|
|
|
SYSTEMATIC PALEONTOLOGY
Order CAUDATA Oppel, 1810 Family AMBYSTOMATIDAE Hallowell, 1856Genus AMBYSTOMA Tschudi, 1838 The family Ambystomatidae is comprised of one genus, Ambystoma, and approximately 32 species (AmphibiaWeb 2011). Ambystomatids are restricted to North America, being found from Alaska to Mexico, and most are moderately large terrestrial salamanders with lungs (Petranka 1998). Terrestrial species are generally pond breeders, laying their eggs in winter or early spring (Petranka 1998; Wells 2007), then residing in leaf litter or burrows not far from the breeding pond for the remainder of the year (Petranka 1998). Neoteny is expressed in several species and populations of ambystomatids that are wholly aquatic; this is especially common at high altitude or in arid regions where the terrestrial environment would be inhospitable to metamorphosed adults (Wells 2007). Neotenic populations also occur in temperate, low altitude regions with permanent bodies of water lacking large-bodied predatory fish (Petranka 1998; Lannoo 2005; Wells 2007). Vertebral MorphologyVertebrae of Ambystoma are amphicoelous and have spinal nerve foramina posterior to the transverse processes in all vertebrae except the first trunk vertebra (T1) (Edwards 1976; Miller 1992). The atlas has paired atlantal cotyles and a non-faceted odontoid process. T1 has widely separated and robust transverse processes, whereas the other trunk vertebrae have less robust transverse processes where the diapophyses and parapophyses are fused for the majority of their lengths. As noted by Tihen (1958) and Miller (1992), the anterior trunk vertebrae (T1-T3) are proportionally shorter and broader, with steeper neural arches and proportionally larger neural canals. Other trunk vertebrae have lower neural arches and are longer. In general the hyperapophyses are high and have a vertical posterior face (Olori and Bell 2007). The last trunk vertebra (TL) has completely fused transverse processes like the two caudosacral (CS) vertebrae, except that the transverse processes are directed dorsally rather than ventrally (Miller 1992). For both the TL and CS the transverse processes are acuminate. In general Ambystoma tend to have inter-prezygapophyseal neural arch margins that are V-shaped or that are deeply convex, except for the T1 which just as often has a linear inter-prezygapophyseal neural arch margin (Olori and Bell 2007).
Ambystoma sp.
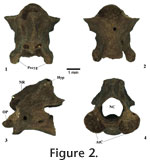
RemarksRobust amphicoelous vertebrae with large transverse processes, V-shaped inter-prezygapophyseal neural arch margins, and notably elevated posterior portion of the neural arch are referred to Ambystoma sp.
First trunk vertebra (T1). Posterior and anterior cotyles are circular. Neural canal is large and constricted dorsally, the canal openings are nearly triangular. The anterior half of the neural arch is horizontal (i.e., flat) with a slightly raised neural crest, whereas the posterior half is rising about 45 degrees to meet a broad hyperapophysis that does not extend beyond the posterior margin of the postzygapophyses. The inter-prezygapophyseal neural arch margin is linear. Postzygapophyseal articular facets are oval-shaped and horizontally oriented; the prezygapophyseal articular facets are also oval-shaped but are slightly elevated posteriorly. Transverse processes are robust, the parapophyses originating in front of the middle of the centrum, the diapophyses in the middle of the neural arch wall; both processes project posteriorly. Large vascular foramina are present in the base of the parapophyses; there are no spinal nerve foramina.
Mid-Trunk vertebrae (TM). These vertebrae have anterior and posterior cotyles that are circular and many have basapophyses. The neural canal is roughly circular and becomes more flattened in the posterior vertebrae.
Sacral (SA). These vertebrae are elongate and have a lower profile than most of the other vertebrae (except TL) that precede it. Posterior and anterior cotyles are circular. The neural canal is flattened, and the neural arch is relatively flat with a nearly obsolete neural ridge. The posterior half of the neural arch rises less than 45 degrees to meet the hyperapophysis, that extends beyond the posterior margin of the postzygapophyses. The inter-prezygapophyseal neural arch margins are generally V-shaped.
