|
|
|
POSTCRANIAL ANATOMY: OVERVIEW AND NEW INTERPRETATIONSDespite the incomplete or disturbed nature of the specimens, many anatomical details of the postcranium of C. hildegardis have been aptly described by Peyer (1931), Westphal (1975) and Pinna (1980, 1992) and thus do not have to be repeated here. In the following, an overview of the underlying endoskeletal elements (i.e., vertebrae, transverse processes, ribs, and elements of the girdles) and the associated units building the postcranial armour in C. hildegardis is given, while highlighting the features essential to the discussion and the new interpretation of the material. General Overview of the Body Armour
Vertebral Column
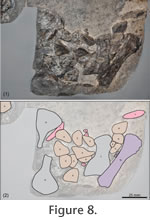
Pectoral and Pelvic GirdlesBased on the juvenile specimen MSNM V458 a reconstruction of both girdles was presented by Pinna (1980). Few girdle bones, although not preserved in articulation, are recognizable in the referred specimen PIMUZ T58. The pectoral and pelvic girdles are not well visible in the holotype specimen PIMUZ T4763 (Peyer 1931). As seen in the juvenile specimen, the pectoral girdle is composed of seven bones: the unpaired clavicle and the paired clavicles, scapulae and coracoids (Pinna 1980). Pinna's (1992) identification of the pectoral bones in specimen PIMUZ T58 is highly schematic, with outlines of individual bones often missing. Although badly preserved, the two scapulae of PIMUZ T58 (Figure 6) are recognisable. I do not concur however with Pinna's (1992) identification of the two coracoids. Instead these elements are interpreted to be enlarged dermal armour plates (Figure 6) from the anterior portion of the main carapace that have been dislocated and in which the visceral surface of the bone is now seen. Because all elements of the pectoral girdle have been dislocated, no further information concerning the relation between these bones and the main carapace is available from this specimen.
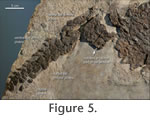
Main CarapaceThe lateral outline of the main carapace can be inferred based on the specimens PIMUZ T58 and MSNM V458, whereas the anterior and posterior margins of the main carapace are not preserved sufficiently in any of the three specimens. Given the length of the transverse processes 1-12 and curvature of the dorsal ribs 3-12 in the juvenile specimen (Pinna 1980, 1992) and the well preserved festooned margin in PIMUZ T58 (Figure 6) consisting of enlarged pointed armour plates, the main carapace shield was round to slightly ovoid in shape (with the anteroposterior axis being only slightly shorter). In this regard, C. hildegardis had a length to width ratio of the carapace of approximately 0.93-0.95 (Table 1) most similar to the Chinese cyamodontoids Psephochelys polyosteoderma (Li and Rieppel 2002a, 2002b) and Glyphoderma kangi (Zhao et al. 2008). Note that these taxa carry only a main carapace shield but no separate pelvic shield. In P. polyosteoderma there are two tapering 'horns' or protrusions at the anterior margin of the carapace, laterally framing a narrow nuchal region and enclosing the pectoral girdle. However, these 'horns' or protrusions are not as strongly developed as the flange-like anterior lobes of the carapace of Henodus chelyops (von Huene 1936, 1938). According to Zhao et al. (2008, figure 3) Glyphoderma kangi lacks tapering 'horns' or flange-like lobes and instead shows only two enlarged osteoderms framing a wide and gently concave nuchal region of the carapace. Already noted by Peyer (1931; see also Rieppel 2002), the external (dorsal) surface of the dermal armour plates in the holotype PIMUZ T4763 has a granular texture and the elements are shallowly pitted (Figure 4, Figure 5). The internal (visceral) surface of the plates, well visible in PIMUZ T58 (Figure 6.1), has a rough texture caused by elevated, often highly regularly arranged cross-hatching mineralized fibres (e.g., Westphal 1975, 1976; Pinna 1992; Rieppel 2002). Pelvic ShieldThe pelvic shield is present in the holotype PIMUZ T4763 and the referred specimen PIMUZ T58. In the holotype, the right lateral margin of the pelvic shield is well preserved, whereas the left margin is partly missing (Figure 4). At the left margin, five enlarged triangular dermal plates are visible. The anterior and