

 Upper Jurassic sauropod record in the Lusitanian Basin (Portugal): Geographical and lithostratigraphical distribution
Upper Jurassic sauropod record in the Lusitanian Basin (Portugal): Geographical and lithostratigraphical distribution
Article number: 20.2.27A
https://doi.org/10.26879/662
Copyright Society for Vertebrate Paleontology, June 2017
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 21 March 2016. Acceptance: 1 May 2017
{flike id=1865}
ABSTRACT
Sauropod remains are relatively abundant in the Upper Jurassic sediments of the Lusitanian Basin. These dinosaurs are recorded in several sub-basins formed during the third rifting episode related to the evolution of the Lusitanian Basin. The Kimmeridgian-Tithonian sedimentary sequence is dominated by siliciclastic deposits, indicating a continental environment. Sauropods are present all along this mainly terrestrial sequence, being recorded in the Alcobaça, Praia da Amoreira-Porto Novo, Sobral, Freixial, and the Bombarral Formations, ranging from the early Kimmeridgian to the late Tithonian. Sauropoda is the most abundant dinosaur group in the Upper Jurassic fossil record of the Lusitanian Basin and is especially well-represented in the Bombarral and Turcifal Sub-basins. Several new specimens, so far unpublished, are reported here. The sauropod fauna identified mainly includes non-neosauropod eusauropods (including turiasaurs), diplodocoids (some specimens with diplodocine affinities), basal macronarians (non-camarasaurids and camarasaurids), and titanosauriforms (some specimens with brachiosaurid affinities). Macronarians, turiasaurs and diplodocoids are generally present along the entire Kimmeridgian-Tithonian continental to transitional deposits of the Lusitanian Basin, but the known fossil record for some more exclusive groups such as camarasaurids, brachiosaurids, and diplodocines, present a more restricted stratigraphic distribution.
Pedro Mocho. The Dinosaur Institute, Natural History Museum of Los Angeles County, 900 Exposition Blvd., 90007 CA, Los Angeles, USA; Grupo de Biología Evolutiva, Facultad de Ciencias, UNED, c/ Senda del Rey, 9, 28040 Madrid, Spain; Laboratório de Paleontologia e Paleoecologia, Sociedade de História Natural, Polígono Industrial do Alto do Ameal, Pav.H02 e H06, 2565-641, Torres Vedras, Portugal. pmocho@nhm.org
Rafael Royo-Torres. Fundación Conjunto Paleontológico de Teruel-Dinópolis/Museo Aragonés de Paleontología, av. Sagunto s/n. E-44002 Teruel, Spain. royo@dinopolis.com
Fernando Escaso. Grupo de Biología Evolutiva, Facultad de Ciencias, UNED, c/ Senda del Rey, 9, 28040 Madrid, Spain; Laboratório de Paleontologia e Paleoecologia, Sociedade de História Natural, Polígono Industrial do Alto do Ameal, Pav.H02 e H06, 2565-641, Torres Vedras, Portugal. fescaso@ccia.uned.es
Elisabete Malafaia. Instituto Dom Luiz, Universidade de Lisboa. Edifício C6, Campo Grande, 1749-016 Lisboa, Portugal; Museu Nacional de História Natural e da Ciência, Rua da Escola Politécnica 56/58, 1250-102 Lisboa, Portugal; Laboratório de Paleontologia e Paleoecologia, Sociedade de História Natural, Polígono Industrial do Alto do Ameal, Pav.H02 e H06, 2565-641, Torres Vedras, Portugal. emalafaia@gmail.com
Carlos de Miguel Chaves. Grupo de Biología Evolutiva, Facultad de Ciencias, UNED, c/ Senda del Rey, 9, 28040 Madrid, Spain. carlos.miguelchaves@gmail.com
Iván Narváez. Grupo de Biología Evolutiva, Facultad de Ciencias, UNED, c/ Senda del Rey, 9, 28040 Madrid, Spain. i.narvaez.padilla@gmail.com
Adán Pérez-García. Grupo de Biología Evolutiva, Facultad de Ciencias, UNED, c/ Senda del Rey, 9, 28040 Madrid, Spain; Laboratório de Paleontologia e Paleoecologia, Sociedade de História Natural, Polígono Industrial do Alto do Ameal, Pav.H02 e H06, 2565-641, Torres Vedras, Portugal. paleontologo@gmail.com
Nuno Pimentel. Instituto Dom Luiz, Universidade de Lisboa, edifício C6, Campo Grande, 1749-016 Lisboa, Portugal. pimentel@fc.ul.pt
Bruno C. Silva. Laboratório de Paleontologia e Paleoecologia, Sociedade de História Natural, Polígono Industrial do Alto do Ameal, Pav.H02 e H06, 2565-641, Torres Vedras, Portugal. bcsilva.paleo@gmail.com
Francisco Ortega. Grupo de Biología Evolutiva, Facultad de Ciencias, UNED, c/ Senda del Rey, 9, 28040 Madrid, Spain; Laboratório de Paleontologia e Paleoecologia, Sociedade de História Natural, Polígono Industrial do Alto do Ameal, Pav.H02 e H06, 2565-641, Torres Vedras, Portugal. fortega@ccia.uned.es
Keywords: Sauropoda; Upper Jurassic; Lusitanian Basin; Bombarral Sub-basin; Arruda Sub-basin; Turcifal Sub-basin
Mocho, Pedro, Royo-Torres, Rafael, Escaso, Fernando, Malafaia, Elisabete, de Miguel Chaves, Carlos, Narváez, Iván, Pérez-García, Adán, Pimentel, Nuno, Silva, Bruno C., and Ortega, Francisco. 2017. Upper Jurassic sauropod record in the Lusitanian Basin (Portugal): Geographical and lithostratigraphical distribution. Palaeontologia Electronica 20.2.27A: 1-50. https://doi.org/10.26879/662
palaeo-electronica.org/content/2017/1856-portuguese-sauropods
Copyright: © June 2017 Society of Vertebrate Paleontology. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. creativecommons.org/licenses/by/4.0/
INTRODUCTION
The Upper Jurassic sediments of the Lusitanian Basin (Portugal) are known to have yielded abundant fossil vertebrates, in particular dinosaurs, turtles, and crocodyliforms (e.g., Sauvage, 1897-98; Lapparent and Zbyszewski, 1957; Dantas, 1990; Antunes and Mateus, 2003; Ortega et al., 2009, 2013). The sauropod fossil record is particularly rich in this basin, and several historical references deal with this clade (e.g., Sauvage, 1897-98; Lapparent and Zbyszewski, 1957).
A systematic revision of the Portuguese taxa of the Late Jurassic was recently performed (for Dinheirosaurus see Mannion et al., 2012; Tschopp et al., 2015; Lusotitan see Mannion et al., 2013; Mocho et al., 2016c; and for Lourinhasaurus, see Mocho et al., 2014b). Several new occurrences have also been reported (Antunes and Mateus, 2003; Mateus, 2005; Royo-Torres et al., 2006; Yagüe et al., 2006; Mateus, 2009; Royo-Torres et al., 2009; Ortega et al., 2010; Mannion et al., 2012; Mocho et al., 2012, 2013a, 2013b; Mateus et al., 2014; Mocho et al., 2014a, 2016b, 2017). This new information suggests a more diverse scenario for sauropod fauna during the Late Jurassic of the Lusitanian Basin than previously considered, with the identification of a clade previously unidentified in this basin, the non-neosauropod eusauropod group Turiasauria. Royo-Torres et al. (2006) suggested the presence of Turiasauria in the Portuguese Upper Jurassic record, and this hypothesis was subsequently corroborated by the presence of new teeth and postcranial material (Mateus, 2009; Royo-Torres et al., 2009; Ortega et al., 2010; Mocho et al., 2012; Royo-Torres et al., 2014a; Mocho et al., 2016b). Four taxa, so far exclusively, were described in the Upper Jurassic of the Lusitanian Basin: the camarasaurid Lourinhasaurus alenquerensis (Lapparent and Zbyszewski, 1957; Dantas et al., 1998; Mocho et al., 2014b); the diplodocid Dinheirosaurus lourinhanensis (Bonaparte and Mateus, 1999; Mannion et al., 2012; recently referred as Supersaurus lourinhanensis by Tschopp et al., 2015), the basal macronarian and putative brachiosaurid Lusotitan atalaiensis (Lapparent and Zbyszewski, 1957; Antunes and Mateus, 2003; Mannion et al., 2013), and the turiasaur Zby atlanticus (Mateus et al., 2014).
The relationships between the Portuguese Upper Jurassic record and North American Morrison Formation dinosaur fauna have been widely discussed. The relatively abundant Portuguese vertebrate fossil record is important in understanding the relationships between North American and European fauna in this period. A combination of shared and exclusive taxa has been used as an argument to justify processes of dispersion and vicariance (Galton, 1980; Pérez-Moreno et al., 1999; Antunes and Mateus, 2003; Escaso et al., 2007a; Ortega et al., 2013). The relationship of the Portuguese Late Jurassic sauropods to taxa from the North American Upper Jurassic Morrison Fm. (e.g., Lapparent and Zbyszewski, 1957) is currently believed to be less close than to other dinosaur groups (Galton, 1980; Pérez-Moreno et al., 1999; Mateus and Antunes, 2003; Escaso et al., 2007a; Malafaia et al., 2007, 2010a, 2010b; Hendrickx and Mateus, 2014; Malafaia et al., 2015) with no species or even genera reported on either side of the Atlantic. Recently, Tschopp et al. (2015) have suggesed the presence of the genus Supersaurus in the Upper Jurassic of the Lusitanian Basin.
The present study provides a stratigraphic context for the Portuguese Late Jurassic sauropods from the Lusitanian Basin, considering several geological areas, such as the Bombarral (Bombarral-Alcobaça and Consolação), Turcifal, and Arruda Sub-basins. Many specimens are reported and briefly analysed here for the first time, including several specimens found in Torres Vedras, Lourinhã, Peniche, Caldas da Rainha, and Pombal. Several specimens are under preparation for study, but a preliminary systematic evaluation is presented here. This study aims to provide information about the composition of the sauropod fauna in the Upper Jurassic sequence of the Lusitanian Basin.
Geological Context
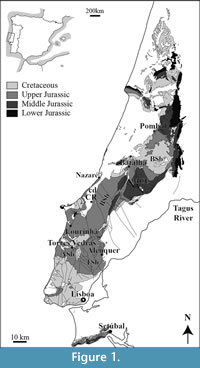 The Lusitanian Basin is located in the west region of the Iberian Peninsula, and it is related to the opening of the North Atlantic. This N-S elongated basin has a maximum extension of 225 km x 70 km (Kullberg, 2000). It is part of a set of marginal and peri-North Atlantic basins, which began to be differentiated in the Triassic due to the fragmentation of Pangaea, and evolved during the Jurassic and Cretaceous, when this shore of the Atlantic became a passive margin (Ribeiro et al., 1979; Boillot et al., 1978; Kullberg et al., 2006; Pena dos Reis et al., 2011; Kullberg et al., 2013). The Lusitanian Basin sedimentary sequence was deposited from the Middle Triassic (?Ladinian - Carnian) (Rocha et al., 1996) to the Late Cretaceous (Turonian) (Rey, 1999) (Figure 1). The evolution of the basin took place mainly in an extensional tectonic context (Kullberg et al., 2006) and in some regions, this sedimentary sequence reaches a thickness of 5000 m (Ribeiro et al., 1979). From the Late Cretaceous onwards, alpine compressive episodes led to the cessation of sedimentation and ultimately the inversion of the basin, with predominant up-lift and the exposure of most of its Mesozoic sequence.
The Lusitanian Basin is located in the west region of the Iberian Peninsula, and it is related to the opening of the North Atlantic. This N-S elongated basin has a maximum extension of 225 km x 70 km (Kullberg, 2000). It is part of a set of marginal and peri-North Atlantic basins, which began to be differentiated in the Triassic due to the fragmentation of Pangaea, and evolved during the Jurassic and Cretaceous, when this shore of the Atlantic became a passive margin (Ribeiro et al., 1979; Boillot et al., 1978; Kullberg et al., 2006; Pena dos Reis et al., 2011; Kullberg et al., 2013). The Lusitanian Basin sedimentary sequence was deposited from the Middle Triassic (?Ladinian - Carnian) (Rocha et al., 1996) to the Late Cretaceous (Turonian) (Rey, 1999) (Figure 1). The evolution of the basin took place mainly in an extensional tectonic context (Kullberg et al., 2006) and in some regions, this sedimentary sequence reaches a thickness of 5000 m (Ribeiro et al., 1979). From the Late Cretaceous onwards, alpine compressive episodes led to the cessation of sedimentation and ultimately the inversion of the basin, with predominant up-lift and the exposure of most of its Mesozoic sequence.
