Discussion
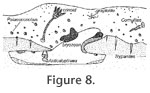 We interpret the environment of the Ohesaare hardground association (Figure 8) as a shallow sea floor of normal salinity below the tidal zone (shelf of moderate depth to shoal). This is supported by the shallow water origin of the
overlying rocks (Nestor 1990) (Figure 2) and the presence of crinoids. The hardground was probably cemented under the sediment cover and exposed later to do winnowing, which also exposed the cryptic surfaces. The abraded bioclasts in the hardground indicate that the hardground was abraded before the encrustation. The sediment layer below the hardground was probably still soft or less cemented than the hardground during the abrasion. We interpret the environment of the Ohesaare hardground association (Figure 8) as a shallow sea floor of normal salinity below the tidal zone (shelf of moderate depth to shoal). This is supported by the shallow water origin of the
overlying rocks (Nestor 1990) (Figure 2) and the presence of crinoids. The hardground was probably cemented under the sediment cover and exposed later to do winnowing, which also exposed the cryptic surfaces. The abraded bioclasts in the hardground indicate that the hardground was abraded before the encrustation. The sediment layer below the hardground was probably still soft or less cemented than the hardground during the abrasion.
The high bioerosion density for the Silurian (Tapanila et al. 2004) (Figure 4.1) presumably indicates a relatively long exposure time for the hardground, before it was buried by clayey sediments (overlying marl interlayer).
Tapanila et al. (2004) found that bioerosion of Ordovician-Silurian corals and stromatoporoids was a widespread process, but that high boring density was generally rare. The situation in Ohesaare hardground is slightly different. Usual 20 to 180 borings per 5 cm2 of hardground upper surface corresponds roughly to high boring density by
Tapanila et al. (2004) (Figure 4.1). However, samples studied by
Tapanila et al. (2004) are stratigraphically older (Hirnantian to Telychian), so high boring densities in Ohesaare could reflect an evolutionary trend. They also may reflect simply a longer exposure time of the substrate. The long exposure time is more likely as the major increase in bioerosion intensity took place earlier in the Middle Ordovician (Wilson and Palmer 2006). Alternatively, the high boring densities can be interpreted as reflecting an environmental or biogeographic difference.
The relatively low skeletal coverage of the hardground (Figure 4) as compared to the other Ordovician to Devonian analogues (Brett and Liddell 1978) can be explained by low productivity (low nutrient levels) in the sea water (Lescinsky et al. 2002) or by a large number of soft-bodied organisms in the community, which did not preserve as fossils.
Lescinsky et al. (2002) found that modern bioerosion is higher at more productive sites in the ocean. They also found that more productive sites have higher encrustation rates. The Ohesaare hardground has a relatively low encrustation density (Segars and Liddell 1988,
Lebold 2000) and a relatively high bioerosion density for the Silurian (Tapanila et al. (2004)) (Figure 4). Thus, if these relations were valid in the late Silurian, it is possible that much of the substrate was occupied by biofilms (preventing attachment of several skeletal encrusters) and soft-bodied encrusters (competing with skeletal ones) as compared to typical Silurian hard substrate communities.
The lower skeletal coverage of the upper surfaces as compared to the cryptic surfaces is due to distribution of sheet-like trepostome bryozoans (Table 2), which cover five times more area on cryptic surfaces than they do on upper surfaces (Figure
4, Figure 6, and
Figure 7). Such strong preference of sheet-like bryozoans for the cryptic niche could be explained by higher predation pressure of grazers on the upper surface. However, the upper surface may also have been more heavily covered by biofilms or soft-bodied encrusters not tolerated by bryozoans.
The larger number of skeletal overgrowths in the cryptic community as compared to the upper surface community can be explained by the higher total skeletal coverage of the cryptic surface. This could also reflect the higher spatial competition between the skeletal encrusters in the cryptic community. However, none of the five overgrowths shows signs that overgrowth of one encruster by another led to the death of the first encruster. Cryptic microconchids were able to stay alive when overgrown by bryozoans by keeping their slightly erected apertures free of bryozoans (Figure 7).
