|
|
|
SYSTEMATIC DESCRIPTIONSubdivision Teleostei
Müller, 1846 Diagnosis. Cladocyclus differing from the type species in the proportions of the skull roof; being 2.1 times as long as wide (excluding the supraoccipital crest) versus 3.1 times as long as wide in the type species: no fenestra present between the parietal, supraoccipital and epioccipital (present in type species): rostrodermethmoid with blunt anterior margin versus anteriorly-projecting margin in type species: angle of parasphenoid is 155o versus 130o in type species. Holotype. United States National Museum of Natural History USNM PAL 521360. Nearly complete braincase missing only part of the left basipterygoid process and edge of supraoccipital spine. Currently the only specimen known.
Etymology. Species named after Mr. Mark Pankowski of Rockville, Maryland, who kindly donated the specimen to the United States National Museum of Natural History, Washington.
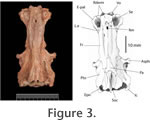
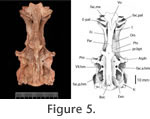
The ethmoid region is completely ossified. All of the ethmoid bones are securely sutured to one another. This is usual for most ichthyodectiforms except for the more primitive forms ( Allothrissops, Eubiodectes, Thrissops and Occithrissops where many spaces persist between the bones). The rostrodermethmoid is relatively small (cf. Saurocephalus) and carries anterolaterally and ventrally projected processes that suture with the ethmopalatine of either side. On the dorsal surface the rostrodermethmoid extends posteriorly as paired splints of bone that suture with the frontals. The diverging splints of the median rostrodermethmoid are matched by opposing diverging ends of the paired frontals, such that there is a median fenestra left between these three bones. A small portion of the supraethmoid is seen to partially occlude the fenestra. A similar fenestra and dorsal expression of the supraethmoid is seen in Cladocyclus gardneri. It is absent from other ichthyodectiforms in which this region of the skull is sufficiently well-known. The supraethmoid forms the dorsal wall of the nasal capsules. As usual the lateral ethmoid is robust and tightly sutured to the supraethmoid, rostrodermethmoid and ethmopalatine. With the last mentioned bone it forms the articulatory facet for palatine. This facet is transversely elongate as it is in Cladocyclus gardneri but unlike the rounded facet seen in Xiphactinus, Ichthyodectes and Gillicus. The extreme ventral edge of the lateral ethmoid is open-ended and must have passed into cartilage in life. The lateral ethmoid is pierced by a large olfactory foramen. The ethmopalatine ('rhinal' of Taverne 1977, 'parethmoid' of Bardack 1965) is very well-developed and forms much of the floor, as well as the inner and posterior walls of the nasal capsule. Anteriorly the ethmopalatine contacts the rostrodermethmoid through a complex suture. The anterior margin of the ethmopalatine is sutured to the vomer, and these two bones form a cup-shaped articulatory facet for the head of the maxilla. Posteriorly it overlies the lateral ethmoid in a splint-like suture. The size and mutual relationships of the ethmopalatine to surrounding bones is very similar to that of Cladocyclus gardneri and saurodontids and ichthyodectids such as Xiphactinus, Ichthyodectes and Gillicus. In primitive ichthyodectiforms such as Allothrissops the ethmopalatine is very small and lies wholly beneath the lateral ethmoid and rostrodermethemoid, which contact each other (Patterson and Rosen 1977, figure 7A); Occithrissops appears to lack an ethmopalatine (Schaeffer and Patterson 1984). The frontal is a long narrow bone resembling most closely that of Cladocyclus gardneri. Differences from other ichthyodectiforms in the lateral margins have already been mentioned. One or two pores are seen on the frontals but there is no clear indication of the path of the supraorbital sensory canal. Posteriorly, there are sinusoidal sutures with the median parietal and the pterotic. On the left side of USNM PAL 521360 there is a very short contact with the epioccipital, which is due to the extreme forward projection of this latter bone in this species, not seen in Cladocyclus gardneri or, indeed, other ichthyodectiforms. The median parietal is placed well-forward at the level of the autosphenotic and lies wholly anterior to the supraoccipital. This is the condition in ichthyodectoids and like those taxa the parietal is pointed anteriorly and expanded posteriorly. Among primitive ichthyodectiforms such as Occithrissops (Schaeffer and Patterson 1984, figure 25), Allothrissops (Patterson and Rosen 1977, figure 5) and Thrissops (Taverne 1977, figure 5) the parietals are small, paired and confined to the hind margin of the skull and meet in the midline above the supraoccipital (the usual condition in primitive teleosts). The parietal is smooth. There is no trace of a pit line as is present in Cladocyclus gardneri. However, this absence may be preservational since such pit line grooves are easily abraded post-mortem. The supraoccipital is large and well-expressed on the dorsal surface. Again, this is similar to the supraoccipital in ichthyodectoids, including Cladocyclus gardneri. The remains of the base of a supraoccipital crest are preserved but clearly most of the crest has been broken off, and it is impossible to be certain of its extent. It must have extended down the posterior face of the supraoccipital as well as the dorsal surface, a similarity with Cladocyclus gardneri. In most ichthyodectoids the crest is very large. In dorsal view the margins of the supraoccipital describe an ellipse. This is very similar to the supraoccipital in Cladocyclus gardneri but is unlike the narrow parallel-sided profile seen in Xiphactinus, Ichthyodectes and Gillicus. In Saurocephalus the supraoccipital outline is also approximately ellipsoidal but the posterior margins are excavated by the neighbouring epioccipitals (Stewart 1999, figure 7). The epioccipital contacts the supraoccipital, parietal, pterotic and frontal through complex interdigitating sutures. It is perfectly smooth: that is, there is no parietal-epioccipital crest as prominently developed in Xiphactinus, Ichthyodectes, Gillicus and Saurocephalus. The contact between the epioccipital and the supraoccipital extends for only about half the length of the supraoccipital. This is similar to Cladocyclus gardneri and also Saurocephalus lanciformis (Stewart 1999, figure 7) but unlike conditions in Xiphactinus, Ichthyodectes and Gillicus where the epioccipital lies along the greater part of the supraoccipital. According to Maisey (1991:195) Chiromystus and Chirocentrites are similar to Cladocyclus in this respect. The pterotic is a large bone as in most ichthyodectiforms. It reaches well-forward above the autosphenotic and carries the otic branch of the sensory canal. It also forms most of the dilatator fossa as well as the posterior half of the anterior facet for the hyomandibula and the anterior half of the posterior facet for the hyomandibula. In posterior view it can be seen to form the roof of the small posttemporal fossa. The autosphenotic is developed as a prominent process that projects directly laterally, not forward as in Xiphactinus, Ichthyodectes and Gillicus. The process is of complex shape and must have provided firm anchorage for dilatator operculi muscles. The autosphenotic forms a small portion of the anterior facet for the hyomandibula.
