| |
Introduction
Carbonate hardgrounds are surfaces of synsedimentarily cemented carbonate layers that have been exposed on the seafloor. Carbonate hardgrounds are common in calcite sea conditions because of favorable conditions for early cementation of carbonate sediments in the seafloor (Wilson and Palmer 1992). Hardgrounds form suitable attachment surfaces for encrusting and bioeroding organisms. There was a calcite sea in the Silurian, and hardgrounds were common, though probably less abundant than in the Ordovician (Taylor and Wilson 2003). There are only four detailed studies of Silurian hardground communities:
Halleck (1973),
Franzén (1977),
Cherns (1980), and
Sumrall et al. (2009). However, no hardground fauna has been described from the latest Silurian (Pridoli).
Silurian hardground faunas are in general similar to those of Ordovician. They are dominated by bryozoans and echinoderms, particularly crinoids. Devonian encrusting communities are better known on shells than other hard substrates (Taylor and Wilson 2003). These communities are dominated by microconchids, hederelloids and tabulate corals instead of bryozoans and echinoderms, although the latter two groups are still common (Kesling et al. 1980;
Sparks et al. 1980;
Bonem 1982;
Brett and Cottrell 1982;
Alvarez and Taylor 1987;
Bordeaux and Brett 1990;
Brice and Mistiaen 1992;
Grimm 1998;
Taylor and Wilson 2003). Especially interesting is the question of how and when typical Ordovician-Silurian sclerobiont communities were replaced by typical Devonian ones.
Hederelloids are extinct colonial animals with calcitic tubular branching exoskeletons, and they appear to be most closely related to phoronids. Tentaculitoid tubeworms could also be phylogenetically closely related to the hederelloids (Taylor and Wilson 2008).
Small, spirally-coiled calcareous worm tubes are common on Paleozoic and Triassic hard substrates (Taylor and Vinn 2006). Such tubeworms have been traditionally assigned to the polychaete genus Spirorbis. However, pre-Cretaceous examples have been reinterpreted as microconchids (Class Tentaculita
Bouček 1964; Order Microconchida
Weedon 1991) on the basis of the early ontogeny and microstructure of their tubes (Weedon 1991,
1994;
Dreesen and Jux 1995;
Taylor and Vinn 2006;
Vinn and Taylor 2007). Two genera of microconchids are known from the Silurian (Vinn 2006a). Several other groups of problematical calcareous tubeworms referred to the Tentaculita
Bouček, 1964, occur in the Paleozoic (Vinn and Mutvei 2005,
2009;
Vinn 2010;
Taylor et al. in press). They include Tentaculitida
Ljaschenko, 1955, Cornulitida
Bouček, 1964, and Trypanoporida
Weedon, 1991, as well as the genera Anticalyptraea (Vinn and Isakar 2007) and Tymbochoos (Vinn 2006b). Endosymbiotic Streptindytes in the Middle Devonian rugose corals and stromatoporoids may also be a tentaculitoid (Vinn and Mõtus 2008).
In this paper, the researchers hope to: 1) describe for the first time a hardground association of Pridoli age; 2) compare the hardground fauna from the Pridoli of Saaremaa to other Silurian and Devonian analogues; 3) contrast upper surface and cryptic communities; 4) discuss the paleoecology of tentaculitid tubeworms; and 5) discern how gradually typical Ordovician-Silurian hardground communities were replaced by typical Devonian communities.