The Upper Jurassic sequence is the focus of this study, and represents the main subsidence phase of the basin, with an accumulation of over 3 km in sedimentary thickness, in a short period, at its main depocenters (Pena dos Reis et al., 1996, 2000). It ranges from the middle Oxfordian to the boundary with the Lower Cretaceous (Schneider et al., 2009). The Upper Jurassic sedimentary sequence represents a third rifting episode (Rasmussen et al., 1998; Kullberg et al., 2006), marked by intense subsidence and an internal differentiation in three main sectors (Rocha and Soares, 1984). The northern sector extends from Coimbra to the Nazaré fault and is characterized by lower subsidence; the central sector extends from this main tectonic line to the Lisboa area and presents the maximum subsidence; and the southern sector extends from Lisboa to the alpine Arrabida chain, with the lowest subsidence. This episode of rifting was marked by an important sedimentary input into the whole basin, which progressively filled the basin (Guéry, 1984; Hill, 1988; Wilson, 1988; Pena dos Reis et al., 2000; Kullberg et al., 2006, 2013). From the lower Kimmeridgian up to the top of the Upper Jurassic, the sedimentary sequence was dominated by abundant siliciclastic inputs, with a mainly continental and transitional signature (e.g., Hill, 1988; Manuppella et al., 1999; Kullberg et al., 2006). As accommodation space decreased, coastal environments became shallower and siliciclastics prograded into the basin, with the gradual development of fluvial environments, giving origin to a predominantly braided fluvial sedimentation in Berriasian times (Torres Vedras Group). Late Jurassic paleogeographic reconstructions point to a narrow trough developing from NE proximal areas to SW distal areas, with both longitudinal axial drainage and lateral inputs from the eastern and western basement borders (Pena dos Reis et al., 2011). Palaeoenvironments grade from proximal braided fluvial deposits to distal fine-grained meandering, and even deltaic to coastal deposits. Siliciclastics are clearly predominant, but some minor carbonate and shell-rich intercalations may be found, particularly in distal areas (Pena dos Reis et al., 2011).
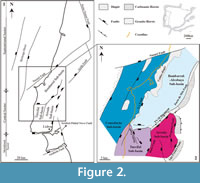 The central sector corresponds mainly to distal fluvio-deltaic and coastal environments, which produced the richest fossil record of terrestrial vertebrates in the Lusitanian Basin (e.g., Lapparent and Zbyszewski, 1957; Dantas, 1990; Antunes and Mateus, 2003; Ortega et al., 2009). No sauropod osteological remains have yet been identified in the Upper Jurassic of the southern or northern sectors of the Lusitanian Basin. The Central Sector of the Lusitanian Basin has been divided into three Upper Jurassic sub-basins, the Turcifal, Arruda, and Bombarral (Figure 2.1), based on isopachs and facies distribution (Pena dos Reis et al., 1996, 2000). The stratigraphy of the Upper Jurassic sequence of the Lusitanian Basin is complex, due the profuse lateral heterogeneity and the proposal of several and different stratigraphic approaches (e.g., Hill, 1988; Leinfelder, 1993; Manuppella et al., 1999; Kullberg et al., 2006; Schneider et al., 2009; Martinius and Gowland, 2011; Taylor et al., 2014). Figure 3 presents the stratigraphic correlations between the nomenclature proposed for the areas considered in this manuscript: i) Turcifal Sub-basin (stratigraphy based on Pereda-Suberbiola et al., 2005; Kullberg et al., 2006; Schneider et al., 2009); ii) Arruda Sub-basin (stratigraphy based on Kullberg et al., 2006); iii) Consolação Sub-basin areas: Torres Vedras-Lourinhã-Peniche (stratigraphy based on Manuppella et al., 1999), Foz do Arelho-Nazaré coastal sector (stratigraphy based on Kullberg et al., 2006; Azerêdo et al., 2010); iv) Bombarral-Alcobaça Sub-basin (stratigraphy based on Kullberg et al., 2006; Azerêdo et al., 2010); v) Batalha-Leiria region (stratigraphy based on Manuppella et al., 2000; Kullberg et al. 2006; Escaso et al., 2007a), and vi)
The central sector corresponds mainly to distal fluvio-deltaic and coastal environments, which produced the richest fossil record of terrestrial vertebrates in the Lusitanian Basin (e.g., Lapparent and Zbyszewski, 1957; Dantas, 1990; Antunes and Mateus, 2003; Ortega et al., 2009). No sauropod osteological remains have yet been identified in the Upper Jurassic of the southern or northern sectors of the Lusitanian Basin. The Central Sector of the Lusitanian Basin has been divided into three Upper Jurassic sub-basins, the Turcifal, Arruda, and Bombarral (Figure 2.1), based on isopachs and facies distribution (Pena dos Reis et al., 1996, 2000). The stratigraphy of the Upper Jurassic sequence of the Lusitanian Basin is complex, due the profuse lateral heterogeneity and the proposal of several and different stratigraphic approaches (e.g., Hill, 1988; Leinfelder, 1993; Manuppella et al., 1999; Kullberg et al., 2006; Schneider et al., 2009; Martinius and Gowland, 2011; Taylor et al., 2014). Figure 3 presents the stratigraphic correlations between the nomenclature proposed for the areas considered in this manuscript: i) Turcifal Sub-basin (stratigraphy based on Pereda-Suberbiola et al., 2005; Kullberg et al., 2006; Schneider et al., 2009); ii) Arruda Sub-basin (stratigraphy based on Kullberg et al., 2006); iii) Consolação Sub-basin areas: Torres Vedras-Lourinhã-Peniche (stratigraphy based on Manuppella et al., 1999), Foz do Arelho-Nazaré coastal sector (stratigraphy based on Kullberg et al., 2006; Azerêdo et al., 2010); iv) Bombarral-Alcobaça Sub-basin (stratigraphy based on Kullberg et al., 2006; Azerêdo et al., 2010); v) Batalha-Leiria region (stratigraphy based on Manuppella et al., 2000; Kullberg et al. 2006; Escaso et al., 2007a), and vi) Pombal region (stratigraphy based on Kullberg et al., 2006; Malafaia et al., 2010a). This nomenclature will be used in the present study. The term Lourinhã Formation was proposed by Hill (1988) and has been widely used in vertebrate paleontological literature. This formation was defined on the coastal sections south of São Bernardino (Peniche) to Porto da Calada (Mafra). This formation seems to be coincident with the upper part of the Alcobaça, Praia da Amoreira-Porto Novo, Sobral, Freixial, and Bombarral Formations referenced by other authors, including the Portuguese Geological Surveys (e.g., Manuppella et al., 1999). The use of this unit outside the area corresponding to the Consolação Sub-basin of Taylor et al. (2014) is not completely established in literature (e.g., Hill, 1988; Kullberg et al., 2006; Schneider et al., 2009; Ribeiro and Mateus, 2012; Kullberg et al., 2013; Mateus et al., 2013). Yagüe et al. (2006) therefore incorporate the entire Alcobaça Formation and the previously defined Lourinhã Formation into a new lithostratigraphic unit, the Lourinhã Group. The stratigraphic proposals of Hill (1988) and Yaguë et al. (2006) are also presented in the Figure 3.
Pombal region (stratigraphy based on Kullberg et al., 2006; Malafaia et al., 2010a). This nomenclature will be used in the present study. The term Lourinhã Formation was proposed by Hill (1988) and has been widely used in vertebrate paleontological literature. This formation was defined on the coastal sections south of São Bernardino (Peniche) to Porto da Calada (Mafra). This formation seems to be coincident with the upper part of the Alcobaça, Praia da Amoreira-Porto Novo, Sobral, Freixial, and Bombarral Formations referenced by other authors, including the Portuguese Geological Surveys (e.g., Manuppella et al., 1999). The use of this unit outside the area corresponding to the Consolação Sub-basin of Taylor et al. (2014) is not completely established in literature (e.g., Hill, 1988; Kullberg et al., 2006; Schneider et al., 2009; Ribeiro and Mateus, 2012; Kullberg et al., 2013; Mateus et al., 2013). Yagüe et al. (2006) therefore incorporate the entire Alcobaça Formation and the previously defined Lourinhã Formation into a new lithostratigraphic unit, the Lourinhã Group. The stratigraphic proposals of Hill (1988) and Yaguë et al. (2006) are also presented in the Figure 3.
MATERIAL AND METHODS
The studied material is mainly deposited in Portuguese institutions, excluding one specimen found in the Muséum National d’Histoire Naturelle (Paris, France) (see list in Appendix 1). All the specimens are described and discussed based on direct observations. We use “Romerian” terms for anatomical structures for the structures (e.g., “centrum”) and their orientation (e.g., “anterior”) (Wilson, 2006). This study applies the landmark-based terminology of Wilson (1999, 2012) and Wilson et al. (2011) for vertebral laminae and fossae.
Anatomical Abbreviations
aacet, articulation for the acetabulum; acc, acromial crest; acet, acetabulum; ant. spdl, anterior spinodiapophyseal lamina; asp, ascending process; aspa, articular surface for the ascending process; awf, apical wear facet; br, bridge; cc, cnemial crest; cml, camellae; cpol, centropostzygapophyseal lamina; cr, caudal rib; cwf, carina wear facet; di, diapophyses; dpc, deltopectoral crest; ec, epicondyle; f, fossa; fic, fibular condyle; ft, fourth trochanter; gl, glenoid; gr, groove; ibi, incipient bifurcation; ilped, iliac peduncle; isped, ischiatic peduncle; lag, labial groove; lb, lateral bulge; lf, lingual facets; lic, lingual crest; lt, lateral trochanter; hy, hyposphene; lat.spol, lateral spinopostzygapophyseal lamina; of, obturator foramen; pafc, posterior astragalar fossa crest; paf, posterior astragalar fossa; pcdl, posterior centrodiapophyseal lamina; pcpl, posterior centroparapophyseal lamina; pfr, pneumatic foramen; podl, postzygodiapophyseal lamina; posl, postspinal lamina; post. spdl, posterior spinodiapophyseal lamina; poz, postzygapophyses; prsl, prespinal lamina; prdl, prezygodiapophyseal lamina; prz, prezygapophyses; spof, spinopostzygapophyseal fossa; spol, spinopostzygapophyseal lamina; sprf, spinoprezygapophyseal fossa; sprl, spinoprezygapophyseal lamina; tap, triangular aliform process; tb, tuberosity; tia, tibial articular surface; tic, tibial condyle; vh, ventral hollow; vlc, ventrolateral crest; vpr, ventral process.
Institutional Abbreviations
MG, Museu Geológico (Lisboa, Portugal); ML, Museu da Lourinhã (Lourinhã, Portugal); MMPM, Museu Municipal de Porto de Mós (Porto de Mós, Portugal); MMB, Museu Municipal do Bombarral (Bombarral, Portugal); MMLT, Museu Municipal Leonel Trindade (Torres Vedras, Portugal); MNHN, Muséum National d’Histoire Naturelle (Paris, France); MNHNUL, Museu Nacional de História Natural e da Ciência (Lisboa, Portugal); SHN, Sociedade de História Natural, plus JJS for the José Joaquim dos Santos collection (Torres Vedras, Portugal).
UPPER JURASSIC SAUROPOD RECORD OF THE LUSITANIAN BASIN
The sauropod record will be presented as following a palaeogeographic distribution, corresponding mainly to the Upper Jurassic sub-basins of the central sector (Figure 2), however, it must be noted that these sub-basins have been defined based on, essentially, geodynamic features, namely the existence of three depocentres separated by main tectonic fault lines and by less subsident areas. These areas could eventually represent paleotopographic highs, which may account for some palaeogeographic differences between sub-basins. Sub-basins could therefore represent low-lands where water, vegetation, and vertebrates would have had better conditions in which to develop and accumulate. If so, these areas might represent different contemporaneous communities, occupying slightly different habitats or palaeoenvironments, more or less open to coastal influences or connections.
Bombarral Sub-basin
The Bombarral Sub-basin is the widest Upper Jurassic sub-basin of the central sector of the Lusitanian Basin. It is also the richest area with respect to fossil sites with vertebrates (e.g., Lapparent and Zbyszewski, 1957; Ortega et al., 2009). This basin is delimited in the north by the Nazaré Fault and in the east and south by the Porto-Tomar, Arrife, and Torres-Vedras-Montejunto Faults (Kullberg, 2000; Kullberg et al., 2006, 2013; Figure 2.1). The Torres Vedras-Montejunto Fault separates the Bombarral Sub-basin from the half-graben Turcifal and Arruda Sub-basins (Kullberg et al., 2006, 2013). Taylor et al. (2014) established two new sub-basins replacing the Bombarral Sub-Basin, called the Consolação and Bombarral-Alcobaça Sub-basins (Figure 2.2).
The Consolação Sub-basin is located on the west of the Lourinhã Fault and the Caldas Diapir, and is bounded in the north by the Nazaré Fault (Figure 2.2). This sub-basin includes the Upper Jurassic coastal sector from Praia da Consolação (Peniche) to Santa Cruz (Torres Vedras) and some Upper Jurassic cliffs north of Peniche, and from Nazaré to Foz do Arelho. The remaining area of the previously defined Bombarral Sub-basin, and that on the east of the Lourinhã Fault and the Caldas Diapir, is now included in the Bombarral-Alcobaça Sub-basin (Figure 2.2; Taylor et al., 2014). We follow the sub-division of the Lusitanian Basin Central Sector proposed by Kullberg et al. (2006, 2013) always referring to the correspondent nomenclature proposed by Taylor et al. (2014).
