DISCUSSION
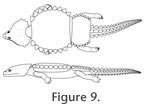 The present study shows that the basic shape of the dorsal armour of the cyamodontoid Cyamodus hildegardis is much narrower than previously reconstructed, with the main carapace having the length-width ratios comparable to the new Chinese cyamodontoids, instead of that of the well-known European forms Psephoderma alpinum and Henodus chelyops. C. hildegardis was thus a medium-sized cyamodontoid placodont (Figure 9), which had a rather round carapace and a separate smaller ovoid and laterally expanded pelvic shield with straight to slightly concave anterior and posterior margins that mainly covered the pelvic region and the base of the tail. The rather short tail itself was armoured with prominent single dorsal and paired lateral series, as well as a more flattened ventral series of dermal plates, all becoming reduced in size in an anteroposterior gradient. Both the anterior shields carrying few enlarged spiked armour plates as well as the armoured tail lend this species a well-fortified character at least as adults. Without new fossil specimens, however, the internal structures and arrangement of the dermal armour of C. hildegardis still remain poorly known to date.
The present study shows that the basic shape of the dorsal armour of the cyamodontoid Cyamodus hildegardis is much narrower than previously reconstructed, with the main carapace having the length-width ratios comparable to the new Chinese cyamodontoids, instead of that of the well-known European forms Psephoderma alpinum and Henodus chelyops. C. hildegardis was thus a medium-sized cyamodontoid placodont (Figure 9), which had a rather round carapace and a separate smaller ovoid and laterally expanded pelvic shield with straight to slightly concave anterior and posterior margins that mainly covered the pelvic region and the base of the tail. The rather short tail itself was armoured with prominent single dorsal and paired lateral series, as well as a more flattened ventral series of dermal plates, all becoming reduced in size in an anteroposterior gradient. Both the anterior shields carrying few enlarged spiked armour plates as well as the armoured tail lend this species a well-fortified character at least as adults. Without new fossil specimens, however, the internal structures and arrangement of the dermal armour of C. hildegardis still remain poorly known to date.
Main Carapace
Because of superficial similarities, the armour of Cyamodus hildegardis is often compared to that of Psephoderma alpinum, and reconstructions of the former often refer to the latter, a well-known species from the Upper Triassic. However, based on the new interpretations and carapacial length-width ratios presented herein, P. alpinum might not be such a good reference model for the reconstruction of the postcranium and overall body shape of C. hildegardis, as maybe Glyphoderma kangi and Psephochelys polyosteoderma from China, although these so far lack a separate hip shield. Although several complete or well articulated specimens of P. alpinum are known, the carapace appears dorsoventrally compacted during fossilisation in most of these specimens (von
Meyer 1858: BSP AS I 8;
Pinna and Nosotti 1989: MCSNB 8358;
Renesto and Tintori 1995: ST82003 stored in the Museo della Vicaria di S. Lorenzo, Zogno, Bergamo, Italy), resulting in a much broader appearance of the carapace than in life. In their emended diagnosis for Psephoderma,
Renesto and Tintori (1995: p.39) stated the main carapace to be "rounded, stout, wide and very flat [...]". Because only the length of 420 mm and not the carapace width was given for specimen ST82003 in that paper, it is not listed in
Table 1. Other specimens, however (e.g.,
Pinna and Nosotti 1989: MSNM V527; undescribed specimen in PIMUZ, H. Furrer, personal commun.), show much less lateral extension of the carapace, resulting in a more anteroposteriorly elongated oval body shape (Pinna and Nosotti 1989, pl. XXVI, XXVII). This is also underlined by the well articulated gastral apparatus in MSNM V527, indicating that the carapace was not broadly expanded. It cannot be ruled out, however, that intraspecific, ontogenetic, or sexual dimorphic variation instead of (or in addition to) taphonomic compaction is at least partly responsible for the different carapace shapes in these specimens as well. Until new material is discovered that clearly indicates the association of the main dermal armour and underlying skeletal elements, it is hypothesized that the three dimensionally preserved specimens like MSNM V527 are better suited for skeletal reconstruction in P. alpinum. In contrast to the condition seen in P. alpinum (Pinna and Nosotti 1989), the main outline of the carapace in C. hildegardis is interpreted to be round to slightly ovoid.