Locality and stratigraphy
 The Ohesaare cliff (58°0'2" N, 22°1'10" E) is located on the western coast of the Sõrve Peninsula (Saaremaa, Estonia) near Ohesaare village (Figure 1). The cliff is over 600 m long and up to 4 m high (c and
Figure 3). The total thickness of the exposed bedrock is 3.5 m, whereas the thicknesses of individual beds are rather variable throughout the outcrop (Hints 2008). The section is characterized by the intercalation of thin-bedded limestones and marlstones (Figure 2 and
Figure 3). The Ohesaare cliff (58°0'2" N, 22°1'10" E) is located on the western coast of the Sõrve Peninsula (Saaremaa, Estonia) near Ohesaare village (Figure 1). The cliff is over 600 m long and up to 4 m high (c and
Figure 3). The total thickness of the exposed bedrock is 3.5 m, whereas the thicknesses of individual beds are rather variable throughout the outcrop (Hints 2008). The section is characterized by the intercalation of thin-bedded limestones and marlstones (Figure 2 and
Figure 3).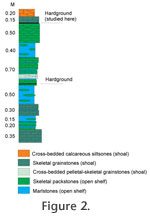 There are two hardgrounds in the section, one in the upper part (studied here) and another in the lower middle part of the cliff (Figure 2 and
Figure 3). The studied hardground is underlain by 0.5 m of skeletal packstone and overlain by 0.15 m silty skeletal grainstone, the upper surface of which bears large ripple marks (Hints 2008). The rocks of Ohesaare section have a high content of terrigenous material. The intense influx of fine siliciclastic material into the basin possibly took place at the final stage of its development (Mõtus and Hints 2007). The rocks of the Ohesaare cliff correspond to the Monograptus transgrediens biozone, late Pridoli, Ohesaare Stage (Hints 2008). There are two hardgrounds in the section, one in the upper part (studied here) and another in the lower middle part of the cliff (Figure 2 and
Figure 3). The studied hardground is underlain by 0.5 m of skeletal packstone and overlain by 0.15 m silty skeletal grainstone, the upper surface of which bears large ripple marks (Hints 2008). The rocks of Ohesaare section have a high content of terrigenous material. The intense influx of fine siliciclastic material into the basin possibly took place at the final stage of its development (Mõtus and Hints 2007). The rocks of the Ohesaare cliff correspond to the Monograptus transgrediens biozone, late Pridoli, Ohesaare Stage (Hints 2008).
 During the Silurian the Baltica continent was located in equatorial latitudes drifting northwards (Melchin et al. 2004). The pericontinental Baltic paleobasin in modern Estonia was characterized by a wide range of tropical environments and diverse biotas (Hints 2008). According to the model worked out by
Nestor and Einasto (1977), five main facies belts can be differentiated in the Baltic Silurian Basin: tidal flat/lagoonal, shoal, open shelf, transitional (basin slope), and a basin depression. The first three facies belts formed a carbonate shelf or carbonate platform and the latter two a deeper pericratonic basin with fine-grained clastic deposits (Raukas and Teedumäe 1997). The rocks of the Ohesaare cliff were formed in shoal and open shelf
conditions. During the Silurian the Baltica continent was located in equatorial latitudes drifting northwards (Melchin et al. 2004). The pericontinental Baltic paleobasin in modern Estonia was characterized by a wide range of tropical environments and diverse biotas (Hints 2008). According to the model worked out by
Nestor and Einasto (1977), five main facies belts can be differentiated in the Baltic Silurian Basin: tidal flat/lagoonal, shoal, open shelf, transitional (basin slope), and a basin depression. The first three facies belts formed a carbonate shelf or carbonate platform and the latter two a deeper pericratonic basin with fine-grained clastic deposits (Raukas and Teedumäe 1997). The rocks of the Ohesaare cliff were formed in shoal and open shelf
conditions.
The Ohesaare cliff has a rich and diverse shelly fauna. The most abundant macrofossils are brachiopods, represented by Delthyris magna
Kozlowsky, 1929, D. elevata
Dalman, 1928, Homoeospira baylei (Davidson 1848), Morinorhynchus orbignyi (Davidson 1848), Isorthis ovalis (Paškevičius 1962), Dalejina hybrida (Sowerby 1839), and Shaleria (Janiomya) ornatella (Davidson 1871). Bryozoans are relatively numerous compared to other eastern Baltic Silurian sections; they include: Fistulipora tenuilamellata (Bassler 1911), F. aculeata
Astrova, 1960 and Eridotrypa parvulipora
Ulrich and Bassler 1913. Bivalves are represented by Grammysia obliqua (McCoy 1852), Cardiola interrupta
Sowerby, 1839, Palaeopecten danbyi (McCoy 1851), and Modiolopsis complanata
Sowerby, 1839. Trilobites are also common. Corals occur at certain levels in the middle part of section. The middle part of the section has also yielded the tentaculitids Tentaculites scalaris (Schlotheim 1820) and Lowchidium inaequale
Eichwald 1860. The microfossil association is very diverse and rich, especially the ostracodes (Mõtus and Hints 2007).
|