|
DISCUSSION
In recent years, the fossil record has seen an increase in the global diversity of sauropods from the Early Cretaceous. In North America, new forms have been described from Utah and Oklahoma. Additional undescribed material was reported from Utah (Britt and Stadtman 1996,
Britt et al. 1997,
1998). Comparison of the Jones Ranch sauropod with specimens from the Early Cretaceous of North America and closely related titanosauriforms from other continents indicates that Paluxysaurus jonesi is morphologically distinct from previously described taxa.
Astrodon johnstoni was named from isolated teeth from the Arundel Formation of Maryland. Leidy (1865) provided a brief description of the teeth.
Marsh (1888) described isolated and associated sauropod material from the Arundel sediments that he referred to the taxon Pleurocoelus nanus, and he referred additional material of what he claimed to be a second, smaller species to Pleurocoelus altus. The distinction made between Pleurocoelus nanus and Pleurocoelus altus is primarily based on size (e.g.,
Marsh (1888),
Gilmore 1921).
Salgado et al. (1995) contended that P. nanus and P. altus are not different size individuals of the same species, but provide no evidence to support this claim. I concur with
Hatcher (1903b) and more recent authors (e.g.,
Carpenter and Tidwell 2005) that there is currently insufficient evidence to recognize more than one distinct species of sauropod from the Arundel Formation (contra
Lull 1911 and
Gilmore 1921).
Hatcher (1903b) argued for synonymizing Astrodon and Pleurocoelus, the former name having precedence; however, Pleurocoelus continues to be used to refer to the material described by
Marsh (1888). Recently,
Carpenter and Tidwell (2005) returned to this topic and suggested that referral of Pleurocoelus type and referred material to Astrodon by
Hatcher (1903b) as first reviser gives Astrodon priority.
Carpenter and Tidwell (2005) did not address the validity of the name Astrodon based only upon teeth. The taxonomy of the Arundel sauropod material is problematic and likely will remain unresolved until additional material is found. Fragmentary sauropod remains from other regions that have been referred to Pleurocoelus (e.g.,
Lydekker 1889,
1890, and
Rich et al. 1983), including specimens from Texas (Langston 1974 and
Gallup 1989), may not be correctly assigned and need to be re-examined. Regardless of the name used for the Maryland taxon, Paluxysaurus can be differentiated from the Arundel sauropod material.
Some differences in morphology between Paluxysaurus and the Arundel sauropod (hereafter referred to as Pleurocoelus for simplicity) can be detected in the skull. Tooth crowns of Paluxysaurus are less compressed than in Pleurocoelus. The caudoventral margin of a small maxilla of Pleurocoelus (USNM 5667) slopes gradually caudodorsally, whereas this portion of the maxilla is more steeply angled in Paluxysaurus. The palatine shelf on the medial surface of the maxilla is narrower dorsoventrally and is more prominent than in Pleurocoelus, and the dorsal surface of the palatine shelf is broadly concave in Paluxysaurus while it is nearly horizontal in Pleurocoelus.
Few diagnostic features can be observed in the axial skeleton of Pleurocoelus. Nearly all of the vertebrae referred to this taxon lack neural arches so they are of limited comparative value. Presacral bone texture of Pleurocoelus is solid but vertebral centra are excavated by large, deep lateral pneumatic fossae. Presacral vertebrae of Paluxysaurus jonesi are more cavernous and more closely resemble vertebrae of Brachiosaurus in their internal structure. However, as noted by
Wedel (2003), all known vertebrae of Pleurocoelus are from juvenile individuals, and it is not well understood how vertebral pneumaticity in sauropods changes with ontogeny.
The appendicular material of Pleurocoelus, however, is more distinctive. Paluxysaurus jonesi differs from Pleurocoelus in having an expanded distal scapular blade that is not rounded on the acromial side. Although the distal end of the Paluxysaurus scapula is expanded on the acromial side, it is distinctly different from a distal scapular blade referred to Pleurocoelus (USNM 8474) and the scapulae of Camarasaurus and Brachiosaurus, in that these taxa have a rounded expansion on the acromial side of the distal scapular blade, but the opposite edge of the blade is nearly straight. In Paluxysaurus, the scapula expands on both the acromial and glenoid margins.
