Results
 Morphology of the teeth
Morphology of the teeth
The cheek teeth of the studied taxa are all brachydont to subhypsodont and relatively low crowned. Lots of the studied teeth are medium to strongly worn and only for some taxa unworn or little worn teeth were observed. The available teeth of the different taxa were not of similar wear stage (e.g.
Figure 1).
Some representative dentitions of Capacikala, "Capatanka" and Palaeocastor arranged with increasing wear are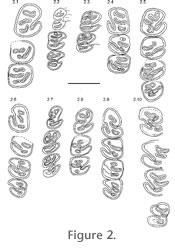 illustrated in
Figure 2,
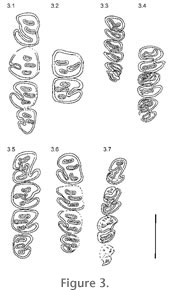
Figure 3,
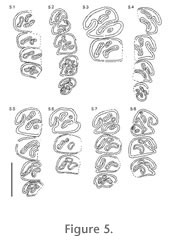
Figure 4, and
Figure 5. The premolars are the largest teeth in the tooth rows, but especially, with little wear, the size difference to molars is small. Whereas germs of all mandibular cheek teeth are slightly rectangular, slightly longer than wide, with wear the lower molars m1/2 become more and more square, m3 more or less rounded.
illustrated in
Figure 2,
Figure 3,
Figure 4, and
Figure 5. The premolars are the largest teeth in the tooth rows, but especially, with little wear, the size difference to molars is small. Whereas germs of all mandibular cheek teeth are slightly rectangular, slightly longer than wide, with wear the lower molars m1/2 become more and more square, m3 more or less rounded.
 The same can be observed in the maxillary cheek teeth, but here molars M1/2 are more square in the beginning and become more oval (broader than long) with wear and M3 more triangular or rounded. The tendency to develop oval M1/2 with wear seems to be strongest in Palaeocastor and here also the M3 seem to be more oval than in the other taxa.
The same can be observed in the maxillary cheek teeth, but here molars M1/2 are more square in the beginning and become more oval (broader than long) with wear and M3 more triangular or rounded. The tendency to develop oval M1/2 with wear seems to be strongest in Palaeocastor and here also the M3 seem to be more oval than in the other taxa.
Overall, the morphology of the cheek teeth appears quite similar and comparatively simple with para-"fossette", meso-"fossette", meta-"fossette" and hypoflexus/id when little to strongly worn. The meta-"fossette" tends to be lost in strongly worn teeth, especially in maxillary teeth. Tooth germs and teeth with little wear are slightly more complex in the occulsal pattern, (e.g. Capacikala gradatus SDSM 53343), sometimes with irregular mesoflexids and connections between "fossettes." The parafossettes are more variable in form, often hook-shaped, in p4 than in m1-3, where they are straighter and more or less rectangular to the lingual side of the tooth.
 They are long, nearly extending from side to side of the tooth in little to medium worn lower cheek teeth, but become shorter in medium to strongly worn teeth.
They are long, nearly extending from side to side of the tooth in little to medium worn lower cheek teeth, but become shorter in medium to strongly worn teeth.
 In maxillary tooth rows P4 is the largest tooth. This becomes more pronounced with wear. Most medium to strongly worn upper molars show only hypoflexus/fossette and mesofossette, strongly diagonally oriented and sometimes curved.
In maxillary tooth rows P4 is the largest tooth. This becomes more pronounced with wear. Most medium to strongly worn upper molars show only hypoflexus/fossette and mesofossette, strongly diagonally oriented and sometimes curved.
The number of fossettes in the anterior and posterior part of the teeth is given in
Table 1. Due to little material in some cases it was not attempted to differentiate according to wear stage here. The Chi2 test including all studied taxa indicates no significant correlation between wear stage and number of anterior fossettes in p4 and P4 and to posterior fossettes in p4, m3, P4, and M1/2. Overall more variability in the number of fossettes can be observed in teeth of no or little wear.
Besides hypostiae/iids and mesostriae/iids, short notches were observed in only
a few cases: a metastriid was found on the m3 of Capacikala gradatus (SDSM
5982); a parafossette with an anterior opening was found on the p4 of
Palaeocastor nebrascensis (UCMP 114452) and P. fossor (F:AM 64188); a
short parastriid or a lateral opening of a paraflexid was found on p4 of
Capacikala gradatus (LACM 9595); a parastriid and metastriid was found on m3
of the same species (LACM 9405); and a probable metastriid was found on m3 of
Palaeocastor sp. (LACM 9494). No such notches were observed in
Capatanka cankpeopi.
