|
|
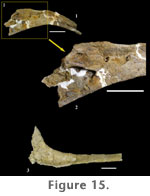
DESCRIPTIONSkull
Axial SkeletonAll of the vertebrae found at Jones Ranch appear to pertain to adult individuals, as neural arches (where preserved) are completely fused to their centra. Presacral vertebrae of P. jonesi are strongly opisthocoelous, lack bifid neural spines, and exhibit an extensive network of vertebral laminae. Herein, I refer to these laminae using the nomenclature of Wilson (1999). Cervical Vertebrae. Portions of at least nine cervical vertebrae have been recovered from the Jones Ranch quarry thus far, four of which are almost complete. Bone associations in the quarry suggest that all but two of these cervical vertebrae, FWMSH 93B-10-8 and FWMSH 93B-10-19, pertain to a single individual. Cervical vertebrae FWMSH 93B-10-32, FWMSH 93B-10-28, FWMSH 93B-10-19, FWMSH 93B-10-29, and FWMSH 93B-10-30 most likely represent vertebrae C3-C9 of the cervical column, based on comparison of the centrum length ratio between the third and fourth cervical vertebrae with that of Brachiosaurus brancai, the position and height of neural spines, and position relative to one another in the quarry. Most of the Jones Ranch cervical vertebrae have experienced some deformation related to crushing. All of the cervical vertebrae have long centra, as exhibited by high length-to-caudal height ratios, expressed as an elongation index [EI] (sensu Wilson and Sereno 1998; Wedel et al. 2000b; contra Upchurch [1998], who uses length/caudal centrum width to represent the EI). EI values for Jones Ranch cervical vertebrae range from 4.7 in C3 to 7.3 in C4 of the same individual (Table 2).
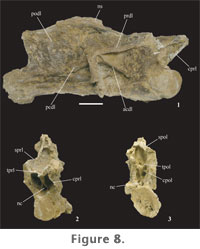
Neural arches span nearly the entire length of the centrum in the cervical series. The neural arch is tall in mid-cervical vertebrae. Prezygapophyses extend beyond the cranial condyle of the centrum. Postzygapophyses are weakly developed and are positioned cranial to the caudal margin of the centrum. Diapophyses are lightly built in cranial cervical vertebrae but become more expansive in the middle and caudal cervical vertebrae. Only cervical vertebrae six (FWMSH 93B-10-28) and eight (FWMSH 93B-10-29) preserve most of the neural spine (Figure 8,
Figure 9.1).
Long, paired spinoprezygapophyseal laminae (sprl) (already defined) originate on the dorsal surface of the prezygapophyses and terminate near the craniodorsal margin of the neural spine (Figure 9). The sprl increases in length and becomes more prominent along the column as the neural spine migrates caudally and increases in height. In addition to broad centroprezygapophyseal laminae (cprl), the prezygapophyses are linked by paired intraprezygapophyseal laminae (tprl)(already defined). The tprl's meet medially where they are joined by a vertical lamina that divides ventrally and connects to the top of the centrum, outlining the neural canal. This accessory lamina creates two bilaterally symmetrical fossae just above the neural canal (Figure 8.2). A similar arrangement of laminae and fossae can be seen in some cervical vertebrae of Brachiosaurus brancai. The spinopostzygapophyseal lamina (spol) is paired, connecting the postzygapophyses to the caudal aspect of the neural spine. Intrapostzygapophyseal laminae (tpol) (already defined) traverse between the medial surface of the postzygapophyses and the midline of vertebrae where they meet above the neural canal. The tpol's and spol's outline a deep fossa behind the neural spine. A median strut runs between the tpol and the base of the neurocentral junction. Two fossae on either side of this lamina, in conjunction with the neural canal, form a tri-radiate pattern, similar to that on the cranial surface of the vertebrae (Figure 8.3). These fossae are bounded laterally by short centropostzygapophyseal laminae (cpol). In contrast to most somphospondylians, the diapophyseal laminae are reasonably well developed in cervical vertebrae of Paluxysaurus jonesi. Four laminae stem from the diapophyses. The posterior centrodiapophyseal (pcdl) and postzygodiapophyseal (podl) laminae branch at a shallow angle away from the diapophysis caudoventrally and caudodorsally, respectively. A dorsoventrally wide, long, and shallow depression opens caudal to the diapophysis between the pcdl and podl. The acdl is a thin strut of bone projecting cranioventrally, terminating near the neurocentral junction. The prezygodiapophyseal lamina (prdl) is not prominent in cranial cervical vertebrae, but in succeeding vertebrae, as the diapophysis migrates caudally and the transverse processes increase in breadth, the prdl expands and forms broad, flat wing-like extensions that connect the prezygapophysis to the diapophysis (Figure 9).