For the schematic reconstruction of C. hildegardis, though, a fairly broad carapace was previously hypothesized with a length-width ratio of about 0.7 (Pinna 1992, figure 19); a value that is lower even than that for H. chelyops (Table
1). Furthermore, based on these differences in the development and orientation of armour, it cannot be implied that the same endoskeletal regions are covered by armour in C. hildegardis and P. alpinum (Pinna and Nosotti 1989: posterior cervicals to sacral 1).
Pelvic Shield
The pelvic shield is situated posterior to the carapace in Cyamodus hildegardis, but again it is not sure whether the same endoskeletal regions are covered as in Psephoderma alpinum (Pinna and Nosotti 1989: sacral 3 to caudal 8). In C. hildegardis, at least part of the sacrum is covered by the pelvic shield, but as the shield is much narrower, it cannot cover the proximal tail region back to the eighth caudal.
Pinna (1992, p.13) assumes that "[...] la seconda vertebra caudale occupi lo spazio non corazzato compreso fra questi due margini," meaning the second caudal vertebra occupies the non-armoured space comprised between the posterior edge of the carapace and the anterior edge of the pelvic
shield. This assumption would indicate, however, that the complete sacrum is covered by the main carapace, and only the proximal tail region is covered by the pelvic shield. Assuming that
Pinna (1992) actually meant 'second sacral' instead of 'second caudal', the pelvic shield would then have covered almost the same portions of the sacrum and the tail as reported by
Pinna and Nosotti (1989) for P. alpinum.
Although the articulated vertebral centra and associated fragmentary lateral bones below the pelvic shield cannot be unambiguously identified as sacrals and sacral ribs, it is still plausible to hypothesise that the complete pelvic shield and the associated vertebral elements were dislocated posteriorly, thus slightly opening up the natural gap between the main carapace and the pelvic shield. This
condition is in accordance with previous assessments that fossils of the Monte San Giorgio locality can show signs of disturbance either by light bioturbation, wave action, synsedimentary slumpings or diagenetic pressure compaction of the sediment (Tintori 1992;
Röhl et al. 2001;
Rieppel 2002).
Pectoral and Pelvic Girdles
In contrast to the interpretative drawing in
Pinna (1992, figure 1), more elements pertaining to the vertebral column in PIMUZ T58 could be identified associated with the pelvic shield, whereas elements previously identified as belonging to the pelvic and pectoral girdles are either not observable or have been misidentified (Figure 6,
Figure 7,
Figure 8). In PIMUZ T58, only the scapulae are well recognisable, whereas armour
plates of the marginal series of the carapace have been misidentified as
coracoids. The position and the size and shape of the bones both argue against
the presence of coracoids in this matter. Other elements of the pectoral girdle
might be present as well, but they are too poorly preserved to be identified
with confidence.
Although often fragmentary, the ilia and ischia are preserved, but no pubic bone could be identified. It is not clear why the pubes and coracoids, both thin, large flat bones in C. hildegardis (e.g.,
Pinna 1980), are not identified in PIMUZ T58. Hypothetically, because of the flattened shape and ventral position in the animal, these bones could have been more easily disturbed and dislocated than other girdle elements, and thus, might not be preserved on the slab with the rest of the skeleton. Alternatively, due to their thin nature, the bones could be present but are too severely crushed and obscured by overlying or underlying postcranial elements or fragments of armour. Based on the PIMUZ specimens, shape and size of the pubic bones and the coracoids thus cannot be inferred, and
Pinna's (1980) reconstruction of the pelvic and pectoral girdles in the juvenile specimen MSNM V458 has to be consulted instead.
Tail Armour
Pinna (1992) and
Rieppel (2002) agree that the dorsal part of the tail is covered by a median row and two lateral rows of armour plates. However, Rieppel interpreted the elements of the lateral series to be enlarged, whereas Pinna identifies only smaller elements whose positions are not exactly known as belonging to the lateral series. Here
Rieppel's (2002) identification of larger plates in the lateral series is confirmed and Pinna's interpretation is attributed to a misidentification of caudal vertebral centra as lateral armour plates in the proximal part of the tail (Pinna 1992, figure 21: v10-v16).