 Morphometrics of the Teeth
Morphometrics of the Teeth
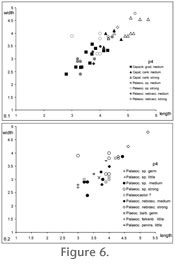
The length x width diagrams (Figure 6 and
Figure 7) illustrate the size of cheek teeth of the studied material. All teeth and teeth of different wear stages have been compared separately. All p4 of Capacikala gradatus, "Capatanka" cankpeopei, Palaeocastor sp. and P. nebrascensis form one group, with no clear-cut subdivisions, Capacikala being the smallest (shorter and less wide) and "Capatanka" the largest (Figure 6.1). Teeth of Palaeocastor sp. overlap with both other taxa. The one measurement for "Capatanka" magnus falls well within the size range of "C". cankpeopi. All length x width data of p4 are parallel to the 45° line, indicating p4s are only slightly longer than wide. The p4 of all taxa increase in length and width with wear. Mandibular p4 of different species of Palaeocastor are difficult to differentiate on the basis of the size of teeth (Figure 6.2); even Pseudopalaeocastor barbouri falls within this group. Just one strongly worn tooth of Palaeocastor sp. is markedly broader (LACM 9305).
The lower molars, separated as m1,2 and m3, also fall within one group parallel but slightly above the 45° degree line, indicating a general near square outline being slightly wider than long with wear. More lower molars than premolars are available, but not of similar wear stages.
 Therefore, medium to strongly worn m1/2 were compared (Figure 6.3). Teeth of "Capatanka" cankpeoi and Capacikala gradatus fall in separate groups, but teeth of Palaeocastor sp. overlap with both (for clarity not shown in the diagram). Even though in general medium worn teeth of Capacikala gradatus are larger than little worn ones, there is considerable overlap of small medium worn teeth and little worn ones, which should not occur in one species. According to size of m1/2 (irrespectively of wear) Palaeocastor sp., P. nebrascensis, P. peninsulatus, P. fossor and Pseudopalaeocastor barbouri cannot be separated (Figure 6.4). Looking at m3 of these taxa there seem to be three size groups, two of Palaeocastor sp. and P. nebrascensis and P. fossor together with larger teeth, but less material of these is available (Figure 6.5). In all studied taxa m3 are slightly smaller than m1/2, and morphometric changes in the molars with wear are small. They increase slightly in length and width in Capacikala, increase slightly more in width than in length in "Capatanka" and Palaeocastor.
Therefore, medium to strongly worn m1/2 were compared (Figure 6.3). Teeth of "Capatanka" cankpeoi and Capacikala gradatus fall in separate groups, but teeth of Palaeocastor sp. overlap with both (for clarity not shown in the diagram). Even though in general medium worn teeth of Capacikala gradatus are larger than little worn ones, there is considerable overlap of small medium worn teeth and little worn ones, which should not occur in one species. According to size of m1/2 (irrespectively of wear) Palaeocastor sp., P. nebrascensis, P. peninsulatus, P. fossor and Pseudopalaeocastor barbouri cannot be separated (Figure 6.4). Looking at m3 of these taxa there seem to be three size groups, two of Palaeocastor sp. and P. nebrascensis and P. fossor together with larger teeth, but less material of these is available (Figure 6.5). In all studied taxa m3 are slightly smaller than m1/2, and morphometric changes in the molars with wear are small. They increase slightly in length and width in Capacikala, increase slightly more in width than in length in "Capatanka" and Palaeocastor.
The upper premolars, mostly medium to strongly worn ones, are generally slightly wider than long. Especially those of "Capatanka" cankpeopi vary considerably in width (Figure 7.1). The one measurement for "C." magnus is larger than for "C". cankpeopi. Capacikala parvus falls well within the size range of C. gradatus here. Length x width data for M1/2 indicate a large variation in size for Capacikala gradatus.
M3 is slightly shorter and less wide than M1/2. Looking at all M1/2 irrespective of wear stage, "Capatanka" cankpeopi is slightly larger, than Capacikala gradatus, C. parvus falls below the range of C. gradatus considering the two data overlapping directly with "Capatanka" minor are not really Capacikala (Figure 7.2). M3 of "Capatanka" minor is clearly smaller than the M3 of Capacikala parvus.
Looking at the upper molars on the basis of wear stages, "Capatanka" minor is smaller than "C". cankpeopi, Capacikala gradatus and Palaeocastor nebrascensis in strongly worn teeth (Figure 7.3), but the medium worn M1/2 fall well within P. nebrascensis. Medium worn teeth of Capacikala parvus are smaller than little worn teeth of C. gradatus but the samples are small.
Discriminant Analysis with Wear Independent Residuals
 The descriptive statistics of the unstandardized residuals based on the tooth measurements are given in
the
Appendix. Using these age-independent residuals either for upper or lower teeth in DFAs shows that separation of the taxa is possible – to some degree.