Limb bones of Paluxysaurus are generally more slender than those of Pleurocoelus (Appendix 1). A juvenile femur of Pleurocoelus (USNM 5696) exhibits a lesser degree of medial deflection of the proximal end than the femora of Paluxysaurus, and the greater trochanter of the femur is less prominent in the Texas sauropod. The shape of the proximal condyle of a tibia referred to Pleurocoelus (USNM 5657) is more rounded (less craniocaudally expanded) than in Paluxysaurus.
Cedarosaurus weiskopfae is known from a partial skeleton of a single individual (DMNH 39045) from the Yellow Cat Member of the Cedar Mountain Formation in east-central Utah.
Tidwell et al. (1999) considered the new taxon to be a brachiosaurid, most closely related to "Pleurocoelus."
The most conspicuous differences in morphology between Paluxysaurus jonesi and Cedarosaurus weiskopfae occur in the caudal vertebral series. Caudal neural spines project craniodorsally from the neural arch in cranial to mid-caudals of C. weiskopfae. All caudal vertebrae of P. jonesi have neural spines that are inclined caudodorsally. Distal caudal centra of P. jonesi are shorter than in C. weiskopfae and are more rounded, with height-to-width ratios, on average, slightly greater than 1.0. Centra of distal caudals in C. weiskopfae are noticeably wider than tall, some having height-to-width ratios as low as 0.58.
Chevrons of P. jonesi have a relatively short haemal canal, comprising less than 40% of the overall length of the bone. A short haemal canal is a feature shared by Brachiosaurus and all more primitive sauropods. The only chevron preserved for C. weiskopfae appears to have a broader, deeper haemal canal; however the shaft of the chevron is broken and it is unclear how long this segment of the bone was.
Sternal plates of P. jonesi and C. weiskopfae are similarly shaped but in the former the bone is less broad transversely. In C. weiskopfae, sternal plate length is 73% greater than the transverse breadth. The sternal plate of P. jonesi, however, is twice as long as it wide. Paluxysaurus jonesi has a shorter acromion processes at the proximal end of the scapula than C. weiskopfae. The acromial side of the proximal end of the scapula in C. weiskopfae is markedly expanded in comparison to P. jonesi. The coracoid of P. jonesi is relatively longer proximodistally than C. weiskopfae and has a more rounded cranioventral margin.
The humerus of C. weiskopfae is slightly more slender than that of P. jonesi (Appendix 1). Paluxysaurus jonesi exhibits a greater humero-femoral length ratio than what is observed in more primitive sauropods, but the forelimb is distinctly shorter relative to the hindlimb compared to the condition in C. weiskopfae and Brachiosaurus. The ulna of P. jonesi is broader at mid-shaft than is C. weiskopfae, and the olecranon region is raised slightly higher above the proximal condyle in the former. The craniolateral arm of the proximal condyle of the ulna is significantly longer in P. jonesi, and the craniomedial arm is broader compared to C. weiskopfae. In C. weiskopfae, the proximal one-half of the preserved segment of the ulna is expanded craniomedially, resulting in a concave craniomedial margin of the shaft. The ulna of P. jonesi is less expanded craniomedially at the proximal end and below the proximal condyle. Cedarosaurus weiskopfae is primitive with respect to P. jonesi and other sauropods, except for Venenosaurus dicrocei, Diplodocus sp., and Vulcanodon karibaensis, in having a radius with a narrow, rounded distal condyle. In other sauropods the distal end of the radius is expanded transversely relative to mid-shaft and the distal condyle is more rectangular in shape (Appendix 3,
Appendix 4).
The proximal end of the C. weiskopfae pubis is more expanded cranially relative to P. jonesi. The pubic peduncle of the ischium is broader in P. jonesi, whereas the iliac peduncle is more expanded craniocaudally in C. weiskopfae. In P. jonesi, the femur exhibits a greater degree of medial deflection along the proximal one-third of the shaft and a less prominent greater trochanter than in C. weiskopfae. The proximal tibia of P. jonesi is more expanded craniocaudally compared to C. weiskopfae and has a more prominent cnemial crest. Metatarsal II of P. jonesi differs from C. weiskopfae in that it possesses a strong medial process at the distal end. The shaft of metatarsal II in C. weiskopfae is more concave medially than in P. jonesi.