Cervical Ribs. Cervical ribs run subparallel to the length of the vertebral centrum, angled slightly ventrally. The ribs are long; in cranial cervical vertebrae, the ribs overlap with at least two succeeding vertebrae. Cervical ribs are dorsoventrally flattened proximally, becoming more rod-shaped along their length distally (Figure 11).
Dorsal Vertebrae. A total of 14 dorsal vertebrae are known from Jones Ranch. Five dorsal vertebrae have been completely prepared, including two nearly complete, articulated cranial dorsal vertebrae, FWMSH 93B-10-13, which were partially described by Gomani et al. (1999). The remaining three dorsal vertebrae were found in different parts of the quarry and represent different positions within the dorsal series, most likely from more than one individual. In addition, a series of the last six vertebrae of the presacral column preserved in articulation with dorsal ribs, a complete pelvis, and a portion of the hindlimb of a single individual have been partially prepared. However, only the cranial most vertebra in this series is informative, as the caudal five are less exposed and are extensively weathered. Measurable lengths of preserved limb elements from the quarry suggest that all sauropods individuals were comparable in size. Therefore, whereas representing different individuals, dorsal vertebrae from different regions of the presacral series are described based on the assumption that they are directly comparable.
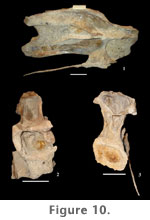
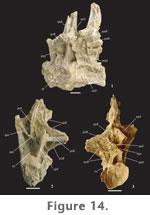
Transverse processes of cranial dorsal vertebrae are robust, expanded dorsoventrally as well as craniocaudally. They are less expanded in caudal dorsal vertebrae. Diapophyses are horizontal in cranial dorsal vertebrae but begin to incline dorsally in the middle of the series and become more strongly angled in caudal dorsal vertebrae, where they form approximately a 30° angle with the neural spine. Neural spines are vertical in cranial dorsal vertebrae but are directed slightly caudally in caudal dorsal vertebrae. This upright orientation is the primitive condition exhibited by all sauropods except somphospondylians. In somphospondylians, dorsal neural spines are strongly angled caudally. In P. jonesi dorsal neural spines are broadly expanded transversely at their base and taper distally. Neural spines do not appear to flare at their distal ends as they do in Brachiosaurus; however, the terminal end of the neural spine is not completely preserved in any of the dorsal vertebrae from Jones Ranch. If dorsal neural spines did flare in P. jonesi, it was very minor. Neural spines have a slightly greater craniocaudal breadth in dorsal vertebrae three and four, due to greater development of the prespinal lamina. There is no evidence of a hyposphene in the first dorsal vertebra. However, a strongly developed hyposphene is present on the caudal surface of the fourth dorsal vertebra. The hyposphene is much reduced on mid-dorsal neural arches, but the condition is presently unknown in caudal dorsal vertebrae. As in the cervical vertebrae, the cranial face of the neural arch in cranial dorsal vertebrae is excavated by symmetrical fossae that occur above the neural canal and below the prezygapophyses.