The presence of several dinosaur remains in the Bombarral Sub-basin has been reported since the end of the nineteenth century (e.g., Sauvage, 1897-98; Lapparent and Zbyszewski, 1957; Dantas, 1990; Antunes and Mateus, 2003), most notably those from Upper Jurassic sediments outcropping in the Lourinhã, Peniche, and Pombal municipalities (e.g., Lapparent and Zbyszewski, 1957; Pérez-Moreno et al., 1999; Antunes and Mateus, 2003). Several occurrences of non-sauropod dinosaurs (theropods, ornithopods, and thyreophorans) were also reported. Theropod fauna are comprised of basal forms representing Ceratosauria (Ceratosaurus sp.), Megalosauroidea (Torvosaurus gurneyi), and Allosauroidea, such as Allosaurus and Lourinhanosaurus antunesi (e.g., Dantas, 1987; Mateus et al., 1997; Mateus, 1998; Dantas et al., 1999; Pérez-Moreno et al., 1999; Mateus and Antunes, 2000a, 2000b; Rauhut, 2000; Mateus, 2005, 2006; Mateus et al., 2006; Malafaia et al., 2007, 2008, 2010a, 2010b; Hendrickx and Mateus, 2012, 2014; Malafaia et al., 2015, 2016, 2017). Derived theropods are also represented by mostly isolated specimens related to the clade Coelurosauria, including the basal tyrannosauroid Aviatyrannis, indeterminate dromeosaurids, and several teeth attributed to cf. Archaeopteryx (Weigert, 1995; Zinke, 1998; Zinke and Rauhut, 1994; Antunes and Mateus, 2003; Mateus, 2005; Malafaia et al., 2007, 2010a, 2014). The most important Bombarral Sub-basin theropod fossil sites are reported from Andrés in Pombal (Dantas et al., 1999; Pérez-Moreno et al., 1999; Malafaia et al., 2007, 2010a), Paimogo in Lourinhã (Dantas, 1987; Mateus et al., 1997), Vale de Pombas-Praia Vermelha coastal cliffs in Lourinhã-Peniche (Mateus, 2005; Hendrickx and Mateus, 2014), and the Guimarota mine in Leiria (Zinke, 1998; Rauhut, 2000, 2003; Rauhut and Fechner, 2005).
The Bombarral Sub-basin is also rich in ornithischian remains, with the presence of at least four ornithopod and three thyreophoran forms. Several historical specimens were referred to the genus Omosaurus (Zbyszewski, 1946; Lapparent and Zbyszewski, 1957), but some part of this material, as well as new specimens, were later assigned to the genus Dacentrurus (e.g., Galton, 1991; Maidment et al., 2008; Ortega et al., 2009). This genus is also present in the Spanish Villar del Arzobisbo Formation (Kimmeridgian-Berriasian in age), as the only known shared dinosaur genus between these territories for the Late Jurassic (Cobos et al., 2010; Cobos and Gascó, 2013). More recently, two other stegosaurian forms were identified in the Upper Jurassic levels of the Bombarral Sub-basin. Escaso et al. (2007a) referred a partial individual found in Casal Novo (Alcobaça Fm., Leiria) to the North American Morrison Fm. genus Stegosaurus, proposing a Late Jurassic contact for the Iberian and North American fauna. Mateus et al. (2009) established a new stegosaurian taxon, Miragaia longicollum, a form closely related to Dacentrurus. Cobos et al. (2010) proposed Miragaia as synonymous with of Dacentrurus, a hypothesis followed by Escaso (2014) and Maidment et al. (2015).
The Upper Jurassic ornithopod record of the Bombarral Sub-basin is represented by the dryosaurid Eousdryosaurus (Escaso et al., 2014) and two styracosternans: Camptosaurus (=Uteodon) aphanoecetes, previously identified in the Morrison Formation (Escaso et al., 2010a, 2010b) and the exclusive Portuguese form Draconyx loureiroi (Mateus and Antunes, 2001). Another ornithopod, based on teeth from the Guimarota mine, was described as Phyllodon henkeli (Thulborn, 1973). Rauhut (2001) referred some additional teeth as well as three partial dentaries to this taxon.
Other vertebrate groups have been reported, such as fishes (Sauvage, 1897-98; Kriwet, 2000; Balbino, 2003), amphibians (Wiechmann, 2000), turtles (Sauvage, 1897-98; Bräm, 1973; Antunes et al., 1988; Gassner, 2000; Pérez-García and Ortega, 2011; Pérez-García, 2015); sphenodonts (Ortega et al., 2006; Malafaia et al., 2010a), squamates (Seiffert, 1973; Broschinski, 2000; Caldwell et al., 2015), pterosaurs (Dantas, 1987; Wiechmann and Gloy, 2000), crocodyliforms (Sauvage, 1897-98; Krebs and Schwarz, 2000; Schwarz and Fechner, 2004; Schwarz and Salisbury, 2005; Tennant and Mannion, 2014; Russo et al., 2017), mammals (e.g., Hahn, 1971; Krusat, 1980; Hahn and Hahn, 2000; Krebs, 2000; Martin, 2000; Martin and Nowotny, 2000; Martin, 2005), and possible plesiosaurs (Castanhinha and Mateus, 2007).
The sauropods are well represented in this sub-basin, being recognised in many fossil sites, with numerous occurrences (e.g., Lapparent and Zbyszewski, 1957; Dantas, 1990; Antunes and Mateus, 2003; Mateus, 2005; Mocho et al., 2013a, 2013b, 2014a, 2016a, 2017). We will report the sauropod record in the following different areas of the Bombarral Sub-basin: i) northern region of Maciço Calcário Estremenho; ii) coastal sector from Foz do Arelho to Nazaré; iii) Alcobaça, Bombarral and A-dos-Cunhados; iv) North Peniche; and v) Praia da Consolação-Lourinhã-Torres Vedras coastal sector. This subdivision seems useful considering the profuse lateral lithological heterogeneity and the proposal of several stratigraphic approaches for each area within this sub-basin, and also to provide a more accurate overview of the abundant sauropod fossil record in the Bombarral Sub-basin.
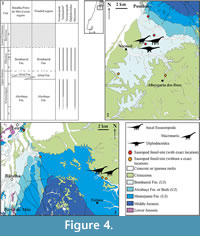 Northern region of Maciço Calcário Estremenho (Figure 4, Figure 5). The region north of the Maciço Calcário Estremenho (MCE) is rich in transitional to continental Upper Jurassic deposits (Camarate França and Zbyszewski, 1963; Teixeira et al., 1966; Manuppella et al., 2000), including several fossil sites in the Pombal, Leiria, Batalha, and Porto-de-Mós municipalities. Several important vertebrate sites are identified in this area, notably the Guimarota mine (Leiria), and Andrés (Pombal) and Casal Novo (Batalha) quarries (e.g., Peréz-Moreno et al., 1999; Rauhut, 2000; Escaso et al., 2007a; Malafaia et al., 2007, 2010a). The sauropod record is relatively poor in this sector of the Bombarral Sub-basin and is mainly based on incomplete specimens (Figure 5). Samples come from sediments in the Alcobaça and Bombarral Formations (Figure 4.2-3, Figure 5), and there is published and unpublished material (e.g., Malafaia et al., 2010a; Mocho et al., 2016a, 2017). The Alcobaça Fm. is interpreted as having been deposited in marine environments, transiting to fluvio-lacustrine deposits at the top of the formation. In this area, the Alcobaça Fm. is lower Kimmeridgian-lower Tithonian in age, being considered as laterally correlated with the marine Abadia Fm., present in the south of the Lusitanian Basin Central Sector (Manuppella et al., 2000). Some important fossil sites found in the Alcobaça Fm. correspond to fluvial (Escaso et al., 2007a) and lagoonal deposits (Schudack, 2000). The Bombarral Fm. is composed of fluvio-lacustrine deposits, dated to the late Kimmeridgian-Tithonian (Manuppella et al., 2000).
Northern region of Maciço Calcário Estremenho (Figure 4, Figure 5). The region north of the Maciço Calcário Estremenho (MCE) is rich in transitional to continental Upper Jurassic deposits (Camarate França and Zbyszewski, 1963; Teixeira et al., 1966; Manuppella et al., 2000), including several fossil sites in the Pombal, Leiria, Batalha, and Porto-de-Mós municipalities. Several important vertebrate sites are identified in this area, notably the Guimarota mine (Leiria), and Andrés (Pombal) and Casal Novo (Batalha) quarries (e.g., Peréz-Moreno et al., 1999; Rauhut, 2000; Escaso et al., 2007a; Malafaia et al., 2007, 2010a). The sauropod record is relatively poor in this sector of the Bombarral Sub-basin and is mainly based on incomplete specimens (Figure 5). Samples come from sediments in the Alcobaça and Bombarral Formations (Figure 4.2-3, Figure 5), and there is published and unpublished material (e.g., Malafaia et al., 2010a; Mocho et al., 2016a, 2017). The Alcobaça Fm. is interpreted as having been deposited in marine environments, transiting to fluvio-lacustrine deposits at the top of the formation. In this area, the Alcobaça Fm. is lower Kimmeridgian-lower Tithonian in age, being considered as laterally correlated with the marine Abadia Fm., present in the south of the Lusitanian Basin Central Sector (Manuppella et al., 2000). Some important fossil sites found in the Alcobaça Fm. correspond to fluvial (Escaso et al., 2007a) and lagoonal deposits (Schudack, 2000). The Bombarral Fm. is composed of fluvio-lacustrine deposits, dated to the late Kimmeridgian-Tithonian (Manuppella et al., 2000).
Near Pombal, sauropod fossils were only identified in the sediments of the Bombarral Formation. A great accumulation of fossils representing a relatively diverse vertebrate fauna was identified at the Andrés locality in the Tithonian levels of the Bombarral Fm. (Dantas et al., 1999; Pérez-Moreno et al., 1999; Malafaia et al., 2007, 2009a, 2010a). Several teeth and postcranial material assigned to Sauropoda were collected at this fossil site (Figure 5.5). A preliminary analysis of the teeth allowed the recognition of four different morphotypes: spatulate, heart-, compressed cone-chisel-, and peg-shaped, which led Malafaia et al. (2010a) to suggest the presence of forms related to Diplodocoidea, Turiasauria, and Titanosauriformes. However, the presence of compressed cone-chisel-shaped teeth in Europasaurus (Sander et al., 2006), which was recently considered as a non-titanosauriform macronarian (Carballido and Sander, 2014), might indicate that this tooth morphology was already present in non-titanosauriform macronarians. More teeth and postcranial elements from the Andrés site are being prepared (EM, personal commun., 2015).
 Also in the Pombal area, a middle/posterior caudal vertebra (Figure 5.4, MG 4811) was found in Albergaria dos Doze (Pombal), probably from the Bombarral Formation (Figure 4.2). Lapparent and Zbyszewski (1957) attributed this element to the theropod Megalosaurus pombali. Subsequently, Mateus (2005) considered this vertebra to belong to an indeterminate theropod. This vertebra probably corresponds to an indeterminate sauropod based on its general morphology, transversely convex ventral face and an anteroposteriorly short neural arch. A middle or posterior dorsal neural spine was also found in this area, near Vermoil (Bombarral Fm., Figure 5.8-9, MNHN.unnumbered). This neural spine probably represents a eusauropod due the presence of a transversely expanded neural spine and a well-defined prespinal lamina (Mocho et al., 2017). More recently, a new fossil site containing a partial sauropod skeleton (dorsal vertebrae and ribs) was found in Pombal (Figure 5.14) during fieldwork carried out by the Museu Nacional de História Natural e da Ciência (Lisboa, Portugal).
Also in the Pombal area, a middle/posterior caudal vertebra (Figure 5.4, MG 4811) was found in Albergaria dos Doze (Pombal), probably from the Bombarral Formation (Figure 4.2). Lapparent and Zbyszewski (1957) attributed this element to the theropod Megalosaurus pombali. Subsequently, Mateus (2005) considered this vertebra to belong to an indeterminate theropod. This vertebra probably corresponds to an indeterminate sauropod based on its general morphology, transversely convex ventral face and an anteroposteriorly short neural arch. A middle or posterior dorsal neural spine was also found in this area, near Vermoil (Bombarral Fm., Figure 5.8-9, MNHN.unnumbered). This neural spine probably represents a eusauropod due the presence of a transversely expanded neural spine and a well-defined prespinal lamina (Mocho et al., 2017). More recently, a new fossil site containing a partial sauropod skeleton (dorsal vertebrae and ribs) was found in Pombal (Figure 5.14) during fieldwork carried out by the Museu Nacional de História Natural e da Ciência (Lisboa, Portugal).
A large area with Upper Jurassic continental sediments from the Alcobaça and Bombarral Formations is located across the localities of Batalha, Vila Nova de Ourém, Leiria, and Porto-de-Mós. In the Alcobaça Fm., one of the most important accumulations was found in the Guimarota mine, in Leiria (e.g., Rauhut, 2000). The Guimarota fossil record is relatively poor in sauropod remains, but some small teeth referred to Brachiosauridae (Rauhut, 2000), or to cf. Lusotitan atalaiensis (Mateus, 2005) were found (Figure 5.2-3). Sauvage (1897-98) and Lapparent and Zbyszewski (1957) also reported some localities with dinosaur remains near Vila Nova de Ourém and Porto de Mós (probably from the Alcobaça Fm.). Two teeth, one had a heart-shaped morphology probably referrable to Turiasauria (Figure 5.6-7, MG 16; Mocho et al., 2012, 2016b) and other had a compressed cone-chisel-shaped morphology (Figure 5.1, MG 8779) common in Titanosauriformes, but also found in the basal macronarian Europasaurus (Sander et al., 2006; Carballido and Sander, 2014). A posterior caudal vertebra (MMPM.P/554) from an indeterminate sauropod was also found in Fonte do Oleiro (Alcobaça Fm.), south of the town of Porto de Mós (Mocho et al., 2017).