Caudosacral vertebra (CS). This vertebra is similar in character to the last trunk vertebra except in that the transverse processes are postero-ventrally projected. Family PLETHODONTIDAE Gray, 1850The salamander family Plethodontidae is the most diverse of the Order Caudata, distributed throughout the Americas and with sparse populations in southern Europe and eastern Asia (Duellman and Trueb 1994; Min et al. 2005). The family is broken up into four subfamilies: Plethodontinae, Spelerpinae, Hemidactylinae, and Bolitoglossinae (Chippindale et al. 2004). Though the family has adapted to a wide array of habitats (arboreal, aquatic, terrestrial, and fossorial) and in general produce direct developing young, they are still restricted to moist environments because they do not have lungs and rely on cutaneous respiration (Wells 2007). Vertebral MorphologyPlethodontid atlantes have odontoid processes with paired articular facets, unlike in Ambystoma (Miller 1992). Vertebrae of plethodontids are amphicoelous (though in some genera they are opisthocoelous) and have spinal nerve foramina posterior to the transverse processes in all vertebrae except the first trunk vertebra (T1), which has spinal nerve foramina anterior to the transverse processes (Miller 1992). The second trunk vertebra (T2) has spinal nerve foramina both posterior and anterior to the transverse processes (Edwards 1976; Miller 1992). With regards to the rest of the trunk vertebrae the first three (T1-T3) have transverse processes that are more posteriorly directed, have proportionally shorter centra, and have larger neural canals (Miller 1992). The remaining trunk vertebrae, except the last trunk vertebra (TL), are very similar to one another. The TL has diapophyses that are reduced, acuminate (Miller 1992), and either have a single very large spinal nerve foramen (as in members of the subfamily Plethodontinae) or two spinal nerve foramina (as in most members of the subfamily Spelerpinae). The sacrum has more robust transverse processes that are united for much of their length (Miller 1992). The caudosacrals have transverse processes that are completely fused and acuminate, and often have alar processes (Miller 1992). In general plethodontids tend to have linear to slightly concave inter-prezygapophyseal neural arch margins, a character used by Olori and Bell (2007) to distinguish plethodontids from some of the more gracile ambystomatids. Subfamily PLETHODONTINAE Gray, 1850 The plethodontid subfamily Plethodontinae (sensu Chippingdale et al. [2004]; Macey [2005]; Min et al. [2005]) is comprised of Aneides, Ensatina, Hydromantes, Karsenia, and Plethodon + Desmognathus.
Plethodon sp. Referred specimens. 5 atlantes (ETMNH 1864, 3847, 4920, 4921), 3 first trunk vertebrae (ETMNH 4922, 4923, 4926), 5 second trunk vertebrae (ETMNH 1865, 4943, 3850, 3851, 3846), 1 third trunk vertebra (ETMNH 7260), 62 mid-trunk vertebrae (ETMNH 7239-7241, 7243, 7244, 7246-7249, 7251, 7252, 7254-7259, 7261-7274, 7276, 7277, 4932-4934, 4929, 4925, 4927, 3863, 1866,1859, 1862), 3 last trunk vertebrae (ETMNH 7242, 7245, 7275), 3 caudosacral vertebrae (ETMNH 1861, 7271, 7253). NIE = 82 and MNI = 5 (based on the number of atlantes). Remarks
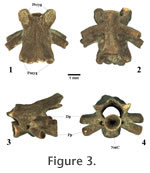
Atlas (AT). The Plethodon sp. atlantes come in two forms: Type A and Type B. Both atlantal morphotypes have odontoid processes with paired articular facets (which clearly distinguish both from the atlantes of ambystomatids). Type A (the smaller form) has circular atlantal cotyles which are laterally placed, and Type B (the larger form) has sub-circular cotyles which are more ventrally placed. Type A has foramina on the ventral surface of the centrum, whereas Type B does not. The neural arch of Type B is thick and tall, and the neural canal is sub-circular. Type A has a thinner, lower neural arch in profile. The posterior cotyle in both morphotypes are circular. None of the atlantal specimens have well-preserved postzygapophyses, so little can be said about their general shape or orientation.
Second trunk vertebra (T2). These trunk vertebrae have the same features as the T1, except that they possess spinal nerve foramina both anterior and posterior to the diapophyses. Third trunk vertebra (T3). This vertebra appears to share the features of the first two trunk vertebrae except that it has a single spinal nerve foramina posterior to the diapophyses. Mid-Trunk vertebrae (TM). These vertebrae are narrow and appear in two morphotypes: Type A and Type B. The anterior and posterior cotyles are circular, with the posterior cotyle being slightly larger and placed more ventrally. The neural canal is wider than tall, and the neural arch is generally flat with a distinct, but medially restricted neural ridge. The posterior portion of the neural arch rises only slightly, ending in paired hyperapophyses that generally do not extend beyond the posterior margin of the postzygapophyses. The prezygapophyseal and postzygapophyseal articular facets are narrow, widening anteriorly, and are slightly laterally inclined. In general the transverse processes are posteriorly projected and relatively short, with the diapophyses being smaller in diameter and placed more posteriorly than the parapophyses. The diapophyses and parapophyses are often connected by a web of bone for part of their lengths. Some of these vertebrae have basapophyses, but most do not. All mid-trunk vertebrae have a single spinal nerve foramina posterior to the diapophyses, and vascular foramina are generally present in the basal portion of the parapophyses. Posteriormost trunk vertebrae (TL). These vertebrae share the same features as the mid-trunk vertebrae except the diapophyses are acuminate, and the parapophyses are connected to the centrum by weakly developed ventral alar processes. Caudosacral vertebrae (CS). These vertebrae are amphicoelous and share most of the features of the last trunk vertebrae except that their transverse processes are completely fused, acuminate, and connected to the centrum by ventral alar processes.