Several laminae support the diapophysis and parapophysis on the neural arches of the dorsal vertebrae. The first dorsal vertebra has prominent cranial and caudal centrodiapophyseal laminae. The acdl meets the cprl at the craniodorsal margin of the centrum, forming deep infraprezygapophyseal and infradiapophyseal fossae craniodorsal and caudoventral to the acdl, respectively. No second dorsal vertebra has been recognized. In the third dorsal vertebra, the acdl is absent. A pcdl is present at this position in the dorsal series, and a single, large fossa is formed on the lateral aspect of the neural arch. In the fourth dorsal vertebra, the parapophysis has migrated dorsally onto the neural arch, just cranial and ventral to the diapophysis. In this case the parapophysis bisects what was the acdl in the first dorsal vertebra and is supported above and below by thin paradiapophyseal (ppdl) and anterior centroparapophyseal (acpl) laminae, respectively. The acpl appears in the mid- and caudal dorsal vertebrae of all sauropods except Shunosaurus (Wilson 2002). It is slightly longer than the ppdl in the fourth dorsal vertebra of P. jonesi and is oriented almost vertically. In more caudal dorsal vertebrae the ppdl is longer than the acpl. The pcdl is less expansive in mid- and caudal dorsal vertebrae as it is in cranial dorsal vertebrae. A posterior centroparapophyseal lamina (pcpl) first appears in the fourth dorsal vertebra and persists into caudal dorsal vertebrae. The presence of this lamina is a derived character shared by most neosauropods plus Jobaria but has been lost several times in the evolutionary history of sauropods. In dorsal vertebra four the pcpl connects the caudoventral aspect of the parapophysis to the cranial surface of the pcdl near its junction with the top of the centrum. The pcpl essentially divides the large infradiapophyseal fossa in half. Consequently, caudal to the parapophysis, the infradiapophyseal fossa is long and narrow. Below the parapophysis the acpl, pcpl, and dorsal margin of the centrum define the borders of a deep, roughly triangular infraparapophyseal fossa. The acpl and pcpl are less well developed and occur higher on the neural arch in caudal dorsal vertebrae. The conspicuous fossae that occur on the lateral surface of the neural arch in cranial and middle dorsal vertebrae also exhibit a dramatic reduction in the last dorsal vertebrae. In dorsal vertebra three and more caudal vertebrae a short prezygoparapophyseal lamina (prpl) connects the parapophysis to the lateral surface of the prezygapophysis. The podl is short in cranial dorsal vertebrae, terminating proximally on the transverse process. In middle and caudal dorsal vertebrae the podl extends farther onto the caudal surface of the transverse process. A prespinal lamina (prsl) is particularly well developed in the third and fourth dorsal vertebrae as a thin plate of bone spanning the entire length of the neural spine. The prsl is rudimentary in the first dorsal and in caudal dorsal vertebrae and does not extend to the distal end of the spine. A weak postspinal lamina (posl) is visible in dorsal vertebra one, but there is no evidence of a posl in the other dorsal vertebrae where the caudal aspect of the neural spine is observable. In cranial dorsal vertebrae the sprl terminates near the base of the neural spine. In caudal dorsal vertebrae the sprl ends high up on the craniolateral aspect of the neural spine (Figure 14.1). In all dorsal vertebrae the spol originates on the lateral surface of the postzygapophyses and projects dorsomedially on both sides toward the midline of the neural arch forming a broad, V-shaped concave surface behind the neural spine. Only in mid-dorsal vertebrae of P. jonesi is the spol divided. A divided spol is a derived character that is lost in titanosaurians and is unknown in Euhelopus, the most basal somphospondylian, but is present in all other eusauropods, excluding Shunosaurus. All P. jonesi dorsal vertebrae have expanded spinodiapophyseal laminae (spdl), which are responsible for producing a webbed appearance in the space between the neural spine and the diapophysis. In caudal