Mocho et al. (2017) described some sauropod specimens found close to the town of Batalha, and they are deposited in the collections of the Museu Geológico and Museu Municipal de Porto-de-Mós, including: i) a partial dorsal centrum and a partial caudal series collected in Abadia (Figure 5.10-13, MG 4974), and ii) a set of sauropod bones including dorsal and caudal vertebrae and a fragmentary ischium from Batalha, which might represent a member of Diplodocinae (Figure 5.15-18, MG 30389). The stratigraphic and geographic context of these specimens is not clear, and they might pertain to the Alcobaça Fm. or to the Bombarral Fm.
The presence of non-neosauropod eusauropods, probably turiasaurs, indeterminate diplodocoids (including members of Diplodocinae), and macronarians, was identified in the area located in the north of the Maciço Calcário Estremenho (Figure 4.1).
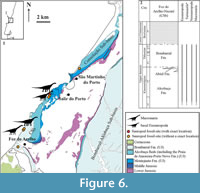 Coastal sector from Foz do Arelho to Nazaré (Figure 6, Figure 7). An extensive sector with Upper Jurassic sediments in the Bombarral (following Kullberg et al., 2006, 2013; Azerêdo et al., 2010), or Consolação Sub-basin (following Taylor et al., 2014), crops westwards out of the Caldas Diapir (Figure 6.1). This sequence extends from the coastal cliffs of Foz do Arelho to Nazaré (e.g., Hill, 1988; Kullberg et al., 2006), and includes sediments of the Alcobaça (Kimmeridgian to basal Tithonian) and Bombarral (Tithonian) Formations (Figure 6.2) (Kullberg et al., 2006, 2013; Schneider et al., 2009; Azerêdo et al., 2010). Both formations are particularly rich in fossil vertebrate finds. One of the most important fossil sites is the historical site in Murteiras (Caldas da Rainha) (Lapparent and Zbyszewski, 1957), due the presence of several specimens referred to Dacentrurus (Galton, 1991; Antunes and Mateus, 2003; Maidment et al., 2008; Escaso, 2014). No sauropod remains were reported there, however (Lapparent and Zbyszewski, 1957).
Coastal sector from Foz do Arelho to Nazaré (Figure 6, Figure 7). An extensive sector with Upper Jurassic sediments in the Bombarral (following Kullberg et al., 2006, 2013; Azerêdo et al., 2010), or Consolação Sub-basin (following Taylor et al., 2014), crops westwards out of the Caldas Diapir (Figure 6.1). This sequence extends from the coastal cliffs of Foz do Arelho to Nazaré (e.g., Hill, 1988; Kullberg et al., 2006), and includes sediments of the Alcobaça (Kimmeridgian to basal Tithonian) and Bombarral (Tithonian) Formations (Figure 6.2) (Kullberg et al., 2006, 2013; Schneider et al., 2009; Azerêdo et al., 2010). Both formations are particularly rich in fossil vertebrate finds. One of the most important fossil sites is the historical site in Murteiras (Caldas da Rainha) (Lapparent and Zbyszewski, 1957), due the presence of several specimens referred to Dacentrurus (Galton, 1991; Antunes and Mateus, 2003; Maidment et al., 2008; Escaso, 2014). No sauropod remains were reported there, however (Lapparent and Zbyszewski, 1957).
Sauropod remains collected in the Alcobaça Formation (following Camarate França and Zbyszewski, 1963; Azerêdo et al., 2010), outcropping in this sector of the Bombarral Sub-basin, (Figure 6.1) include axial elements (Figure 7.1-3, MMPM.P/551) found close to São Martinho do Porto (Mocho et al., 2017), and some unpublished caudal vertebrae and pelvic fragments (Figure 7.18, SHN 537) found in Salir do Porto. An anterior dorsal neural spine (Figure 7.8, MG 4920; Lapparent and Zbyszewski, 1957) of an indeterminate eusauropod from Foz do Arelho (Caldas da Rainha) was collected from sediments of the Bombarral Fm.
 Several teeth were collected in this coastal area, corresponding to different morphotypes: i) heart-shaped teeth (Figure 7.4-7, 7.11-15, 7.16-17), tentatively referred to Turiasauria (see Royo-Torres and Upchurch, 2012; Royo-Torres et al., 2009; Mocho et al., 2012, 2016b); ii) spatulate teeth (Figure 7.9-10; unpublished material housed in MG and SHN) referred to an indeterminate eusauropod (probably a macronarian); and iii) compressed cone-chisel-shaped teeth (unpublished material housed in the SHN), referred to Macronaria. The first of these morphotypes is the most abundant in this coastal sector.
Several teeth were collected in this coastal area, corresponding to different morphotypes: i) heart-shaped teeth (Figure 7.4-7, 7.11-15, 7.16-17), tentatively referred to Turiasauria (see Royo-Torres and Upchurch, 2012; Royo-Torres et al., 2009; Mocho et al., 2012, 2016b); ii) spatulate teeth (Figure 7.9-10; unpublished material housed in MG and SHN) referred to an indeterminate eusauropod (probably a macronarian); and iii) compressed cone-chisel-shaped teeth (unpublished material housed in the SHN), referred to Macronaria. The first of these morphotypes is the most abundant in this coastal sector.
The known and the new occurrences put in evindece a relatively high potential for future discoveries in this area, However, the so far published (Mocho et al., 2012, 2016b, 2017) and unpublished material of sauropods only allows identification of indeterminate sauropods and eusauropods, possibly members of Turiasauria, and indeterminate macronarians. The presence of several compressed cone-chisel teeth in the sedimentary sequence suggests the presence of macronarian forms within the clade formed by Europasaurus + Titanosauriformes (Figure 6.2).
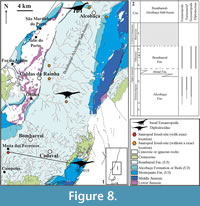 Alcobaça, Bombarral, and A-dos-Cunhados (Figure 8, Figure 9). A large area with Upper Jurassic sediments, included within the Bombarral Sub-basin, is found east of the Lourinhã Fault and the Caldas Diapir, defined as the Bombarral-Alcobaça Sub-basin by Taylor et al. (2014) (Figure 8.1). This area is poorly prospected when compared with other areas, such as the coastal sector from Praia da Consolação to Torres Vedras. In fact, this area is generally covered by soil with natural vegetation or agriculture, resulting in only a few outcrops that could produce vertebrate fossils. The most important localities are Moita dos Ferreiros (Mateus, 2005; Mannion et al., 2012) and Miragaia, where the type material for Miragaia longicollum was found (Mateus et al., 2009). Sauropod remains are scarce in this area (Figure 8.1), although there are also some specimens housed in private collections (e.g., Mateus, 2005; pers. observ., PM), such as a partial tail of an indeterminate sauropod, collected near the town of Bombarral.
Alcobaça, Bombarral, and A-dos-Cunhados (Figure 8, Figure 9). A large area with Upper Jurassic sediments, included within the Bombarral Sub-basin, is found east of the Lourinhã Fault and the Caldas Diapir, defined as the Bombarral-Alcobaça Sub-basin by Taylor et al. (2014) (Figure 8.1). This area is poorly prospected when compared with other areas, such as the coastal sector from Praia da Consolação to Torres Vedras. In fact, this area is generally covered by soil with natural vegetation or agriculture, resulting in only a few outcrops that could produce vertebrate fossils. The most important localities are Moita dos Ferreiros (Mateus, 2005; Mannion et al., 2012) and Miragaia, where the type material for Miragaia longicollum was found (Mateus et al., 2009). Sauropod remains are scarce in this area (Figure 8.1), although there are also some specimens housed in private collections (e.g., Mateus, 2005; pers. observ., PM), such as a partial tail of an indeterminate sauropod, collected near the town of Bombarral.
Some older occurrences from the Alcobaça Formation were found in the Alcobaça, Bombarral, and A-dos-Cunhados regions (Bombarral-Alcobaça Sub-basin) and referred to Sauropoda (Lapparent and Zbyszewski, 1957): i) an heart-shaped tooth (Figure 9.6-7) from Fervença (Sauvage, 1897-98; Lapparent and Zbyszewski, 1957), tentatively referred to an indeterminate eusauropod probably related to Turiasauria (Mocho et al., 2016b); and ii) a posterior caudal vertebra from Chiqueda de Cima (Lapparent and Zbyszewski, 1957), representing an indeterminate sauropod. Some elements were found in the Bombarral Fm.: i) an anterior caudal vertebra (Figure 9.5, MG 4804) of an indeterminate eusauropod from Salir de Matos; ii) a middle caudal vertebrae (Figure 9.3-4, MG 4819, 4821, 4826) of an indeterminate diplodocine from São Gregório da Fanadia; and iii) a large left astragalus (Figure 9.1-2, MMPM.P/75) attributed to an indeterminate eusauropod was found in Imaginário (Caldas da Rainha) (Mocho et al., 2016a, 2017).
 An incomplete skeleton composed of axial elements (cervical and dorsal vertebrae; Figure 9.8, ML 418), considered as aff. Dinheirosaurus (Antunes and Mateus, 2003) and Apatosaurus sp. (Mateus, 2005), was found close to Moita dos Ferreiros (Lourinhã, Bombarral Formation). These elements are poorly preserved. Mannion et al. (2012) distinguished this specimen from Dinheirosaurus lourinhanensis and suggested that it might represent a second diplodocid taxon from the Upper Jurassic of the Lusitanian Basin. Tschopp et al. (2015) suggested that it represents an indeterminate diplodocine different from Dinheirosaurus. The full preparation of the dorsal vertebrae of Dinheirosaurus lourinhanensis will be important to test this hypothesis.
An incomplete skeleton composed of axial elements (cervical and dorsal vertebrae; Figure 9.8, ML 418), considered as aff. Dinheirosaurus (Antunes and Mateus, 2003) and Apatosaurus sp. (Mateus, 2005), was found close to Moita dos Ferreiros (Lourinhã, Bombarral Formation). These elements are poorly preserved. Mannion et al. (2012) distinguished this specimen from Dinheirosaurus lourinhanensis and suggested that it might represent a second diplodocid taxon from the Upper Jurassic of the Lusitanian Basin. Tschopp et al. (2015) suggested that it represents an indeterminate diplodocine different from Dinheirosaurus. The full preparation of the dorsal vertebrae of Dinheirosaurus lourinhanensis will be important to test this hypothesis.
In summary, the dinosaur fauna in the Alcobaça, Bombarral and A-dos-Cunhados area are poorly understood. The recorded sauropod fauna in this area is composed of indeterminate taxa, and indeterminate eusauropods (some tentatively assigned to Turiasauria), and indeterminate diplodocines. One of the diplodocine specimens (ML 418) might represent a diplodocine different from Dinheirosaurus lourinhanensis (Mannion et al., 2012; Tschopp et al., 2015).
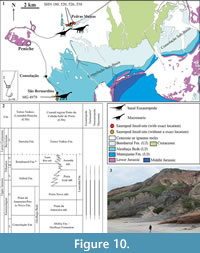 North Peniche (Figure 10, Figure 11). An Upper Jurassic section, including the Praia da Amoreira-Porto Novo and Bombarral Formations, crops out north of the town of Peniche (Manuppella et al., 1999; Azerêdo et al., 2010) (Figure 10). Some stratigraphic approaches have suggested the presence of the Sobral Fm. along this sequence (e.g., Hill, 1988; Martinius and Gowland, 2011) (Figure 10.2). This section is not very large (its maximum length is about 2 km), but it is rich in fossil sites, some of them known since the first half of the twentieth century. The most important dinosaur site is located in Pedras Muitas (Peniche) where remains of sauropods and stegosaurs have been collected (Zbyszewski, 1946; Lapparent and Zbyszewski, 1957). Their precise location is not clear. According to the available stratigraphic information (e.g., Lapparent and Zbyszewski, 1957; Camarate França et al., 1960; Hill, 1988; Bernardes, 1992; Schneider et al., 2009; Azerêdo et al., 2010; Martinius and Gowland, 2011; Taylor et al., 2014), these elements probably came from sediments of the Praia da Amoreira-Porto Novo Fm. Some partial middle and posterior sauropod cervical vertebrae (MG 4915, 4916, 4917 and 4919) were found in Pedras Muitas. These remains include a misidentified cervical vertebra, previously considered part of an Omosaurus individual (Zbyszewski, 1946, pl. I, figure 1). This specimen includes middle and posterior cervical vertebrae (Figure 11.5-6), which have been subject to a systematic revision and might represent a neosauropod form due the presence of polycamerate cervical vertebrae with bifurcated neural spine.