The exoccipital forms the posterior wall of the subtemporal fossa and part of the posterior wall of the braincase. It has complex interdigitating sutures with the basioccipital and intercalar but a simple suture with the prootic and pterotic. The exoccipitals of either side meet in the midline to form the floor of the cranial cavity. On the lateral aspect a small, posteriorly directed foramen can be seen exiting the cranial cavity immediately behind the posterior exit of the jugular vein. The size of this foramen suggests that it carried the supratemporal branch of the glossopharyngeal nerve. On the posterior face of the exoccipital there is a very large foramen that leads to a canal penetrating the floor of the cranial cavity. This is the combined foramen for the glossopharyngeal and vagus nerves. Such a large combined foramen is a common feature in ichthyodectoids. However, in saurodontids and ichthyodectids this foramen also carried the jugular vein. The intercalar is developed in typical ichthyodectiform fashion, having complex sutures with all surrounding bones. It is exposed on the dorsal surface of the skull where it is developed as a boss above the foramen for the glossopharyngeal+vagus. It is probable that the ventral limb of the posttemporal anchored to the braincase at this point. The intercalar forms the posterior end of the posterior facet for the hyomandibular. The basioccipital forms the entire occipital condyle and becomes constricted as it extends anteriorly to contact the prootic. There is a median orbitosphenoid, perforated anteriorly to allow the passage of the olfactory tracts through the roof of the orbit.
The pterosphenoid forms part of the margin of the optic foramen. The surface is marked by a prominent longitudinal groove that presumably carried the anterior dorsal lateral line nerve. The basisphenoid is robust. Dorsally there are laterally diverging wings that subtend the ventral border of the optic foramen. These wings suture with the pterosphenoid and prootic of either side. The basisphenoid pedicel is broad, laterally compressed with a tight suture with the parasphenoid. The parasphenoid extends nearly the entire length of the braincase, being sutured to the vomer anteriorly and ending as posteriorly diverging wings that embrace the basioccipital. Below the orbit the parasphenoid is triangular in section. Beneath the otic region it is flat. In lateral profile there is a marked angle at the level of the basipterygoid processes and separating the orbital from the otic limbs. The angle between these two limbs is 155o. The basipterygoid process of either side is stout and extends laterally and slightly dorsally (in most ichthyodectoids it reaches anteroventrally). The base of the basipterygoid process is penetrated by two foramina. The anterior foramen carried the efferent pseudobranchial artery into the base of the orbit. The posterior foramen carried the internal carotid artery. The hyomandibular facet is clearly divided to an anterior and a posterior facet separated by a narrow bridge of perichondral-covered bone. This appears different from Cladocyclus gardneri where the facet, although clearly bilobed, is continuous. In saurodontids and ichthyodectids it is also continuous. The subtemporal fossa is extremely deep as in all ichthyodectoids. It reaches well dorsally, so much so that there is a fenestra between it and the posttemporal fossa.
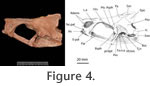
The dorsal, oral margin of the dentary is sinusoidal in outline, rising to a low coronoid process posteriorly. The most characteristic feature is the nature of the dentition. There are 15 teeth or sockets arranged in a single row. The size of the teeth varies. The two anterior teeth are very large. Although the tips of these teeth are not preserved, the diameters of the bases (6.4mm) are larger than those of the complete teeth (5.6 mm) seen more posteriorly. These complete teeth measure 22 and 23 mm in length, suggesting that the symphysial teeth were longer. Behind the symphysial teeth there is the base of a broken tooth of about the same diameter. This is followed by sockets and bases of much smaller teeth. At the level of the dorsal convexity in the oral margin there are three large teeth, two complete with the base of a particularly large tooth between them. The base of this large tooth measures 7.7 mm and appears to be the largest in the jaw. These teeth are followed by seven smaller teeth. There is no simple gradation of tooth size: rather, adjacent teeth may be of very different sizes. It is possible that this is due to replacement teeth lying alongside mature teeth. Each tooth is rounded or slightly ovoid in cross section. The bases are hollow, the surface is smooth and the complete teeth are slightly curved posteriorly. They also show acrodin caps at their tips. |
|