dorsal vertebrae the spol is expanded equal to or more than the spdl and therefore also results in a broadly expanded region at the base of the neural spine. The spdl parallels the spol proximally on mid-dorsal neural arches. It is not clear whether the two laminae meet distally on the spine. Dorsal vertebrae possess a narrow but deep fossa on the caudal half of the lateral aspect of the neural spine that is bounded by the spdl, spol, and podl. One mid-dorsal vertebra, FWMSH 93B-10-27, exhibits a short accessory lamina that extends cranioventrally from the postzygapophysis and connects to the caudal aspect of the pcdl. In FWMSH 93B-10-27 this lamina defines the dorsal margin of a large, triangular fossa. In dorsal vertebra four (FWMSH 93B-10-13), there is an accessory horizontal lamina connecting the hyposphene to the middle of the pcdl, which serves as the dorsal border to the infrapostzygapophyseal fossa below, bounded by the pcdl and cpol cranially and caudally, respectively. The accessory postzygodiapophyseal lamina observed in FWMSH 93B-10-27 occurs above the fossa in FWMSH 93B-10-13. An isolated centrum of a dorsal vertebra (FWMSH 93B-10-48) that has lost most of its exterior bone to erosion reveals a network of thin laminae of bone forming a honeycomb-like pattern. This picture of the internal pneumatic structure of presacral vertebrae in the Jones Ranch sauropod differs from the conclusions reached by Gomani et al. (1999) and Wedel (2003).
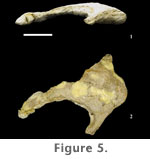
Sacral Vertebrae. A nearly complete sacrum (FWMSH 93B-10-27) is known for P. jonesi but is still undergoing preparation from a massive concretion. Only the ventral surface of the sacrum has been prepared. Four sacral vertebrae are preserved (S2-S5). A dorsosacral vertebra would have contributed a fifth vertebra (S1) to the sacral series but the centrum is missing, and all that is preserved are the transverse processes, which articulate with the ilia at the cranial end of the sacrum. The absence of a sixth sacral vertebra distinguishes P. jonesi from more derived somphospondylian sauropods. The centrum of the second sacral vertebra (S2) is partially exposed in three dimensions. It is small and appears compressed dorsoventrally. Relatively large sacral foramina occur between the transverse processes, or sacral ribs, of the vertebrae. Transverse processes are narrow craniocaudally at mid-length but expand distally and fuse to form the sacrocostal yoke. A more complete description of the sacrum awaits further preparation of the specimen.
Caudal Vertebrae. More than 37 caudal vertebrae have been collected from Jones Ranch. This number includes two associated series: five mid-caudal vertebrae (FWMSH 93B-10-21) and eight articulated distal caudals (FWMSH 93B-10-17). Pleurocoels are absent from the lateral surfaces of the centra, and the lateral depressions that are seen in caudal centra of some sauropod taxa have not been observed in P. jonesi except in one proximal centrum. All caudal vertebrae of P. jonesi are amphiplatyan. Centrum height is less than centrum width in proximal and mid-caudal vertebrae (Table 4). Distal caudal centra are spool-shaped and are typically slightly taller than they are wide. Some mid-caudal centra are rhombus-shaped in lateral view, with the articular surfaces angled craniodorsally (Figure 16.1).
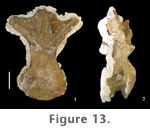
Transverse processes, or caudal ribs, are short and span the neurocentral junction in the first few caudal vertebrae. The processes are triangular in shape and curve slightly in the caudal direction. Transverse processes increase in length in mid-caudals and occur high on the centrum but do not extend onto the neural arch. In mid-caudal vertebrae transverse processes are dorsoventrally compressed and broad craniocaudally at their origin, tapering laterally with a stronger caudal curvature. The neural arch is positioned cranially on the centrum of caudal vertebrae. Neural spines are short in proximal and mid-caudal vertebrae and are slightly caudally inclined (Figure 16.1). Proximal and mid-caudal neural spines flare laterally at the distal end (Figure 16.2–3).