North Peniche (Figure 10, Figure 11). An Upper Jurassic section, including the Praia da Amoreira-Porto Novo and Bombarral Formations, crops out north of the town of Peniche (Manuppella et al., 1999; Azerêdo et al., 2010) (Figure 10). Some stratigraphic approaches have suggested the presence of the Sobral Fm. along this sequence (e.g., Hill, 1988; Martinius and Gowland, 2011) (Figure 10.2). This section is not very large (its maximum length is about 2 km), but it is rich in fossil sites, some of them known since the first half of the twentieth century. The most important dinosaur site is located in Pedras Muitas (Peniche) where remains of sauropods and stegosaurs have been collected (Zbyszewski, 1946; Lapparent and Zbyszewski, 1957). Their precise location is not clear. According to the available stratigraphic information (e.g., Lapparent and Zbyszewski, 1957; Camarate França et al., 1960; Hill, 1988; Bernardes, 1992; Schneider et al., 2009; Azerêdo et al., 2010; Martinius and Gowland, 2011; Taylor et al., 2014), these elements probably came from sediments of the Praia da Amoreira-Porto Novo Fm. Some partial middle and posterior sauropod cervical vertebrae (MG 4915, 4916, 4917 and 4919) were found in Pedras Muitas. These remains include a misidentified cervical vertebra, previously considered part of an Omosaurus individual (Zbyszewski, 1946, pl. I, figure 1). This specimen includes middle and posterior cervical vertebrae (Figure 11.5-6), which have been subject to a systematic revision and might represent a neosauropod form due the presence of polycamerate cervical vertebrae with bifurcated neural spine.
 Other sauropod remains were also identified to the north of Peniche. Most of these specimens are deposited in the collections of the MG, ML and SHN. A currently unpublished, partial spatulate tooth (Figure 11.1, MG 8783), bearing lingual facets, was found in Baleal. It can be attributed to an indeterminate eusauropod, possibly a basal macronarian. This tooth morphology is common in mamenchisaurids (Ouyang and Ye, 2002), basal macronarians (Osborn and Mook, 1921; Gilmore, 1925; Ostrom and McIntosh, 1966), and in the euhelopodid Euhelopus (Wilson and Upchurch, 2009). Heart-shaped teeth (Figure 11.2-3) are also reported from this area (Mocho et al., 2012, 2016b), which indicate the presence of turiasaurian eusauropods in the outcropping sediments north of Peniche. The SHN houses several specimens from Baleal, Pedras Muitas, and Almagreira (Figure 10.3), which still need preparation, including several axial and appendicular elements (e.g., Figure 11.8-10). One of those specimens is an anterior caudal vertebra with a slightly procoelous centrum (Figure 11.9, SHN 180). The overall morphology (e.g., slightly procoelous centrum and short neural spine with distal rugosities) resembles that of the anterior caudal vertebrae collected from the Spanish sediments of the Villar de Arzobispo Formation, in San Lorenzo (Riodeva), attributed by Cobos et al. (2011) to Turiasauria. The procoelous condition is shared with several clades within Eusauropoda (e.g., Wilson, 2002; Upchurch et al., 2004). Some compressed cone-chisel-shaped teeth (Figure 11.4, 11.7) collected in the Baleal-Pedras Muitas coastal section are deposited in the SHN collections.
Other sauropod remains were also identified to the north of Peniche. Most of these specimens are deposited in the collections of the MG, ML and SHN. A currently unpublished, partial spatulate tooth (Figure 11.1, MG 8783), bearing lingual facets, was found in Baleal. It can be attributed to an indeterminate eusauropod, possibly a basal macronarian. This tooth morphology is common in mamenchisaurids (Ouyang and Ye, 2002), basal macronarians (Osborn and Mook, 1921; Gilmore, 1925; Ostrom and McIntosh, 1966), and in the euhelopodid Euhelopus (Wilson and Upchurch, 2009). Heart-shaped teeth (Figure 11.2-3) are also reported from this area (Mocho et al., 2012, 2016b), which indicate the presence of turiasaurian eusauropods in the outcropping sediments north of Peniche. The SHN houses several specimens from Baleal, Pedras Muitas, and Almagreira (Figure 10.3), which still need preparation, including several axial and appendicular elements (e.g., Figure 11.8-10). One of those specimens is an anterior caudal vertebra with a slightly procoelous centrum (Figure 11.9, SHN 180). The overall morphology (e.g., slightly procoelous centrum and short neural spine with distal rugosities) resembles that of the anterior caudal vertebrae collected from the Spanish sediments of the Villar de Arzobispo Formation, in San Lorenzo (Riodeva), attributed by Cobos et al. (2011) to Turiasauria. The procoelous condition is shared with several clades within Eusauropoda (e.g., Wilson, 2002; Upchurch et al., 2004). Some compressed cone-chisel-shaped teeth (Figure 11.4, 11.7) collected in the Baleal-Pedras Muitas coastal section are deposited in the SHN collections.
The systematic affinities of the sauropod remains collected north of Peniche still need to be established in a detailed study. However, the currently available information allows the identification of possible turiasaurs and basal macronarians, including members of the clade that includes Titanosauriformes and Europasaurus (Figure 10).
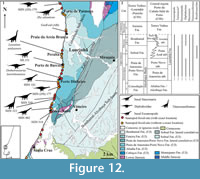 Praia da Consolação-Lourinhã-Torres Vedras coastal sector (Figure 12, Figure 13, Figure 14, Figure 15). This is the richest area in the Lusitanian Basin concerning Upper Jurassic dinosaur remains (e.g., Lapparent and Zbyszewski, 1957; Dantas, 1990; Antunes and Mateus, 2003; Ortega et al., 2009, 2013) (Figure 12.1). A thick Upper Jurassic sedimentary sequence crops out in this sector and includes deposits of the Praia da Amoreira-Porto Novo, Sobral and Bombarral Formations. This continental sedimentary sequence was deposited above the marine Abadia Fm. (Manuppella et al., 1999) (Figure 12.2). The type specimens of Lusotitan atalaiensis, Dinheirosaurus lourinhanensis and Zby atlanticus (Lapparent and Zbyszewski, 1957; Dantas et al., 1992; Bonaparte and Mateus, 1999; Antunes and Mateus, 2003; Mannion et al., 2012, 2013; Mateus et al., 2014) were found in this area of the Bombarral Sub-basin (Consolação Sub-basin following Taylor et al., 2014). In addition, many published and unpublished specimens, most of them housed in the palaeontological collections of the MG, ML, and SHN, were also collected in this area (Bonaparte and Mateus, 1999; Mateus, 2005; Yaguë et al., 2006; Mannion et al., 2012; Mocho et al., 2013a, 2013b, 2014a).
Praia da Consolação-Lourinhã-Torres Vedras coastal sector (Figure 12, Figure 13, Figure 14, Figure 15). This is the richest area in the Lusitanian Basin concerning Upper Jurassic dinosaur remains (e.g., Lapparent and Zbyszewski, 1957; Dantas, 1990; Antunes and Mateus, 2003; Ortega et al., 2009, 2013) (Figure 12.1). A thick Upper Jurassic sedimentary sequence crops out in this sector and includes deposits of the Praia da Amoreira-Porto Novo, Sobral and Bombarral Formations. This continental sedimentary sequence was deposited above the marine Abadia Fm. (Manuppella et al., 1999) (Figure 12.2). The type specimens of Lusotitan atalaiensis, Dinheirosaurus lourinhanensis and Zby atlanticus (Lapparent and Zbyszewski, 1957; Dantas et al., 1992; Bonaparte and Mateus, 1999; Antunes and Mateus, 2003; Mannion et al., 2012, 2013; Mateus et al., 2014) were found in this area of the Bombarral Sub-basin (Consolação Sub-basin following Taylor et al., 2014). In addition, many published and unpublished specimens, most of them housed in the palaeontological collections of the MG, ML, and SHN, were also collected in this area (Bonaparte and Mateus, 1999; Mateus, 2005; Yaguë et al., 2006; Mannion et al., 2012; Mocho et al., 2013a, 2013b, 2014a).
The Praia da Amoreira-Porto Novo Formation crops out in several areas along this sector, especially in the sedimentary sections from São Bernardino to Paimogo and from Porto Dinheiro to Praia de Santa Rita (Hill, 1988; Manuppella et al., 1999; Mateus et al., 2013). This late Kimmeridgian-basal Tithonian formation (Fürsich, 1981; Manuppella et al., 1999) is well-known for its abundant dinosaur fossil remains, including several sauropod specimens (Lapparent and Zbyszewski, 1957; Dantas, 1990; Manuppella et al., 1999; Antunes and Mateus, 2003; Ortega et al., 2009). This formation is interpreted as having been deposited in a distal alluvial to fluvial meandriform environment (Hill, 1988, 1989).
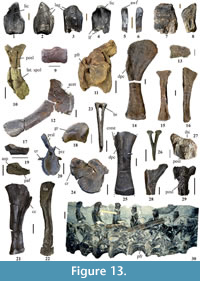 The holotype of Zby atlanticus (Figure 13.13-16, ML 368) was found in the Vale de Pombas cliffs (north of Forte de Paimogo), in the Praia da Amoreira-Porto Novo Formation. This partial skeleton includes a tooth, a chevron, a right partial scapula, and coracoid, an almost complete right forelimb, and some indeterminate elements (Mateus, 2005, 2009; Mateus et al., 2014; pers. observ. PM). The material of Zby atlanticus was first attributed to the North American Morrison genus Camarasaurus (Mateus, 2005), and subsequently to Turiasaurus riodevensis (Mateus, 2009). Mateus et al. (2014) described the species Zby atlanticus as a member of Turiasauria. The affinities between Zby and the members of the Turiasauria clade were previously noted by other authors (Mateus, 2009; Mocho et al., 2012; Royo-Torres and Upchurch, 2012).
The holotype of Zby atlanticus (Figure 13.13-16, ML 368) was found in the Vale de Pombas cliffs (north of Forte de Paimogo), in the Praia da Amoreira-Porto Novo Formation. This partial skeleton includes a tooth, a chevron, a right partial scapula, and coracoid, an almost complete right forelimb, and some indeterminate elements (Mateus, 2005, 2009; Mateus et al., 2014; pers. observ. PM). The material of Zby atlanticus was first attributed to the North American Morrison genus Camarasaurus (Mateus, 2005), and subsequently to Turiasaurus riodevensis (Mateus, 2009). Mateus et al. (2014) described the species Zby atlanticus as a member of Turiasauria. The affinities between Zby and the members of the Turiasauria clade were previously noted by other authors (Mateus, 2009; Mocho et al., 2012; Royo-Torres and Upchurch, 2012).
Lapparent and Zbyszewski (1957) reported the presence of some specimens collected north of the Forte de Paimogo (Praia da Amoreira-Porto Novo Formation). Non-sauropod dinosaurs were also found in this locality (Lapparent and Zbyszewski, 1957; Escaso et al., 2008; Hendrickx and Mateus, 2014). A partial caudal series (MG 4978), with 15 caudal vertebrae, was found in São Bernardino (Peniche). A partial right humerus (Figure 13.25, MG 4976) of a sauropod was found in Praia dos Frades (Peniche) and was attributed to Apatosaurus alenquerensis (Lapparent and Zbyszewski, 1957). This humerus has a crest on the posterior face of the proximal end, which is here identified as a shared feature with the humerus of Duriatitan humerocristatus, from the British lower Kimmeridge Clay Fm. (Barrett et al., 2010), being considered as cf. Duriatitan humerocristatus (Mocho et al., 2016a). An unpublished metacarpal I (SHN 583) found in Praia dos Frades (Peniche) shares the morphology of metacarpal I of Turiasaurus riodevensis and Zby atlanticus with the presence of a rough platform on the laterodorsal border of the distal end. It is thus tentatively considered as cf. Turiasauria.
A so far unprepared diplodocid specimen (Figure 13.27, SHN (JJS) 179) from Praia da Vermelha (Peniche, Praia da Amoreira-Porto Novo Formation), comprises axial and appendicular elements (Peniche; Mocho et al., 2014a). The morphology of the neural spines (posterior dorsal or anterior caudal), with a well-defined prespinal lamina, a rectangular shape (anteroposteriorly compressed), and with a slight dorsal bifurcation, is similar to the apomorphic morphology in the posterior dorsal and anterior caudal neural spines of diplodocids, such as Supersaurus, Dinheirosaurus, Diplodocus, and Barosaurus (e.g., Hatcher, 1901; Lull, 1919; McIntosh, 2005; Mannion et al., 2012; pers. observ., PM). Other unprepared sauropod specimens from this sector, also from the Praia da Amoreira-Porto Novo Fm., are housed in the collections of the SHN and ML (see list of Mateus, 2005, for ML specimens; pers. observ., PM). Several teeth have also been found between Forte de Paimogo and Praia da Consolação, including heart-shaped teeth, probably referable to Turiasauria (Mocho et al., 2012, 2016b), spatulate teeth (e.g., SHN 516, SHN 540), attributed to indeterminate eusauropods, probably basal macronarians, and compressed cone-chisel-shaped teeth (e.g., SHN 546), a morphology shared with titanosauriforms and the basal macronarian Europasaurus. A skull fragment bearing heart-shaped teeth was found in Praia dos Frades (Figure 13.7-8, SHN 582) and probably represents a turiasaurian sauropod.
To the south of Porto Dinheiro, the type locality of Dinheirosaurus lourinhanensis (Dantas et al., 1992; Bonaparte and Mateus, 1999; Mannion et al., 2012), an extensive section of the Praia da Amoreira-Porto Novo Formation crops out. The border between the municipalities of Torres Vedras and Lourinhã is located in this relatively poorly prospected area. New discoveries reveal a rich dinosaur fauna, including theropods, ornithopods, thyreophorans, and sauropods, as well as other vertebrate groups, such as turtles and crocodyliforms (e.g., Malafaia et al., 2008; Escaso et al., 2010a, 2010b; Pérez-García and Ortega, 2011). Several fossil sites containing sauropods were found in this area, including some partial skeletons.