Venenosaurus dicrocei comes from the Poison Strip Member of the Cedar Mountain Formation in Utah, which is stratigraphically younger than the Yellow Cat Member that produced Cedarosaurus weiskopfae. Venenosaurus dicrocei is represented by a partial skeleton of a single individual (DMNH 40932). Some juvenile remains from the same locality may also be referable to this species.
Tidwell et al. (2001) identified V. dicrocei as a titanosauriform that is intermediate in its morphology between Brachiosaurus and titanosaurians.
Caudal centra of Paluxysaurus jonesi lack the fossae present on the lateral surfaces of cranial caudal vertebrae of V. dicrocei. Like Cedarosaurus weiskopfae, mid-caudal neural spines of V. dicrocei are directed cranially, a condition that is unique to these two taxa.
The glenoid process of the scapula is weakly developed in P. jonesi compared to V. dicrocei, in which the glenoid process is quite long. Similar to C. weiskopfae, the craniolateral arm of the proximal ulnar condyle is short in V. dicrocei relative to P. jonesi. The proximal condylar arms in V. dicrocei are straight, robust processes. The ulna of V. dicrocei possesses a shallower radial groove than P. jonesi and C. weiskopfae. As in C. weiskopfae, V. dicrocei possesses a narrow, rounded distal radial condyle that is in sharp contrast to the distal radius of P. jonesi.
The pubis of P. jonesi differs from V. dicrocei in that the cranial edge of the pubic blade is straight, as opposed to concave in V. dicrocei. The ischium of P. jonesi exhibits a less pronounced caudal curvature than V. dicrocei.
Similar to C. weiskopfae, the shaft of metatarsal II in V. dicrocei has a stronger medial concavity than in P. jonesi. The proximomedial process on the caudal surface of metatarsal II in P. jonesi is positioned closer to the midline of the bone in V. dicrocei. Metatarsal IV of P. jonesi is missing a portion of the medial surface, but the bone is more robust than the fourth metatarsal of V. dicrocei.
The type specimen of Sauroposeidon proteles, which consists of four articulated cervical vertebrae (OMNH 53062), is known from the Antlers Formation of southern Oklahoma.
Wedel et al. (2000a,
2000b) concluded that S. proteles is most closely related to Brachiosaurus brancai. Elongation indices (EI) for cervical centra of Paluxysaurus jonesi overlap with the ranges for both S. proteles and B. brancai. Cervicals of B. brancai generally have EI values below 5.0, but reach a maximum of 5.4 (Wedel et al. 2000b). In addition, EI values for the sixth cervical vertebra in individuals of B. brancai vary by as much as 0.9. In S. proteles, EI values range from 4.6 to 6.1 (Wedel et al. 2000b). Similarly, P. jonesi cervical vertebrae exhibit a large range in EI values, and the centrum length-to-caudal height ratios are similar to Malawisaurus dixeyi, a significantly smaller taxon. Overlap in EI values among sauropod taxa of different ages and of different recent ancestry, demonstrates that length of cervical centra relative to caudal height is not necessarily a useful character for distinguishing between taxa.
Cervical vertebrae from Jones Ranch differ from S. proteles and B. brancai in having a well developed, long anterior centrodiapophyseal lamina (acdl). This lamina is short in B. brancai and cannot be seen in lateral view in illustrations of the cervicals provided by
Janensch (1950). The acdl is completely absent from the four cervicals of S. proteles. Neural arches of mid-cervical vertebrae of P. jonesi are broader than in S. proteles and B. brancai. Mid-cervicals of P. jonesi have prominent, transversely expanded prezygodiapophyseal laminae (prdl) (see
Figure 10.2). In both S. proteles and B. brancai transverse processes are less broad and thus the prdl is less expanded.