Distinct spinoprezygapophyseal and spinopostzygapophyseal laminae are visible in proximal and mid-caudal vertebrae. Mid-caudal neural arches possess an intraprezygapophyseal lamina (tprl), which together with the sprl on each side of the arch encloses a prespinal fossa above the neural canal (Figure 16.2). Weakly developed prezygodiapophyseal laminae connect the transverse processes to the prezygapophyses in some mid-caudal vertebrae, but are not visible in proximal caudals.
Chevrons. Three chevrons and fragments of at least three others were recovered from the Jones Ranch quarry (Table 5). Chevrons are Y-shaped in craniocaudal view but the rami are weakly forked proximally resulting in a narrow haemal canal.
Appendicular SkeletonPectoral Girdle
ForelimbAlmost every bone from the forelimb is known for Paluxysaurus jonesi with the exception of some manual elements. P. jonesi has a shorter forelimb than hindlimb, which is typical of most sauropods except for Brachiosaurus brancai, B. altithorax, and Cedarosaurus weiskopfae. The forelimb bones of P. jonesi are slender relative to their overall length. Limb ratio measurements for P. jonesi and comparable values for other sauropod taxa are provided in Appendix 1.
The distal end of the humerus is expanded transversely. Distal breadth is approximately twice the minimum breadth of the shaft. On the caudal surface of the distal end, a well-defined olecranon fossa is bounded on both sides by distinct supracondylar ridges that extend from the lateral and medial epicondyles nearly to mid-shaft in some individuals. The distal articular surface is flat but with rugose texture, and in distal view the condyles angle caudolaterally.
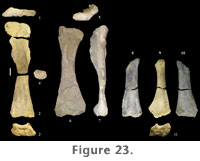
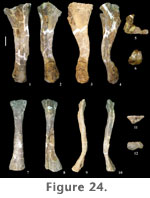
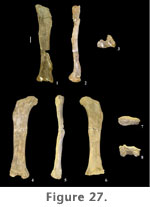
Radius. A complete left radius (FWMSH 93B-10-7) of P. jonesi is preserved, found associated with the ulna described above (Figure 24.7–12). Similar to the ulna, the distal third of the radius is bent proximocaudally relative to the long axis of the shaft. The radius is somewhat shorter than the ulna (Table 7). The radius is compressed craniocaudally along the length of the shaft (Figure 24.9–10). Both ends are expanded transversely, the distal end being slightly more expanded than the proximal end. Distal breadth is close to twice the breadth at mid-shaft. The proximal end is triangular in proximal view, and in distal view the condyle is sub-rectangular, the long axis running mediolaterally (Figure 24.11–12). A prominent ridge begins on the lateral margin of the shaft approximately one-third the length of the bone from the distal end, extends onto the caudal surface about halfway, and terminates at the caudal edge of the proximal condyle (Figure 24.8). This ridge fits into the radial fossa of the ulna. Manus. A nearly complete set of metacarpals was found together along with a distal radius (FWMSH 93B-10-36). In addition to those, two complete, isolated metacarpals are known along with a third that is missing its distal portion. Comparisons of the isolated metacarpals with the articulated hand suggest that the third metacarpal was the longest in the forefoot (Table 7). The longest metacarpal-to-radius ratio could not be determined for a single individual. FWMSH 93B-10-10, identified as a metacarpal IV, was found near the ulna and radius described above (FWMSH 93B-10-7) and may belong to the same individual. A minimum ratio of the longest-metacarpal-to-radius-length using the latter element was calculated to be 0.47 (Appendix 1). Metacarpals of P. jonesi are relatively long and slender compared to diplodocids. Assuming metacarpals of FWMSH 93B-10-36 are identified to the correct position in the manus, metacarpal I is shorter than metacarpal III, which is a primitive condition among sauropods (Upchurch 1998).