 The first remains of the holotype of Dinheirosaurus lourinhanensis were identified in 1987, in the cliffs of Porto Dinheiro. Consequently, in 1988 and 1991, a team composed of members of the Museu Nacional de História Natural e da Ciência (Lisboa, Portugal), Salamanca University (Salamanca, Spain) and GEAL (Lourinhã, Portugal) proceeded to the extraction of a series of partially articulated cervical and dorsal vertebrae, with associated dorsal ribs, as well as caudal vertebrae and pelvic elements (Figure 13.28-30, Figure 14.1) (Dantas et al., 1992). Bonaparte and Mateus (1999) defined a new diplodocid taxon, Dinheirosaurus alenquerensis, considered as a diplodocine form closely related to the North American Late Jurassic Supersaurus (Rauhut et al., 2005; Whitlock, 2011; Mannion et al., 2012; Tschopp and Mateus, 2013; Tschopp et al., 2015). In fact, Tschopp et al. (2015) suggested that Dinheirosaurus lourinhanensis might represent a species of Supersaurus, proposing the new combination Supersaurus lourinhanensis. Several remains of the type specimen (ML 414), including several caudal vertebrae (pers. observ., FO and PM), need to be prepared for the confirmation or refutation of this hypothesis. The Dinheirosaurus type locality is stratigraphically close to the boundary between the Praia da Amoreira-Porto Novo and Sobral Formations (Manuppella et al., 1999; field observations, PM).
The first remains of the holotype of Dinheirosaurus lourinhanensis were identified in 1987, in the cliffs of Porto Dinheiro. Consequently, in 1988 and 1991, a team composed of members of the Museu Nacional de História Natural e da Ciência (Lisboa, Portugal), Salamanca University (Salamanca, Spain) and GEAL (Lourinhã, Portugal) proceeded to the extraction of a series of partially articulated cervical and dorsal vertebrae, with associated dorsal ribs, as well as caudal vertebrae and pelvic elements (Figure 13.28-30, Figure 14.1) (Dantas et al., 1992). Bonaparte and Mateus (1999) defined a new diplodocid taxon, Dinheirosaurus alenquerensis, considered as a diplodocine form closely related to the North American Late Jurassic Supersaurus (Rauhut et al., 2005; Whitlock, 2011; Mannion et al., 2012; Tschopp and Mateus, 2013; Tschopp et al., 2015). In fact, Tschopp et al. (2015) suggested that Dinheirosaurus lourinhanensis might represent a species of Supersaurus, proposing the new combination Supersaurus lourinhanensis. Several remains of the type specimen (ML 414), including several caudal vertebrae (pers. observ., FO and PM), need to be prepared for the confirmation or refutation of this hypothesis. The Dinheirosaurus type locality is stratigraphically close to the boundary between the Praia da Amoreira-Porto Novo and Sobral Formations (Manuppella et al., 1999; field observations, PM).
Another partial diplodocid individual was found in Valmitão (Lourinhã), south of Porto Dinheiro (Mocho et al., 2014a). This specimen and the Dinheirosaurus holotype are the most complete diplodocids of the European Upper Jurassic record. The Valmitão specimen (SHN (JJS) 177) comprises elements of the axial skeleton (dorsal?, sacral, and anterior caudal vertebrae; ribs and chevrons) and pelvic girdle (ilia, ischia, and pubis) (Figure 13.10-12). SHN (JJS) 177 can be referred to Flagellicaudata on the basis of the presence of an expanded distal end of the ischia (following Whitlock, 2011). Rectangular anterior caudal neural spines in anterior view and the presence of diapophyseal laminae on the anterior caudal ribs support the assignation of SHN (JJS) 177 to Diplodocidae (sensu Whitlock, 2011). The presence of wing-like, anterior caudal ribs, dorsal concavity in the neural spines (slightly bifurcated), and ventral and lateral pneumaticity, suggest that the SHN (JJS) 177 is closely related to diplodocines, such as Diplodocus, Barosaurus, and Tornieria (Osborn, 1899; Hatcher, 1901; Lull, 1919; McIntosh, 2005; Remes, 2006; Whitlock, 2011; Tschopp et al., 2015).
Many sauropod specimens from the coastal cliffs between Porto Dinheiro and Santa Rita are housed in the SHN, and most still require preparation. Mateus (2005) also reported an appreciable number of specimens from this area and deposited in the ML. In 2003 and 2009, the SHN excavated a partial and articulated skeleton (Figure 14.2-3) in Santa Rita (Torres Vedras), including a partial tail and pelvis, associated with limb bones (Figure 13.26, SHN 534). The systematic affinities of this specimen still need to be clarified. Close to this quarry is the type locality of the pleurosternid turtle Selenemys lusitanica, also from the Praia da Amoreira-Porto Novo Formation (Pérez-García and Ortega, 2011). Another fossil site prospected by the SHN is located in Porto Novo (Torres Vedras). Several axial and appendicular sauropod bones were recovered there. This specimen (SHN 002) shares the general morphology of the forelimb bones with the camarasaurid Lourinhasaurus alenquerensis; nevertheless, no diagnostic features can be identified. These two fossil sites are stratigraphically close to the upper boundary of the Praia da Amoreira-Porto Novo Fm. with the Sobral Fm. (Manuppella et al., 1999; field observations, PM). Another specimen (SHN 530), including sacral and caudal vertebrae, and appendicular bones (Figure 13.23-24), was found in the cliffs of Praia da Corva (Torres Vedras). The anterior caudal vertebrae (Figure 13.24) resemble the morphology present in Iberian turiasaurs (e.g., Casanovas et al., 2001), with slightly procoelous centra and fan-like caudal ribs, lacking the lateral and ventral pneumaticity and dorsalised neural spine, which would be typical of diplodocids (e.g., Hatcher, 1901; McIntosh, 2005; Remes, 2006). This specimen also preserves long and bridged anterior chevrons. An unpublished set of anterior, middle, and posterior caudal vertebrae (Figure 13.9), probably belonging to a single individual, was also found at Praia da Corva (SHN 523). It might represent an indeterminate titanosauriform, based on the presence of anteriorly displaced neural arches, pneumatic fossae (as occur in Giraffatitan or Andesaurus, Mannion and Calvo, 2011; Wedel and Taylor, 2013), and dorsoventrally compressed centrum. Finally, an also unpublished medium-sized individual (SHN 181) found in Valmitão is currently being prepared and described. It includes caudal vertebrae, and pectoral, pelvic, and hindlimb elements (Figure 13.19-22). This specimen presents several peculiar features and probably represents a new sauropod taxon. Furthermore, several teeth belonging to different morphotypes were recovered from the area between Porto Dinheiro and Santa Rita. These specimens include spatulate (Figure 13.3-4, SHN 513), and heart- (Figure 13.1-2, see Mocho et al., 2012, 2016b), and compressed cone-chisel-shaped teeth (Figure 13.5-6, SHN 574, 575, 578).
 The Sobral Formation is a lateral equivalent of the lowest part of the Bombarral Fm., representing a regional transgression (e.g., Hill, 1988). The Sobral Fm. was deposited in a deltaic to marginal marine environment (Hill, 1988; Manuppella et al., 1999). In the western part of the Bombarral Sub-basin (i.e., Consolação Sub-basin, following Taylor et al., 2014), this formation crops out in the coastal section from Peralta to Porto Dinheiro. Several sauropod specimens from the Sobral Fm. were reported, most notably the lectotype of Lusotitan atalaiensis. It was found in the Peralta cliffs, close to the Atalaia locality (Lourinhã). Lapparent and Zbyszewski (1957) originally considered this specimen to represent a new species of Brachiosaurus (at that time the genus Brachiosaurus included two species, the North American B. altithorax and the African B. brancai). Subsequently, Antunes and Mateus (2003) established the new genus, Lusotitan, to denominate this specimen. Mannion et al. (2013) presented a systematic revision of the Peralta specimen, considering it a basal macronarian, and, possibly, a brachiosaurid. The brachiosaurid affinities were recently supported by Mocho et al. (2016c). This specimen includes dorsal, sacral, and caudal vertebrae, dorsal ribs, chevrons, pelvic, forelimb, and hindlimb elements (Figure 15.9-14; Lapparent and Zbyszewski, 1957; Antunes and Mateus, 2003; Mannion et al., 2013; Mocho et al., 2016c). Several sauropod teeth were found at Peralta, including spatulate (Figure 15.1-2, Mocho et al., 2011) and heart- (Mocho et al., 2012, 2016b), and compressed cone-chisel-shaped teeth (Figure 15.3-6). Some isolated bones were also found in this region (Figure 15.7).
The Sobral Formation is a lateral equivalent of the lowest part of the Bombarral Fm., representing a regional transgression (e.g., Hill, 1988). The Sobral Fm. was deposited in a deltaic to marginal marine environment (Hill, 1988; Manuppella et al., 1999). In the western part of the Bombarral Sub-basin (i.e., Consolação Sub-basin, following Taylor et al., 2014), this formation crops out in the coastal section from Peralta to Porto Dinheiro. Several sauropod specimens from the Sobral Fm. were reported, most notably the lectotype of Lusotitan atalaiensis. It was found in the Peralta cliffs, close to the Atalaia locality (Lourinhã). Lapparent and Zbyszewski (1957) originally considered this specimen to represent a new species of Brachiosaurus (at that time the genus Brachiosaurus included two species, the North American B. altithorax and the African B. brancai). Subsequently, Antunes and Mateus (2003) established the new genus, Lusotitan, to denominate this specimen. Mannion et al. (2013) presented a systematic revision of the Peralta specimen, considering it a basal macronarian, and, possibly, a brachiosaurid. The brachiosaurid affinities were recently supported by Mocho et al. (2016c). This specimen includes dorsal, sacral, and caudal vertebrae, dorsal ribs, chevrons, pelvic, forelimb, and hindlimb elements (Figure 15.9-14; Lapparent and Zbyszewski, 1957; Antunes and Mateus, 2003; Mannion et al., 2013; Mocho et al., 2016c). Several sauropod teeth were found at Peralta, including spatulate (Figure 15.1-2, Mocho et al., 2011) and heart- (Mocho et al., 2012, 2016b), and compressed cone-chisel-shaped teeth (Figure 15.3-6). Some isolated bones were also found in this region (Figure 15.7).
Porto das Barcas is another relevant locality concerning dinosaur remains. The holotype of the ornithopod Eousdryosaurus (Dantas et al., 2000; Escaso et al., 2014) and nests with theropods eggs tentatively referred to Torvosaurus (Castanhinha et al., 2009; Araújo et al., 2013), were found there. Several sauropod specimens from Porto das Barcas were referred to Apatosaurus alenquerensis by Lapparent and Zbyszewski (1957), including caudal vertebrae (MG 8800, 8805). MG 30390 is an unpublished partial skeleton that represents an indeterminate eusauropod, according to the presence of procoelous anterior caudal vertebrae, which are only known in Eusauropoda, such as mamenchisaurids, turiasaurs, diplodocids, and titanosaurs (e.g., Salgado, 1997; Wilson, 2002; Upchurch et al., 2004; Carballido and Sander, 2014). Another partial skeleton from Porto das Barcas is housed in the ML (ML 351), including a partial caudal series, sacrum and fibula (Antunes and Mateus, 2003; Mateus, 2005). Antunes and Mateus (2003) and Mateus (2005) referred this specimen to Lourinhasaurus alenquerensis, but this specimen does not bear diagnostic features that support this taxonomic assignment. The detailed preparation and study of this material is in process (RC, personal commun., 2015). Other specimens from this locality are housed in the ML and SHN (Mateus, 2005, pers. observ, PM), including heart-shaped (Mocho et al., 2012, 2016b), and compressed cone-chisel-shaped teeth (e.g., SHN 576). From Lage Fria (Porto das Barcas), Tschopp, and Mateus (2012) described a sternal plate (ML 684), suggesting that it might pertain to Turiasaurus riodevensis or Lusotitan atalaiensis. Lapparent and Zbyszewski (1957, pl. XIII, figure 31-33) identified one vertebra from Porto das Barcas (MMLT 602528) as a posterior dorsal vertebra of Megalosaurus pombali. However, it represents an anterior caudal vertebra of an indeterminate sauropod. Another sauropod caudal vertebra (MMLT 602529) from Porto das Barcas was found in the collections of the MMLT, and probably corresponds to a vertebra mentioned by Lapparent and Zbyszewski (1957, pg. 38). This vertebra represents an indeterminate sauropod.
The Tithonian Bombarral Formation crops out in the coastal section from Praia da Areia Branca to Paimogo (including Vale de Frades) (Figure 12.1). This formation lies above the Sobral Fm. (on the Forte de Paimogo locality) and Praia da Amoreira-Porto Novo Fm. (Hill, 1988; Manuppella et al., 1999). Furthermore, the Bombarral Fm. crops out in a wide area to the east of the Lourinhã Fault, from A-dos-Cunhados to Alcobaça (an area included in the Bombarral-Alcobaça Sub-basin by Taylor et al., 2014). The Bombarral Fm. was deposited in a coastal lagoonal to meandering fluvial environment (Manuppella et al., 1999).
