|
|
|
COMPARISONS AND SIGNIFICANCE
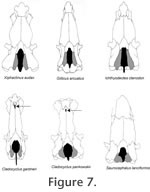
There are two nominal species of Cladocyclus; the type species C. gardneri Agassiz 1841 and C. ferus Silva Santos 1950 from the Apto-Albian Romaldo Member of the Santana Formation of Brazil. Patterson and Rosen (1977) noted that there were considerable differences between some of their specimens that they regarded as C. gardneri and cf. C. ferus, chiefly in the nature of the margins of the opercular bones. Castro Leal and Brito (2004) found specimens showing combinations of both these features and concluded that the genus was monospecific. We have no knowledge of the opercular bones in the specimen from the Kem Kem Beds described here but note that there is a marked difference in the skull roof proportions between C. pankowksii: length/breadth ratio = 2.1 versus 3.1 in C. gardneri, the rostrodermethmoid is blunt in C. pankowskii and pointed in C. gardneri, and the angle of the flexure of the parasphenoid at the level of the basipterygoid process is 155o in C. pankowskii and 130o in C. gardneri. The hyomandibular facet of C. pankowskii is divided by a narrow bridge of perichondral bone not present in C. gardneri. Other ichthyodectiforms with which C. pankowskii might be compared are Chiromystus mawsoni Cope1886 from the Late Hauterivian – early Barremian of Recôncavo Basin and Aptian-early Albian of Sergipe basin, Brazil (Maisey 2000), Ichthyodectes bardacki Cavin 1997 from the Turonian of Goulmima, Morocco, and Unamichthys espinosai Alvaredo-Ortega 2004 from the middle or late Albian of the Tlayúa Formation, Mexico. Chiromystus shows well-developed parieto-epiocciptal crests and thus differs in this respect. However, the palatine articulatory surface upon the ethmopalatine is developed as a groove as in Cladocyclus. Ichthyodectes bardacki differs in that the parieto-epioccipital crests are well-developed, the ethmopalatine has a complex double faceted articulatory condyle and the jugular vein appears to have been completely bone enclosed through its traverse through the otic portion of the braincase. In all of these features I. bardacki appears more derived and similar to Xiphactinus, Ichthyodectes and Gillicus. Unamichthys espinosai differs in that it shows paired parietals (Alvaredo-Ortega 2004). The presence of species of Cladocylcus in the mid-Cretaceous of Morocco and North Eastern Brazil is concordant with other similar distributions of sister-group taxa of fishes and increases an already large body of evidence suggesting the contiguity of these two parts of the world at this time. Maisey (2000, figure 3) plots many of these comparisons that involve at least six separate fish clades. To this we can now add sister species of Calamopleurus (Forey and Grande 1998) and now a sister species of Cladocyclus. |
|