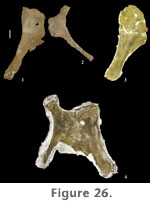
Pelvic GirdleA large sandstone concretion (FWMSH 93B-10-27), containing a sacrum with associated pelvic girdle elements, other hindlimb bones, and dorsal vertebrae was removed from the Jones Ranch locality in 1997 and is currently being prepared in Fort Worth. The following description of the pelvic girdle is based on partially prepared elements of FWMSH 93B-10-27 plus additional material (pubes and ischia) found in the quarry. Ilium. The cranial half of the ilium curves strongly laterally. The medial surface of the cranial half of the ilium faces cranioventrally but is not quite perpendicular to the axis of the sacrum. Its preacetabular process projects craniolaterally and is somewhat tapered. The distance separating the preacetabular processes of the ilia is significantly greater than the craniocaudal length of the ilium (Table 8). The pubic peduncle of the ilium is relatively short, and the ischial peduncle is significantly reduced, as is typical in all sauropods. Cranial to the pubic peduncle the ventral surface of the ilium is roughly straight, as opposed to concave. Pubis. In addition to FWMSH 93B-10-27, an associated pubis and ischium (TMM 42488 JP 1.47, figures 26.1 and 26.2) were collected by J. Pittman in the early years of excavation at Jones Ranch, opposite pubes are associated with an ischium from another individual (FWMSH 93B-10-35, figure 26.3), and an articulated pubis and ischium were recently removed from the field and prepared (FWMSH 93B-10-51, figure 26.4). The pubis is a long, robust bone. The proximal end is moderately expanded craniocaudally and is thickened transversely at the iliac peduncle and acetabular surface. Development of the ambiens process is minor. The shaft of the pubis is uniformly broad craniocaudally and the distal end is slightly expanded transversely relative to mid-shaft. The proximal one-half of the pubis is slightly laterally deflected beginning at the pubic apron, creating an S-shaped pubic symphysis similar to other eusauropods (Wilson 2002). The shaft of the pubis is long relative to the length of the puboischial articular surface, which is a primitive sauropod feature (Wilson and Sereno 1998).
HindlimbComplete examples of upper and lower hindlimb bones are known for P. jonesi but few of the bones of the hindfoot have been identified. As in the forelimb, hindlimb bones are long and slender. Limb ratio measurements for the femur are compared across some sauropod taxa in Appendix 1. Similar comparisons were not made for the tibia and fibula due to the inconsistency in orientation of the tibia and direction of measurement chosen by different researchers for these bones.
At mid-shaft the transverse breadth is nearly twice the craniocaudal width in most individuals (Appendix 1). The proximal one-third of the shaft exhibits the prominent lateral bulge and medial deflection that has been interpreted as a synapomorphy of Titanosauriformes. As in other sauropods, the fourth trochanter is preserved as a broad, low ridge on the caudomedial surface of the shaft. At the proximolateral border of the femur the greater trochanter is not prominent. The proximal surface of the femoral head is notably higher than the proximolateral border. The distal end of the femur is oriented perpendicular to the shaft and distal condyles are well developed. In distal view the condyles trend slightly craniolaterally-caudomedially and are separated by deep intercondylar fossae on both the cranial and caudal surfaces. A lateral epicondyle also appears to be moderately developed at the distal end. Based on closely associated but not articulated humeri and femora, humero-femoral length ratios for P. jonesi fall between 0.85 and 0.88.