West of the Lourinhã Fault, the Bombarral Formation crops out between Peralta and Vale de Frades (south of Paimogo), and from south of Santa Rita to the town of Santa Cruz. Some bones from the Bombarral Fm. were reported by Lapparent and Zbyszewski (1957). A partial left femur (Figure 15.15, MG 4986) was found in Praia da Areia Branca and was originally referred to Brachiosaurus atalaiensis (Lapparent and Zbyszewski, 1957). It has recently been reinterpreted as an indeterminate titanosauriform (Mannion et al., 2013). Two caudal vertebrae housed in the palaeontological collections of the Museu Nacional de História Natural e da Ciência (Lisboa) were recovered in Areia Branca: an anterior caudal vertebra previously referred to Brachiosaurus atalaiensis (Lapparent and Zbyszewski, 1957), and a procoelous anterior caudal centrum (Figure 15.8, MNHNUL.P.R27) referred to Apatosaurus alenquerensis (Lapparent and Zbyszewski, 1957). The first element can only be identified as an indeterminate sauropod. The second bone is recognised as belonging to an indeterminate eusauropod, based on the procoelous condition, a feature present in several eusauropod groups (e.g., Salgado et al., 1997; Wilson, 2002; Upchurch et al., 2004; Carballido and Sander, 2014). Lapparent and Zbyszewski (1957, pg. 17) reported the occurrence of a small femur from a sauropod from Vale de Frades, but its present whereabouts are unknown.
Yagüe et al. (2006) published a partial tail found in Praia de Areia Branca (GeoFCUL (AB), Lourinhã), recovered by members of the Department of Geology of the Sciences Faculty of Lisboa University (GeoFCUL). This specimen comprises several middle caudal vertebrae and chevrons (Figure 15.16-19), and was tentatively attributed to a basal macronarian (Yagüe et al., 2006). Comparisons with more complete caudal series (e.g., Osborn and Mook, 1921; McIntosh et al., 1996a, 1996b; Mannion et al., 2013) indicate that the most anterior preserved vertebra probably corresponds to the thirteenth or fourteenth caudal element. The reduction of the caudal ribs before reaching the twentieth caudal vertebra is common in eusauropods (Upchurch et al., 2004). The absence of middle chevrons without anterior and posterior process led Yagüe et al. (2006) to refer this sauropod to Macronaria. Nevertheless, this feature is also present in the non-neosauropod Jobaria (Sereno et al., 1999). The available elements do not present features supporting its placement within Macronaria, so that we prefer an identification of this material as Eusauropoda indet. A procoelous anterior caudal vertebra (MMPM.P/73) of an indeterminate eusauropod was collected in Paimogo (Mocho et al., 2016b). No precise location was given for this element. As a result, and taking into account that in this locality sediments of both the Bombarral and Sobral Formations crop out (Hill, 1988; Mannuppella et al., 1999), its precise stratigraphic context is not known. An important section of the Bombaral Fm. crops out south of Santa Rita and several unpublished elements were collected there, including some presacral vertebrae preserved in the SHN (e.g., SHN 006, 535).
In conclusion, the Praia da Consolação-Lourinhã-Torres Vedras coastal sector presents the most diverse Portuguese Upper Jurassic fossil record for sauropods, including non-neosauropod eusauropods, such as turiasaurs (e.g., Zby atlanticus), diplodocines (e.g., Dinheirosaurus lourinhanensis and SHN (JJS) 177), basal macronarians (e.g., SHN 181), and titanosauriforms, including brachiosaurids (e.g., Lusotitan atalaiensis). The presence of camarasaurid macronarians cannot be confirmed in this area, but the systematic study of SHN 002 will allow the testing of its affinities with Lourinhasaurus alenquerensis and other camarasaurids.
Arruda Sub-Basin
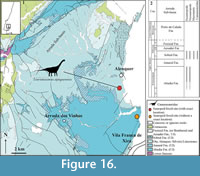 The Arruda Sub-basin is located in the SE part of the central sector of the Lusitanian Basin and has few known dinosaur fossil sites (Figure 16, Figure 17), a fact that may in part be related to the absence of a wide area of exposed Upper Jurassic outcrops. This half-graben basin has a similar Upper Jurassic stratigraphy to that of the Turcifal Sub-basin (Kullberg et al., 2006, 2013; Figure 2). The vertebrate fossil record from this sub-basin is mainly composed of sauropod dinosaurs (Lapparent and Zbyszewski, 1957; Antunes and Mateus, 2003; Ortega et al., 2009).
The Arruda Sub-basin is located in the SE part of the central sector of the Lusitanian Basin and has few known dinosaur fossil sites (Figure 16, Figure 17), a fact that may in part be related to the absence of a wide area of exposed Upper Jurassic outcrops. This half-graben basin has a similar Upper Jurassic stratigraphy to that of the Turcifal Sub-basin (Kullberg et al., 2006, 2013; Figure 2). The vertebrate fossil record from this sub-basin is mainly composed of sauropod dinosaurs (Lapparent and Zbyszewski, 1957; Antunes and Mateus, 2003; Ortega et al., 2009).
One of the most complete sauropod individuals from the European Upper Jurassic record was discovered in the Arruda Sub-basin. The sole specimen identified was used to define a new species of Apatosaurus: Apatosaurus alenquerensis (Lapparent and Zbyszewski, 1957). Dantas et al. (1998) proposed a new genus for this taxon, Lourinhasaurus, resulting in the new combination Lourinhasaurus alenquerensis. The specimen includes cervical, dorsal, sacral, and anterior caudal vertebrae, dorsal and sacral ribs, scapular and pelvic elements, and bones of the hindlimb and forelimb (Figure 17.1-6, 17.8-17; Lapparent and Zbyszewski, 1957; Mocho et al., 2014b). The recent systematic revision of the Lourinhasaurus lectotype confirmed the placement of this taxon within Camarasauridae (Mocho et al., 2014b), a hypothesis previously suggested by other authors (McIntosh, 1990a, 1990b; Wilson and Sereno, 1998). A partial opisthocoelous dorsal centrum (Figure 17.7, MG 4799) was found in Castanheira (Vila Franca de Xira), in coarse sandy arkosic sediments of the Abadia Formation (Leinfelder and Wilson, 1989). This element was originally described as a metatarsal V by Lapparent and Zbyszewski (1957). There are some unpublished fossils deposited in the MG collections that might come from Alenquer or even from Moinho do Carmo, but these specimens show different preservation and taxonomic features when compared with the material of the Lourinhasaurus lectotype. This raises doubt about their locality of origin.
Lapparent and Zbyszewski, 1957; Mocho et al., 2014b). The recent systematic revision of the Lourinhasaurus lectotype confirmed the placement of this taxon within Camarasauridae (Mocho et al., 2014b), a hypothesis previously suggested by other authors (McIntosh, 1990a, 1990b; Wilson and Sereno, 1998). A partial opisthocoelous dorsal centrum (Figure 17.7, MG 4799) was found in Castanheira (Vila Franca de Xira), in coarse sandy arkosic sediments of the Abadia Formation (Leinfelder and Wilson, 1989). This element was originally described as a metatarsal V by Lapparent and Zbyszewski (1957). There are some unpublished fossils deposited in the MG collections that might come from Alenquer or even from Moinho do Carmo, but these specimens show different preservation and taxonomic features when compared with the material of the Lourinhasaurus lectotype. This raises doubt about their locality of origin.
The scarce record of the Arruda Sub-basin only allows the identification of an indeterminate neosauropod and a camarasaurid macronarian (Lourinhasaurus alenquerensis) preserved in sediments of the Abadia and the Sobral Formations, respectively (Figure 16.1-2).
Turcifal Sub-basin
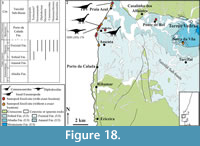 The Turcifal Sub-basin is located west of the Arruda Sub-basin, in the SW part of the central sector, corresponding to distal palaeoenvironments, but showing abundant deltaic inputs from the west. The stratigraphy of both sub-basins is similar (Kullberg et al., 2006, 2013). Several authors provided recent updates on the stratigraphy of this area (e.g., Pereda-Suberbiola et al., 2005; Kullberg et al., 2006, 2013). Its Upper Jurassic outcrops are not as extensive as those of the Bombarral Sub-basin, but dinosaur fossil sites are particularly abundant in the coastal sector (Figure 18, Figure 19). A diverse dinosaur fauna, composed of sauropods, theropods, thyreophorans, and ornithopods, has been identified. This sub-basin is bordered in the north by the Torres Vedras-Montejunto Fault and in the east by the Runa Fault, which separates it from the Arruda Sub-basin (e.g., Wilson, 1988; Figure 2).
The Turcifal Sub-basin is located west of the Arruda Sub-basin, in the SW part of the central sector, corresponding to distal palaeoenvironments, but showing abundant deltaic inputs from the west. The stratigraphy of both sub-basins is similar (Kullberg et al., 2006, 2013). Several authors provided recent updates on the stratigraphy of this area (e.g., Pereda-Suberbiola et al., 2005; Kullberg et al., 2006, 2013). Its Upper Jurassic outcrops are not as extensive as those of the Bombarral Sub-basin, but dinosaur fossil sites are particularly abundant in the coastal sector (Figure 18, Figure 19). A diverse dinosaur fauna, composed of sauropods, theropods, thyreophorans, and ornithopods, has been identified. This sub-basin is bordered in the north by the Torres Vedras-Montejunto Fault and in the east by the Runa Fault, which separates it from the Arruda Sub-basin (e.g., Wilson, 1988; Figure 2).
Theropods are represented by allosaurids, with an important specimen coming from Cambelas (Malafaia et al., 2009b; Ortega et al., 2009). Thyreophoran dinosaurs are represented in this sub-basin by the holotype of the ankylosaur Dracopelta zbyszewskii (Galton, 1980; Pereda-Suberbiola et al., 2005), found in sediments of the Freixial Formation in Assenta (Torres Vedras). Another partial ankylosaur skeleton was found in the Porto Calada locality (Mafra), and tentatively identified as a ‘polacanthid’ nodosaurid (Mateus et al., 2013). Material referred to the stegosaur Dacentrurus was also described from the Moçafaneira locality (Escaso et al., 2007b).
Non-dinosaur vertebrates are also present in the sediments of this basin. Among them, the turtle record is very significant, including the type material of “Plesiochelys” choffati and Hylaeochelys kappa (Sauvage, 1897-98; Pérez-García et al., 2008; Pérez-García and Ortega, 2013; Pérez-García, 2015). The record of crocodyliforms is also abundant, represented not only by osteological elements, but also by a nest with eggs, found in the Cambelas area (Russo et al. 2014a, 2014b, 2017).
 Sauropods are well-represented in the Turcifal Sub-basin. Most of the record of this clade is from recent findings, and so relatively little information is available yet, because most of these specimens are currently under study and/or in preparation. A short section of the Praia da Amoreira-Porto Novo Formation (fluvial environment) is found south of the Santa Cruz locality. A middle caudal vertebra (Figure 19.10, MG 4802) was collected in this area and was attributed to Apatosaurus alenquerensis (Lapparent and Zbyszewski, 1957). This vertebra is here considered to represent an indeterminate sauropod due to the absence of diagnostic characters that would allow a more detailed taxonomic assignment.
Sauropods are well-represented in the Turcifal Sub-basin. Most of the record of this clade is from recent findings, and so relatively little information is available yet, because most of these specimens are currently under study and/or in preparation. A short section of the Praia da Amoreira-Porto Novo Formation (fluvial environment) is found south of the Santa Cruz locality. A middle caudal vertebra (Figure 19.10, MG 4802) was collected in this area and was attributed to Apatosaurus alenquerensis (Lapparent and Zbyszewski, 1957). This vertebra is here considered to represent an indeterminate sauropod due to the absence of diagnostic characters that would allow a more detailed taxonomic assignment.
Most of the sauropod discoveries in the Turcifal Sub-basin come from the Sobral (coastal shallow marine to deltaic environment) and Freixial (coastal delta plain to fluvial environment) Formations (Hill, 1988; Manuppella et al., 1999). The Sobral Fm. crops out on the northern sector of this sub-basin. MG 8803 is an unpublished element from Serra da Vila (a locality reported by Lapparent and Zbyszewski, 1957, figure 1), identified herein as a fragment of the distal end of a tibia (Figure 19.11-12). In Ponte de Rol (Torres Vedras), yet unpublished sauropod remains were found (Figure 19.15-16), which include dorsal and caudal vertebrae, and several appendicular elements (scapula, femur, radius, etc.), probably belonging to a single individual (SHN 532). A detailed study of this specimen is in progress, in order to provide a more detailed systematic assessment.
Praia Azul (Torres Vedras) is a locality with several fossil sites. Several sauropod specimens have been collected here, including a single individual comprising dorsal and sacral vertebrae and dorsal ribs (SHN 533), of appreciable size, and dorsal vertebrae with high neural spines and dorsolaterally projecting transverse processes. SHN 533 was extracted in 2014 by the SHN. Other vertebrate groups are also reported from this locality (e.g., Pérez-García, 2015). Several unpublished sauropod specimens are deposited in the palaeontological collections of the SHN, including a compressed cone-chisel-shaped tooth (SHN 549), and some unprepared partial skeletons and isolated bones. Some caudal vertebrae (MG 8804, MG 25254.2-4) were found close to Maceira (Torres Vedras). In particular, MG 8804 (Figure 19.14) was identified as Lusotitan atalaiensis by Lapparent and Zbyszewski (1957) and listed as cf. Lusotitan atalaiensis by Mateus (2005).