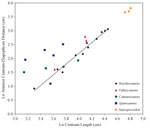 Wedel et al. (2000a,
2000b) cited the extreme caudal position of the diapophyses on the neural arch of S. proteles cervicals as a character that distinguishes it from B. brancai and other sauropods. However, the distance of the diapophysis from the cranial end of the centrum (minus the cranial condyle) in S. proteles increases with total centrum length serially along the column by the same proportion as in B. brancai (Figure 30). This suggests that the difference observed in the relative position of the diapophysis in cervicals of S. proteles and B. brancai may simply be a function of size of the individual. In cervical vertebrae of P. jonesi, Camarasaurus, and Apatosaurus, distance of the diapophysis from the cranial end of the centrum varies relative to centrum length by a different scaling factor or shows no significant relation. Wedel et al. (2000a,
2000b) cited the extreme caudal position of the diapophyses on the neural arch of S. proteles cervicals as a character that distinguishes it from B. brancai and other sauropods. However, the distance of the diapophysis from the cranial end of the centrum (minus the cranial condyle) in S. proteles increases with total centrum length serially along the column by the same proportion as in B. brancai (Figure 30). This suggests that the difference observed in the relative position of the diapophysis in cervicals of S. proteles and B. brancai may simply be a function of size of the individual. In cervical vertebrae of P. jonesi, Camarasaurus, and Apatosaurus, distance of the diapophysis from the cranial end of the centrum varies relative to centrum length by a different scaling factor or shows no significant relation.
SMU 61732 consists of a series of 21 caudal vertebrae, fragments of cervical and dorsal vertebrae, a chevron, dorsal ribs, a distal scapula, and two teeth. The teeth from SMU 61732 are not well preserved. They are generally similar in size and morphology to those from Jones Ranch, FWMSH 93B-10-33 in particular (Figure 6.9–11), but possess a high-angled planar wear facet as opposed to the V-shaped wear facets observed in most of the teeth from Jones Ranch. The difference in wear morphology may simply be a function of position in the tooth row. The one cervical and one dorsal vertebra from the Wise County locality both have badly crushed neural arches and reveal little about their morphology. No obvious differences can be seen from cervical and dorsal centra from Jones Ranch beyond what could be considered serial and/or individual variation in sauropod presacral vertebrae. From what is preserved of the neural arch, the cervical vertebra of SMU 61732 has prominent diapophyseal laminae as in cervicals from Jones Ranch.
Caudal centra of Paluxysaurus jonesi have flat articular surfaces whereas cranial caudal centra of SMU 61732 have moderately concave cranial articular surfaces and slightly convex caudal faces. A distinct hyposphene articulation is visible on the neural arch of a cranial to mid-caudal vertebra of SMU 61732. Nothing similar to this is preserved in the caudals known from Jones Ranch. Cranial and mid-caudal vertebrae of P. jonesi have transversely broader neural spines. Differences portrayed in the caudal vertebrae from Jones Ranch and SMU 61732 may be misleading if the vertebrae represent different positions in the tail. If that is the case, considering the amount of serial variation in sauropod caudal vertebrae, these differences seem less significant. Procoelous and amphiplatyan centra are found in the tail of Malawisaurus dixeyi, a basal titanosaurian sauropod from the Early Cretaceous of East Africa. In M. dixeyi, however, the procoelous caudals occur at the front of the tail, and caudal vertebrae from the cranial portion of the tail in P. jonesi (FWMSH 93B-10-14) are clearly not procoelous.
The distal end of the scapula of P. jonesi is slightly more expanded on the acromion side than in SMU 61732. Consequently, the acromial, or craniodorsal, margin of the scapular blade appears concave while it is straighter in the Wise County specimen. In scapulae of Brachiosaurus brancai significant variation exists in the amount of expansion to the acromial side of the distal blade (see
Janensch 1961; plate 15, figures 1 and 2). The difference in morphology between the scapulae of Palauxysaurus jonesi and SMU 61732 noted here is no greater than the range of variation seen in Brachiosaurus brancai. The shape of the distal scapula of SMU 61732 differs from the condition in Pleurocoelus, the genus that
Langston (1974) referred this specimen to. Despite the differences noted above between Paluxysaurus jonesi and SMU 61732, from the currently available material, the latter cannot confidently be distinguished as a separate genus.