The patchy distribution of Trypanites borings and microconchids on the hardground can be explained by gregarious larval settling behavior or by space being preoccupied by soft-bodied organisms or by uneven survival of larva because of differences in microenvironmental conditions (Figure
4 and Figure 8 8). The organisms that produced Trypanites borings preferentially drilled the high points on hard substrates in the Ordovician, probably for better filter-feeding (Brett and Liddell 1978;
Brett and Brookfield 1984;
Bodenbender et al. 1989;
Wilson and Palmer 1992). This appears to have been the case as well for the Ohesaare hardground.
The Ohesaare hardground fauna is unique for its numerical dominance by Palaeoconchus (Figure 6). Their high abundance in the community (Table 1), large total area covered and high percentage of relative skeletal cover (Table 2) are unmatched elsewhere in the Silurian (Segars and Liddell 1988;
Lebold 2000). Why did the Palaeoconchus dominate this association? They presumably were opportunistic suspension feeding encrusters, successful in conditions of low competition with other suspension feeders. The low total coverage by bryozoans and lack of stromatoporoids and favositids could have enhanced the abundance of Palaeoconchus in the community. The unusually high number of Palaeoconchus on the Ohesaare hardground compared to other Silurian hard substrate communities (Segars and Liddell 1988;
Lebold 2000) presumably also reflects an evolutionary trend. Microconchids are more abundant in Devonian than in Ordovician-Silurian hard substrate communities (Taylor and Wilson 2003). This association is the only hardground fauna known from the Pridoli. Thus, the abundance of microconchids in hard substrate communities may have increased by the Pridoli to the typical Devonian level. Alternatively, the high abundance of Palaeoconchus could have been caused by the local environmental factors, such as low abundance of the other sclerobionts. Crinoids are usually a very important component of Silurian hardground communities (Halleck 1973;
Franzén 1977). The Ohesaare hardground is not typical for the Silurian because of its low abundance of crinoids (Figure 8). Hederelloids, which are very characteristic of Devonian sclerobiont communities (Taylor and Wilson 2003), are lacking in the Ohesaare hardground fauna. The lack of hederelloids may be due to their later evolutionary diversification in the Devonian (Taylor and Wilson 2008).
It is most intriguing why stromatoporoids and favositids are absent from the Ohessaare hardground (Figure 8). They constitute an important component of other Silurian to Devonian hardground communities (Taylor and Wilson 2003). Stromatoporoids are not known from the Ohessaare cliff, but some favositids such as Favosites forbesi, F. pseudoforbesi and F. vectorius are common in the section. In addition to tabulates, rugose corals also occur in the Ohessaare cliff (Mõtus and Hints 2007). Possible explanations include the presence of the biofilms preventing settlement of stromatoporoid and favositid larvae, or too much suspended sediment for stromatoporoids and favositids. However, the hardground environment would likely not have had much sedimentation because it is by definition one where deposition rates are very low. Thus, biofilms seem to be the most realistic explanation for the lack of stromatoporoids and favositids. In addition, there may have been a lot of sediment in suspension, creating turbid and/or abrasive conditions.
Palaeoconchus preference for hardground upper surfaces (Figure 4 and
Figure 8) could be due to higher concentration of nutrients available in the currents or weaker feeding and spatial competition with the other suspension feeders such as bryozoans (e.g., lower coverage by skeletal encrusters of the upper surface). Our observations are in agreement with the results of previous studies of microconchids on Silurian stromatoporoids. Microconchids are more common on stromatoporoid upper surfaces than on their cryptic surfaces (Segars and Liddell 1988;
Lebold 2000). Thus, the preference for upper surfaces of hard substrates may be a general feature of Silurian microconchids. In contrast, the phylogenetically close Anticalyptraea (Vinn and Isakar 2007) preferred cryptic surfaces to the upper surfaces of the hardground (Figure 6 and
Figure 8). Anticalyptraea grew to a much larger size than Palaeoconchus and could have been more attractive for the predators, including durophagous predators. This is supported by the occurrence of shell repair presumably resulting from attempted predation in Anticalyptraea from the Pridoli of Estonia (Vinn and Isakar 2007). Thus, Anticalyptraea's preference for the cryptic surface of the hardground could be due to predation pressure.
Future work needs to be done on Silurian hardground faunas in order to find whether the high abundance of microconchids in the Ohesaare is a local phenomenon or characteristic of the late Silurian hardground associations. In addition, the palaeoecology of Anticalyptraea needs further study, especially regarding the predation and probable preference of cryptic environments.
|