Following Pinna (1992) the ventral series of the tail was also covered by a row of armour plates, whereas the presence of chevron bones in the holotype PIMUZ T4763 commented upon by
Peyer (1931) could not be confirmed. Chevron bones, if present, would be rather slender elements situated below the posterior edge of the vertebral centra; however, the only bones lying ventral to the caudal centra are the massive posteriorly tapering elements identified as dermal tail armour herein. These plates are positioned below the anterior edges of the vertebral centra, respectively. If hemapophyses are present in the more proximal part of the tail, they are completely obscured by the large overlying armour plates.
Novel Interpretation of the Postcranial Skeleton and Associated Armour
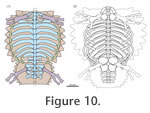 Based on the data and discussion presented herein, a new hypothetical association between the reconstructed skeleton and the dermal armour of C. hildegardis is proposed (Figure 10.1). According to this new interpretation, the round to ovoid main shield covers the main trunk of the animal from about the first to the
1n dorsal. If the posterior margin of the main carapace would be reconstructed with a convex margin instead of a straight or slightly concave one, the main armour might also span onto the 12th dorsal. The width of the main carapace is basically reflected by the length of the transverse processes plus ribs. Measuring the length of the transverse processes plus associated ribs at dorsal 6, the carapace has a maximum width of approximately 215 mm. As mentioned before, the length of the main carapace is more difficult to assess. A first approximation, however, can be given by taking the length of the dorsal vertebral centra associated with the carapace as a proxy. Those centra of PIMUZ T58 preserved in ventral view (dorsals 2-4, 8-9?; sacrals 1-3?, caudal 1?) range between 15 and 22 mm (average 18.5) in length indicating a carapace length (for eleven dorsals) of approximately 205 mm. Although these numbers should be regarded with caution, they overall fit the measurements taken directly at the specimen, leading to similar length-width ratios in both approaches (see
Table 1). The pelvic shield is interpreted to cover partly dorsal 12, the complete sacrum and the base of the tail roughly to caudal 3 (Figure 10.1).
Based on the data and discussion presented herein, a new hypothetical association between the reconstructed skeleton and the dermal armour of C. hildegardis is proposed (Figure 10.1). According to this new interpretation, the round to ovoid main shield covers the main trunk of the animal from about the first to the
1n dorsal. If the posterior margin of the main carapace would be reconstructed with a convex margin instead of a straight or slightly concave one, the main armour might also span onto the 12th dorsal. The width of the main carapace is basically reflected by the length of the transverse processes plus ribs. Measuring the length of the transverse processes plus associated ribs at dorsal 6, the carapace has a maximum width of approximately 215 mm. As mentioned before, the length of the main carapace is more difficult to assess. A first approximation, however, can be given by taking the length of the dorsal vertebral centra associated with the carapace as a proxy. Those centra of PIMUZ T58 preserved in ventral view (dorsals 2-4, 8-9?; sacrals 1-3?, caudal 1?) range between 15 and 22 mm (average 18.5) in length indicating a carapace length (for eleven dorsals) of approximately 205 mm. Although these numbers should be regarded with caution, they overall fit the measurements taken directly at the specimen, leading to similar length-width ratios in both approaches (see
Table 1). The pelvic shield is interpreted to cover partly dorsal 12, the complete sacrum and the base of the tail roughly to caudal 3 (Figure 10.1).
A similar comparison (Figure 10.2) of previous interpretative reconstructions of C. hildegardis based on
Pinna (1980, figure 1: skeletal reconstruction based on the juvenile specimen MSNM V458;
1992, figure 19: schematic reconstruction of overall body shape), yielded no close association between dermal armour and the underlying skeletal elements. By simply overlaying and scaling the drawings to the same size (based on humerus length), however, it became apparent that
Pinna (1992) most probably meant the sacral 2 and not caudal 2 when identifying the position of the gap between the main carapace and pelvic shield. Apart from this, the sketches of the zeugopodial elements were found to be disproportionally large in the skeletal drawing based on the juvenile specimen. It could be that the zeugopodial elements have been misidentified in MSNM V458 by
Pinna (1990), leading to skeletal reconstructions of these elements which are only marginally shorter than the humerus. This observation is further strengthened by the fact that the larger (and heavier) humerus has been dislocated significantly to the posterior trunk region, whereas the supposed (lighter) zeugopodial elements remained close to their natural position in the anterior trunk region.