Cloverly Formation
Sauropod bones from the Cloverly Formation in Wyoming and Montana were described by Ostrom (1970), which he tentatively referred to the "Titanosauridae." Two dorsal vertebrae of the Cloverly sauropod (YPM 5449) figured by
Ostrom (1970, plate 13f and 13g) are similar to those from Jones Ranch and may be referable to the same genus. However, noticeable differences in morphology exist between the specimens that may preclude from calling them the same species. Dorsal vertebrae of P. jonesi have slightly taller neural arches than the dorsals of YPM 5449 and the centrum of the first dorsal vertebra of P. jonesi (FWMSH 93B-10-11) has a caudoventral lip that is absent in the cranial dorsal of the Cloverly specimen. Postzygapophyses are horizontal in FWMSH 93B-10-11, whereas they are angled in the Cloverly sauropod, and the neural spine is slightly broader in the P. jonesi vertebra. Neural spines of caudal dorsals of the Cloverly sauropod are slightly more caudally inclined than those of the Texas sauropod. Pleurocoels on the lateral surfaces of centra of Jones Ranch dorsal vertebrae are generally eye-shaped, as opposed to the more oval cavities in the Cloverly sauropod. Apart from these minor differences, the cranial dorsal vertebra of YPM 5449 is virtually indistinguishable from FWMSH 93B-10-11.
Appendicular elements from the Cloverly Formation show fewer similarities to P. jonesi than do the dorsal vertebrae. The dorsal vertebrae of YPM 5449 are close to the size of dorsals of P. jonesi. However, an ulna associated with the same dorsal vertebrae of the Cloverly sauropod is approximately 20 cm shorter and more robust than the ulna of P. jonesi. Humeri of P. jonesi differ from an isolated, partial humerus from the Cloverly Formation (YPM 5452) in that the proximal surface is inclined medially, forming an angle greater than 90° at the proximolateral border. In the Cloverly specimen, the proximal surface is flat and forms a right angle with the lateral margin of the shaft. TMM 42488 JP 1.21 from Jones Ranch has a more horizontal proximal surface than other humeri from Jones Ranch and resembles the Cloverly humerus more than FWMSH 93B-10-2 and FWMSH 93B-10-7. The proximomedial border of the Cloverly humerus is more expanded and pointed, and the medial edge of the shaft is more concave than in humeri of P. jonesi.
A right femur of the Cloverly material (YPM 5451) is missing the proximal end. The measured distance between the distal end of the femur and the fourth trochanter is greater than that in femora from Jones Ranch, suggesting that the complete length of YPM 5451 would have exceeded that known for P. jonesi. An isolated left tibia (YPM 5450) from the Cloverly Formation is longer and more slender than tibiae of P. jonesi. In the latter, tibiae are more expanded at the proximal end. Moreover, the dorsal margin of the cnemial crest occurs slightly below the proximal condyle, whereas in the Cloverly tibia the top of the cnemial crest and the proximal condyle appear to be at the same level. Teeth of P. jonesi are broadly similar in form to isolated teeth from the Cloverly Formation but have a tendency to be broader at mid-crown.
Maxillae of Paluxysaurus jonesi and Euhelopus zdanskyi exhibit some obvious differences in morphology. The ascending, or nasal, process in P. jonesi is relatively short and more than twice as broad as it is E. zdanskyi. The nasal process of the maxilla is angled less caudally (i.e., more vertical) in P. jonesi, forming a broader cranioventral margin of the antorbital fenestra. Teeth of P. jonesi have a more prominent longitudinal ridge on the lingual surface of the crown. Presacral vertebrae of P. jonesi lack the bifurcating neural spines that are present in E. zdanskyi, and in contrast to the Asian taxon, cranial cervical centra of P. jonesi are wider than they are tall (Table 2).
Cranial cervical vertebrae of Paluxysaurus jonesi are transversely narrower than those of Phuwiangosaurus sirindhornae. As in Euhelopus zdanskyi, P. sirindhornae has bifid caudal cervical and cranial dorsal neural spines, which is absent in P. jonesi. The glenoid process of the scapula is directed more ventral and the distal humerus is more transversely expanded in P. sirindhornae. The craniomedial process of the proximal condyle of the ulna in P. jonesi is roughly horizontal and level with the craniolateral arm compared to P. sirindhornae, in which the craniomedial arm is directed ventrally. Cranial caudal neural spines are more vertically oriented in P. sirindhornae and are not transversely expanded at the distal end. Distal caudal centra of P. jonesi are shorter than in P. sirindhornae.
|