posterior margins of the pelvic shield are partly covered and fused to armour plates from the main carapace and plates from the caudal armour respectively, thus the exact borders of the shield are obscured. Within the pelvic shield, it is not possible to identify the delineation between individual osteoderms. As noted already by Peyer (1931), few dermal plates show a flat keel, which is generally oriented in anteroposterior fashion. In PIMUZ T58, the pelvic shield is seen in ventral view, so the main part of the internal organisation of the dermal plates is not visible. However, especially under strongly oblique light conditions (Figure 2.2, Figure 7), the left and right crescent-shaped lateral margins of the pelvic shield are observable. The right part of the shield has been slightly disarticulated from the left part leaving a gap of about 30 mm. The anterior part of the right portion of the pelvic shield is partly hidden beneath four transverse processes and associated vertebrae as well as an isolated rib, which presumably shifted here from the posterior margin of the carapace region. As in the holotype, the margins of the pelvic shield are composed of enlarged triangular dermal armour plates (Figure 7). The left margin shows seven or possibly eight of these lateral plates. By rotating and translocating the right part of the pelvic shield back into its natural position, the anterior and posterior margins of the shield would be straight to very slightly concave (Figure 7). Ventral to the pelvic shield, three articulated and three isolated vertebrae and associated, more or less well preserved transverse processes were identified. A single bone fragment, presumably from a caudal centrum, lies directly adjacent to the posterior margin of the pelvic shield. The isolated pubis and ischium that according to Pinna (1992, figure 1) should be at least partly covered by the posterior margin of the pelvic shield, could not be identified here. Caudal ArmourThe caudal armour is best preserved in the holotype PIMUZ T4763. Only the posterior-most part of the tail and the last vertebrae and associated armour plates of the tip of the tail, and according to Peyer (1931) no more than three distal caudals, are missing. The tail is angled sharply towards the right, thus losing connection with the pelvic shield. A single squared isolated fragment of caudal armour and possibly associated underlying proximal caudals lies between the main part of the tail and the pelvic shield (Figure 5). Although no bone contact exists, the exact position of this armour piece is given by the exactly fitting sediment matrix (Peyer 1931). The tail armour plates generally become reduced in size in an anteroposterior gradient. Because the tail of the specimen is inclined to the right side, its armour was embedded in left lateral view. On the right side of the caudal vertebrae, a dorsal row of posteriorly tapering armour plates is visible. Laterally, the tail was covered on its left and right side by a single row of tubercular armour plates, similar in size to the dorsal series. At the proximal part of the tail, the caudals are largely covered by the lateral row of dermal plates; only in the distal part of the tail the lateral plates have been dislocated enough to reveal the underlying caudal vertebrae exposed in left lateral view. Ventrally, the tail was armoured by an unpaired series of more tapering plates whose apices were sharply inclined towards posterior. Hemapophyses between this ventral series of armour plates and the caudal centra are not visible. Individual positions to the respective caudals and/or associated armour plates in the tail cannot be assigned with accuracy, because only the posterior-most caudals are visible, and a clear contact with the pelvic shield is missing. Gastral ApparatusAs indicated by Pinna (1992, figure 2) the gastral apparatus, consisting of two lateral rows and a medial row of rod-like gastral ribs, is best preserved in the juvenile specimen MSNM V458. In PIMUZ T58, the other specimen exposing the ventral side, the arrangement of gastral ribs is largely disturbed. Pinna's (1992) assessment that the open v-shaped gastralia in PIMUZ T58 belong to the medial row is correct, whereas the only slightly recurved gastralia pertain to the lateral series. |
|