecologically important, it is worth considering the history of crawling and its evolutionary implications. In reconstructing behavior of extinct organisms, one often relies on functional morphology (Plotnick and Baumiller 2000), and as a start, we have identified several features among stalked crinoids that are necessary, although not sufficient, for crawling. Among these are ability of re-attaching the stalk to the substrate, and arms that are both flexible and strong enough to generate the power/recovery stroke to pull the animal along the bottom. Both features characterize the post-Paleozoic holocrinids and isocrinids (Simms 1999), and some of the taxa generally thought to be closely allied or ancestral to the post-Paleozoic articulates, that is, the Late Paleozoic advanced cladids (Simms and Sevastopulo 1993). Stalked crinoids belonging to these clades possess a stalk that bears cirri with a transverse ridge and a terminal claw-like cirral. With the transverse ridge acting as a fulcrum, such cirri are capable of rapid oral-aboral flexure and of acting as effective anchoring appendages for re-attachment; stalked crinoids lacking cirri or those bearing cirri with multiradiate articula lacked the ability to re-attach. It is also among these clades that one finds well-developed muscular arm articulations (Simms and Sevastopulo 1993). Such brachial articulations are characterized by a central fulcrum, which in extant isocrinids divides the facet into an oral (ambulacral) side bearing both muscles and ligaments and an entirely ligamentary aboral side. Thus oral-aboral flexure of the arms is a consequence of the antagonistic action across this fulcrum. While it is difficult to prove whether extinct crinoids with such arm articulations possessed similar soft tissues (Lane and Macurda 1975; Ausich 1977; Ausich and Baumiller 1993), the recently discovered contractile properties of crinoid ligaments (Motokawa et al. 2004) suggest that such fulcrum-bearing facets, even if connected only by ligaments, could undergo active oral-aboral flexure to generate the power and recovery strokes during crawling, although undoubtedly at much lower speeds than if muscles were present. What is clear, however, is that a well-developed brachial fulcrum necessary for crawling characterized a number of Paleozoic lineages closely related to the post-Paleozoic articulates (Webster 2003). Although trace fossil data, such as the drag marks or scratch marks discussed above, could provide more direct evidence of crinoid crawling, such trace fossils have yet to be recognized. Taphonomic information such as death posture may provide evidence of crawling, but to date it has only been employed to demonstrate crawling in Jurassic comatulids and stalked isocrinids (Baumiller et al. 2004, in press).
|
||||
|
|
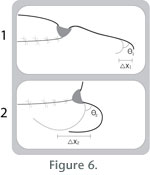
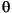 1
on a distal articulation during its power stroke produces horizontal
displacement, X1, of the distal tip which is in contact with the
substrate.
1
on a distal articulation during its power stroke produces horizontal
displacement, X1, of the distal tip which is in contact with the
substrate. 