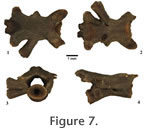
Genus DESMOGNATHUS Baird, 1850 Referred specimens. 1 atlas (ETMNH 1875); 15 mid-trunk vertebrae (ETMNH 1857, 1874, 1883-1886, 1889, 3844, 4930, 4931, 4937); 1 last trunk vertebra (ETMNH 1890); 3 caudosacral vertebrae (ETMNH 1858, 1887, 1888). NIE = 20 and MNI = 2 (based on the number of caudosacrals). RemarksOpisthocoelous vertebrae sharing general features with modern Desmognathus are here referred to Desmognathus sp. Atlas (AT). This fragmentary atlas is missing the entire neural arch and much of the atlantal cotyles, the odontoid process appears worn so it cannot be said if it has articular facets. This specimen is tentatively referred to Desmognathus sp. based on the distinctly ventral location of the posterior cotyle relative to the odontoid process, though it is not as ventrally located as in modern Desmognathus, it is more ventrally located than in other modern plethodontids (and other salamanders). This atlas may represent a transitional form between the generalized plethodontid atlas and the atlas of modern Desmognathus. Mid-Trunk vertebrae (TM). These vertebrae are generally more robust than the Plethodon sp. vertebrae described above. The posterior cotyles are larger than the anterior condyle and more ventrally placed than those of Plethodon sp. and other plethodontids, a morphological arrangement diagnostic to Desmognathus (Holman, 2006). The neural canal and neural arch are similar to the Plethodon sp., but the posterior portion of the neural arch is wider and higher with distinct neural ridges connecting to hypapophyseal ridges, a feature also diagnostic to Desmognathus (Holman, 2006). The preszygapophyseal and postzygapophyseal articular facets are generally oval-shaped and more horizontally oriented. A number of the mid-trunk vertebrae have posterior hypophyses, another feature diagnostic to Desmognathus. The transverse processes are like those in Plethodon sp. A single spinal nerve foramina is present posterior to the diapophyses. Posteriormost trunk and caudosacral vertbrae (TL and CS). These opisthocoelous vertebrae match the overall morphology of Desmognathus. Subfamily SPELERPINAE Cope, 1859 The plethodontid subfamily Spelerpinae (sensu Chippingdale et al. [2004]) is comprised of Gyrinophilus, Stereochilus, Pseudotriton, Eurycea, and Haideotriton. Characters for the plethodontid subfamily Spelerpinae are given as exemplified by Eurycea. Vertebrae generally have widely separated transverse processes (with parapophyses anterior to the diapophyses) that do not exceed the lateral extent of the zygapophyses (Miller 1992; Holman 2006). Vertebrae of many of the extant species examined appear opisthocoelous due to a calcified ring on the anterior cotyle, though in some specimens examined this ring had detached. Conforming with the observations of Edwards (1976), the vertebrae of spelerpine plethodontids (except Gyrinophilus) diagnostically have two spinal nerve foramina exiting posterior to the transverse processes in all trunk vertebrae except T1 (which has two spinal nerve foramina anterior to the transverse processes) and T2 (which has a spinal nerve foramina anterior to and two foramina posterior to the transverse processes).
Spelerpinae, gen. et sp. indet. Referred specimens. 1 first trunk vertebra (ETMNH 1870); 1 third trunk vertebra (ETMNH 1869); 35 mid-trunk vertebrae (ETMNH 1867, 1868, 1891-1900, 7225-7236, 7238); 2 last trunk vertebrae (ETMNH 1871, 7237). NIE = 39 and MNI = 2. Remarks
Family SALAMANDRIDAE Gray, 1825Salamandrids (true salamanders and newts) are primarily found in Europe and Asia, with a few species in North America, Africa, and the Middle East (Wells 2007). Most are semi-aquatic, breed in water, and usually live at least part of their lives on land (Wells 2007). The North American Notophthalmus breed mostly in ponds, larvae transform into a terrestrial 'eft' stage, and then return to the pond several years later to breed and reside as aquatic adults (Wells 2007). The vertebrae of salamandrids are fully opisthocoelous and have spinal nerve foramina exiting posterior to the transverse processes in all vertebrae except the first trunk vertebra (T1) which has spinal nerve foramina exiting both posterior to and anterior to the transverse processes (Edwards 1976; Naylor 1978). Genus NOTOPHTHALMUS Rafinesque, 1820 The vertebrae of Notophthalmus have neural crests and hyperapophyses that are extremely tall and capped by flat, porous bone. Though very similar in morphology to Taricha (the only other salamandrid genus in North America), Notophthalmus has linear to convex inter-prezygapophyseal neural arch margins. In Taricha this margin is concave, which makes distinguishing the two genera very simple. Of the three species of Notophthalmus, N. viridescens is unique in having a highly convex inter-prezygapophyseal neural arch margin (Miller 1992), whereas the margin is linear in the others.
Notophthalmus sp. Referred specimens. 2 trunk vertebrae (ETMNH 3845 and 4919). NIE = 2 and MNI = 1. Remarks
|
|