The descriptive statistics of the unstandardized residuals based on the tooth measurements are given in
the
Appendix. Using these age-independent residuals either for upper or lower teeth in DFAs shows that separation of the taxa is possible – to some degree.
In a DFA based on the maxillary cheek teeth of the taxa with larger samples, Capacikala gradatus, "Capatanka" cankpeopi and Palaeocastor nebrascensis these seem to separate well, but there is no statistical significance for the functions (Figure 8). Using mandibular teeth in the DFA (Figure 9)
 the means of these three taxa are well separated along function 1 (mainly influenced by the residual based on length of p4), is statistically significant, but there are lots of non-grouped cases, including all available taxa (quite different in sample sizes) in the DFA with residuals of maxillary teeth (Figure 10). Pseudopalaeocastor barbouri and Capacikala gradatus separate well from the others. Using residuals based on lower teeth (Figure 11) P. nebrascensis separates strongly from the others along function 2 mainly based on the residual of m1, 2 width.
the means of these three taxa are well separated along function 1 (mainly influenced by the residual based on length of p4), is statistically significant, but there are lots of non-grouped cases, including all available taxa (quite different in sample sizes) in the DFA with residuals of maxillary teeth (Figure 10). Pseudopalaeocastor barbouri and Capacikala gradatus separate well from the others. Using residuals based on lower teeth (Figure 11) P. nebrascensis separates strongly from the others along function 2 mainly based on the residual of m1, 2 width.
 A DFA with different species originally assigned to Palaeocastor and residuals based on mandibular tooth measurements (not shown) indicates separation of P. nebrascnesis, and closeness of P. peninsulatus, P. fossor and Pseudopalaeocastor barbouri based on mandibular teeth. But there is no statistical significance for the functions.
A DFA with different species originally assigned to Palaeocastor and residuals based on mandibular tooth measurements (not shown) indicates separation of P. nebrascnesis, and closeness of P. peninsulatus, P. fossor and Pseudopalaeocastor barbouri based on mandibular teeth. But there is no statistical significance for the functions.
 A DFA based on maxillary teeth for the same taxa indicates stronger separation, here of P. nebrascensis along function 2 based mainly on the residual of length of M3 and Pseudopalaeocastor barbouri along function 1 mainly based on the residual of width of P4. In this case functions 1-4 are statistically significant.
A DFA based on maxillary teeth for the same taxa indicates stronger separation, here of P. nebrascensis along function 2 based mainly on the residual of length of M3 and Pseudopalaeocastor barbouri along function 1 mainly based on the residual of width of P4. In this case functions 1-4 are statistically significant.
Tooth Rows
 Few data of tooth row lengths were available and are illustrated comparatively in
Figure 12.2. As with individual teeth "Capatanka" cankpeopi is clearly larger than Capacikala gradatus. The size range for the lower dentitions of the larger samples are about 4 mm, still smaller than in the subhypsodont European Early Miocene beaver Steneofiber eseri
von Meyer 1846. The range for the hypsodont and considerably larger Recent Castor fiber is about 10 mm. Palaeocastor sp. shows the largest range in size of lower dentitions, P. nebrascensis overlaps with "Capatanka" cankpeopi. The picture for maxillary dentitions is similar, but fewer data were available. The large size range indicated for Palaeocastor fossor is based on two measurements only.
Few data of tooth row lengths were available and are illustrated comparatively in
Figure 12.2. As with individual teeth "Capatanka" cankpeopi is clearly larger than Capacikala gradatus. The size range for the lower dentitions of the larger samples are about 4 mm, still smaller than in the subhypsodont European Early Miocene beaver Steneofiber eseri
von Meyer 1846. The range for the hypsodont and considerably larger Recent Castor fiber is about 10 mm. Palaeocastor sp. shows the largest range in size of lower dentitions, P. nebrascensis overlaps with "Capatanka" cankpeopi. The picture for maxillary dentitions is similar, but fewer data were available. The large size range indicated for Palaeocastor fossor is based on two measurements only.
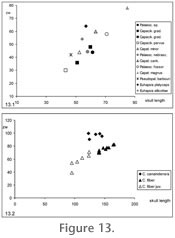 Skulls
Skulls
The morphology of skull material of the considered taxa has been discussed elsewhere (Martin 1987;
Xu 1996), and a relevant description in all detail is beyond the scope of this paper. However, the available data for skull length and zygomatic width are briefly considered in the context of the morphometrics discussion here. Comparing the skull length x width data for some taxa shows that all skulls are nearly as broad at the zygomatic arch as long; this is particularly strong for Euhapsis (Figure 13.1). "Capatanka" magnus is clearly larger than the others. Considering the potential range indicated by two measurements of Capacikala gradatus considerable overlap between the nominated species in the length x width ratios of skulls can be assumed. For comparison, the range of the larger Castor fiber and C. canadensis (Kuhl 1820) are given.