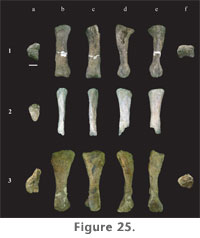
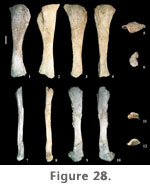
The shaft of the tibia is strongly compressed craniomedially-caudolaterally and is more than twice as broad in the opposite direction, craniolaterally-caudomedially. The tibia is expanded at both ends relative to mid-shaft, but to a much lesser extent at the distal end (Table 9). The distal end is slightly wider transversely than craniocaudally, the articular surface for the ascending process extending out craniolaterally. The craniolateral margin of the shaft is concave, whereas the caudomedial aspect is straight. The proximal condyle is flat and diamond-shaped in proximal view. At the lateral margin, the proximal surface is strongly expanded, forming a broad tubercle that continues a short distance distally along the shaft as a low ridge. The proximal condylar surface, however, is more elongate fore to aft than transversely (Table 9). Paluxysaurus jonesi tibiae possess a pronounced cnemial crest. It is long, projecting craniolaterally, curving laterally at its distal extreme, and angling ventrally from the proximal condylar surface. A faint ridge extends the length of the shaft connecting the cnemial crest proximally with the articular surface for the ascending process at the distal end. Both the tibia and fibula of FWMSH 93B-10-15 were found near a right femur and right fibula (FWMSH 93B-10-25). On average, it is estimated that P. jonesi tibiae reach close to 60% of femur length (Appendix 1). Fibula. Two fibulae from Jones Ranch are in the collections (FWMSH 93B-10-15 and FWMSH 93B-10-25). Both are heavily weathered. As noted above, FWMSH 93B-10-15 was found with a left tibia and FWMSH 93B-10-25 was associated with a right femur. The fibula is a slender bone (Figure 28.8–13) that is slightly longer than the tibia and close to two-thirds the length of the femur (Appendix 1). The proximal and distal ends are moderately expanded relative to mid-shaft, particularly in the craniocaudal direction, as in the tibia. Craniocaudal breadth at mid-shaft is slightly greater than the transverse breadth (Table 9). The distal end of FWMSH 93B-10-25 is expanded transversely to approximately 1.6 times the mid-shaft breadth and in FWMSH 93B-10-15 distal breadth is more than twice mid-shaft breadth. The lateral trochanter is well developed on the lateral surface approximately one-third the distance from the proximal end and is connected to the distal condyle by a low ridge. The proximal end is concave medially and convex laterally. The fibula thins at the proximocranial border and has a small cranial prominence. The tibial articular surface, or tibial scar, is plainly visible. The fibula/femur length ratio is 0.67.
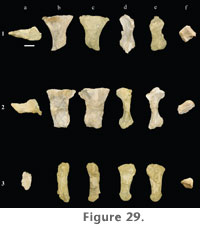
Pes. Only metatarsals I, II, and IV are known for Paluxysaurus jonesi. Excluding fragmentary metapodials that could not be assigned confidently to the fore or hindfoot, no other pedal elements, including phalanges, tarsals, astragali, or calcanei, have been discovered so far. Metatarsals I and II (FWMSH 93B-10-16) are from the right side and were found together in the quarry, while metatarsal IV (FWMSH 93B-10-26) is probably a left and was found as an isolated bone. The shorter metatarsal of FWMSH 93B-10-16 is identified here as metatarsal I but might in fact be metatarsal V.
Metatarsal II is longer than metatarsal I (Table 9). Both are robust bones and are particularly expanded at the proximal end (Figure 29). Proximal condyles of the first two metatarsals are compressed dorsoventrally, and the shaft of metatarsal II is also compressed. The transverse breadth of the distal end of metatarsal II is subequal to mid-shaft breadth. The distal end of metatarsal I is rotated medially with respect to the axis of the shaft. In metatarsal II, the distal end is rotated medially and bevelled proximoventrally with respect to the shaft. A well-developed proximomedial process is present on the ventral surface of metatarsals I and II. In metatarsal I this process extends onto the shaft and nearly reaches the distal end. Metatarsal IV is relatively slender compared to metatarsals I and II. The fourth metatarsal is expanded dorsoventrally at the proximal end. All three preserved metatarsals have well-developed, rounded distal condyles. |
|