 Three of the most important quarries with sauropod dinosaurs in the Turcifal Sub-basin are located near the Cambelas locality, where the Freixial Formation crops out. One is located in Casal da Costa, where sauropod appendicular and axial material were recovered (Figure 20, SHN 531), including dorsal vertebrae, and material from other groups of dinosaurs (theropods and ornithischians), turtles, and crocodyliforms. The dorsal vertebrae show a similar morphology to those of Camarasaurus supremus (Mocho et al., 2013a, 2013b). Some of these specimens were recently recovered from the field and are now under preparation. Several finds were also reported on the Cambelas cliffs, including a site where several elements, probably from the same individual, were collected. This sauropod (SHN (JJS) 178) comprises axial and appendicular elements (Figure 19.17-19), including pelvic girdle and hindlimb remains (Mocho et al., 2014a). It was preliminarily referred to Diplodocidae based on the presence of a markedly expansion of the ischiatic distal end (Mocho et al., 2014a). The pronounced lateral bulge on the femur observed in this specimen is generally considered to be a synapomorphy of Titanosauriformes (e.g., Wilson, 2002) and also occurs in Diplodocus (e.g., Hatcher, 1901). A preliminary evaluation of SHN (JJS) 178 suggests a close relationship with an unpublished diplodocid recovered in Valmitão (SHN (JJS) 177), which shares affinities with the Diplodocinae (Mocho et al., 2014a). Finally, a partial skeleton with axial and appendicular elements (Figure 19.7-8, SHN 529) was found in a distinct fossil site from the Cambelas locality, and includes an anterior caudal neural spine with a delta-shaped distal end, a feature common in camarasaurids (Ikejiri, 2004; Mocho et al., 2014b). Several sauropod finds have been identified in the Freixial Fm. in the section of the Cambelas, Gentias, and Assenta cliffs, including several teeth, axial, and appendicular elements (Figure 19.1-5, 19.9, 19.13). The collected teeth include heart-shaped (Figure 19.1-2; see also Mocho et al., 2012, 2016b) and spatulate teeth (Figure 19.5, see also Mocho et al., 2011), and one compressed cone-chisel-shaped tooth (Figure 19.3-4). In Porto Barril (Mafra), an unpublished possible middle cervical vertebra (Figure 19.6, SHN 528) was found, with a well-developed camellate internal bone tissue similar to the condition present in basal titanosauriforms and mamenchisaurids (following Wedel, 2003).
Three of the most important quarries with sauropod dinosaurs in the Turcifal Sub-basin are located near the Cambelas locality, where the Freixial Formation crops out. One is located in Casal da Costa, where sauropod appendicular and axial material were recovered (Figure 20, SHN 531), including dorsal vertebrae, and material from other groups of dinosaurs (theropods and ornithischians), turtles, and crocodyliforms. The dorsal vertebrae show a similar morphology to those of Camarasaurus supremus (Mocho et al., 2013a, 2013b). Some of these specimens were recently recovered from the field and are now under preparation. Several finds were also reported on the Cambelas cliffs, including a site where several elements, probably from the same individual, were collected. This sauropod (SHN (JJS) 178) comprises axial and appendicular elements (Figure 19.17-19), including pelvic girdle and hindlimb remains (Mocho et al., 2014a). It was preliminarily referred to Diplodocidae based on the presence of a markedly expansion of the ischiatic distal end (Mocho et al., 2014a). The pronounced lateral bulge on the femur observed in this specimen is generally considered to be a synapomorphy of Titanosauriformes (e.g., Wilson, 2002) and also occurs in Diplodocus (e.g., Hatcher, 1901). A preliminary evaluation of SHN (JJS) 178 suggests a close relationship with an unpublished diplodocid recovered in Valmitão (SHN (JJS) 177), which shares affinities with the Diplodocinae (Mocho et al., 2014a). Finally, a partial skeleton with axial and appendicular elements (Figure 19.7-8, SHN 529) was found in a distinct fossil site from the Cambelas locality, and includes an anterior caudal neural spine with a delta-shaped distal end, a feature common in camarasaurids (Ikejiri, 2004; Mocho et al., 2014b). Several sauropod finds have been identified in the Freixial Fm. in the section of the Cambelas, Gentias, and Assenta cliffs, including several teeth, axial, and appendicular elements (Figure 19.1-5, 19.9, 19.13). The collected teeth include heart-shaped (Figure 19.1-2; see also Mocho et al., 2012, 2016b) and spatulate teeth (Figure 19.5, see also Mocho et al., 2011), and one compressed cone-chisel-shaped tooth (Figure 19.3-4). In Porto Barril (Mafra), an unpublished possible middle cervical vertebra (Figure 19.6, SHN 528) was found, with a well-developed camellate internal bone tissue similar to the condition present in basal titanosauriforms and mamenchisaurids (following Wedel, 2003).
In conclusion, the Turcifal Sub-basin presents a relatively continuous Upper Jurassic sequence, with a rich sauropod record. The sauropod fauna identified in the Turcifal Sub-basin is composed of non-neosauropod eusauropods (probably turiasaurs), diplodocids (with probable diplodocine affinities), and macronarians, including camarasaurids and members within Titanosauriformes + Europasaurus.
SAUROPOD PALAEOBIODIVERSITY AND LITHOSTRATIGRAPHIC DISTRIBUTION FOR THE LUSITANIAN BASIN
The Upper Jurassic record of the Lusitanian Basin is rich in dinosaur occurrences and, in particular, in fossil sites with sauropod remains. The Portuguese Late Jurassic sauropods have been the focus of several recent papers, including the description of new material (Yaguë et al., 2006; Mocho et al., 2011; Mannion et al., 2012; Mocho et al., 2012, 2013a, 2013b; Mateus et al., 2014; Mocho et al., 2014a, 2016a, 2016b, 2017), as well as the systematic revision of previously established taxa (Mannion et al., 2012, 2013; Mocho et al., 2014b, 2016c). A geographical and lithostratigraphic context for the sauropod record reported in the Lusitanian Basin is proposed here for the first time. We also report several new finds that highlight the potential of the Portuguese record for the understanding of the palaeobiodiversity and the evolutionary history of the Late Jurassic sauropod fauna from the Iberian Peninsula. The stratigraphic distribution of this fossil record suggests that the major clades (i.e., non-neosauropod eusauropods, diplodocoids, and macronarians) are present throughout the continental and transitional sedimentary sequence dated from the early Kimmeridgian to the late Tithonian. The only exceptions, which might reflect sampling bias, are: i) Camarasauridae (specimens attributed to Macronaria indet. might pertain to Camarasauridae), which were only identified with confidence in the Sobral and Freixial Formations, at the most meridional sub-basins of Arruda and Turcifal; ii) Diplodocinae remains unreported in the Sobral Fm., the most marine-influenced Tithonian unit in these sub-basins; and iii) Brachiosauridae, only present with confidence in the Sobral Fm. accepting Lusotitan as a member of this clade (Mannion et al., 2013; Mocho et al., 2016c).
Turiasauria is a non-neosauropod eusauropod clade, and its presence in the Upper Jurassic of the Lusitanian Basin was first recognised by Royo-Torres et al. (2006). The heart-shaped tooth morphology has been proposed as exclusive of turiasaurs (Royo-Torres et al., 2006, 2009; Mocho et al., 2012; Royo-Torres and Upchurch, 2012; Royo-Torres et al., 2014a). Nevertheless, the wide stratigraphic (Middle Jurassic to Lower Cretaceous) and palaeogeographic (Europe and Africa) range for this shape suggests that this morphology of teeth could be a feature of a more inclusive group or a convergent feature in other sauropod clades (Mocho et al., 2016b). Heart-shaped teeth have a wide stratigraphic distribution throughout the Upper Jurassic levels of the Lusitanian Basin, being present in the Alcobaça, Praia da Amoreira-Porto Novo, Sobral, Bombarral and Freixial Formations. Zby atlanticus, found in Praia da Amoreira-Porto Novo Formation, is the only specimen that can be confidently assigned to Turiasauria (Mateus, 2005; Mateus et al., 2014). Nevertheless, other specimens found in the Portuguese Upper Jurassic record have been preliminarily identified as non-neosauropod eusauropods, and they could represent turiasaurians (non-neosauropod eusauropods outside Turiasauria have not been identified from the Iberian Upper Jurassic record so far). Non-neosauropod eusauropods have a wide stratigraphic range in the Upper Jurassic levels of Portugal (Figure 3). Non-neosauropod eusauropod remains have been recovered from the coastal marine to lagoonal and fluvio-deltaic Alcobaça Fm., the fluvial Praia da Amoreira-Porto Novo Fm. (e.g., ML 368, SHN 530), the marine to deltaic Sobral Fm. (e.g., SHN 533), the fluvial to deltaic Freixial Fm. and the fluvial-lacustrine Bombarral Fm.
Diplodocoidea is a clade recorded from the upper Kimmeridgian up to probably the upper Tithonian of the Portuguese Upper Jurassic record (Figure 3). This clade is represented by one described diplodocine, Dinheirosaurus lourinhanensis, and material considered as belonging to indeterminate diplodocines (Mannion et al., 2012; Mocho et al., 2014a; Tschopp et al., 2015; Mocho et al., 2016a, 2017). Based on the material from Moita dos Ferreiros (ML 418), Mannion et al. (2012) suggested the presence of at least two diplodocids in the Portuguese Upper Jurassic levels. The specimen from Moita dos Ferreiros was subsequently considered as an indeterminate diplodocine by Tschopp et al. (2015). Diplodocines have been recorded from the Praia da Amoreira-Porto Novo, Bombarral, and Freixial Formations. The presence of this clade in the Alcobaça (coastal marine to lagoonal and fluvio-deltaic environments) or Sobral (deltaic environments) Formations is so far unreported. Apatosaurines are apparently absent in the Upper Jurassic of the Iberian Peninsula. Apatosaurinae is considered an endemic group from the Late Jurassic of the North America (Foster and Peterson, 2015).
Macronarians are a diverse group in the Iberian territory, represented by the Portuguese Lourinhasaurus and Lusotitan, and the Spanish Aragosaurus and Galveosaurus (e.g., Barco, 2009; Mannion et al., 2013; Mocho et al., 2014b; Royo-Torres et al., 2014b), and a putative new form from Portugal (SHN 181). Several indeterminate specimens assigned to Macronaria have been found in the Upper Jurassic record of the Lusitanian Basin, present in the Alcobaça, Praia da Amoreira-Porto Novo, Sobral, Freixial, and Bombarral Formations. A stratigraphic range from the lower Kimmeridgian to the probably upper Tithonian is therefore recognised (Figure 3). Camarasaurids are clearly identified in the Sobral Fm. (Lourinhasaurus alenquerensis) and Freixial Fm. (SHN 531), but the systematic affinities of several other specimens needs to be evaluated (e.g., SHN 002, from the Praia da Amoreira-Porto Novo Fm., found in Porto Novo, Torres Vedras; and SHN 529, from the Freixial Fm., found in Cambelas, Torres Vedras), in order to understand the true stratigraphic range of this group. At the moment, Camarasauridae is recorded from the upper Kimmeridgian-basal Tithonian to probably upper Tithonian (Figure 3). If the Porto Novo specimens belong to Camarasauridae, the stratigraphic range of the clade would be upper Kimmeridgian to probably upper Tithonian, and SHN 002 would be the first occurrence of a camarasaurid from the Praia da Amoreira-Porto Novo Fm.
Titanosauriforms are present in the Praia da Amoreira-Porto Novo, Sobral, and Bombarral Formations. Lusotitan atalaiensis is a possible brachiosaurid (Mannion et al., 2013; Mocho et al., 2016c) from the Sobral Fm. Apart from the presence of several specimens with titanosauriform affinities, Lusotitan might be the strongest evidence for the presence of this clade during the Upper Jurassic of the Lusitanian Basin. Several compressed cone-chisel teeth were found in the Upper Jurassic sedimentary sequence of the Lusitanian Basin (Alcobaça, Praia da Amoreira-Porto Novo, Sobral, Freixial, and Bombarral Formations). This tooth morphology was traditionally considered as synapomorphic of Titanosauriformes (e.g., Upchurch et al., 2004; D’Emic, 2012), but this morphology was also found in the possible basal macronarian Europasaurus (Carballido and Sander, 2014), which was also considered to be a member of Titanosauriformes by D’Emic (2012) and Mannion et al. (2013). If we accept the phylogenetic position proposed for Europasaurus by Carballido and Sander (2014), this tooth morphology should be present in a more inclusive group than Titanosauriformes. In this case, the compressed cone-chisel teeth found in the Lusitanian Basin should be referred to Macronaria indet. New discoveries are necessary to improve our knowledge about the Iberian titanosauriforms, especially about brachiosaurids, which is mainly based on incomplete specimens.
Several new sauropod specimens, belonging to different clades, are herein reported. Nevertheless, the collections of the ML and SHN still house many specimens that need to be prepared, catalogued, and studied (see Mateus, 2005; Mocho et al., 2013a, 2013b, 2014a; pers. observ., PM). New relevant specimens have also recently been discovered and collected by the Museu Nacional de História Natural e da Ciência, SHN and ML. Herein, we report some of the most important sauropod finds, including informative partial skeletons, such as the specimens from Praia Vermelha (SHN (JJS) 179, Peniche), Porto das Barcas (ML 351, Lourinhã; see Antunes and Mateus, 2003; Mateus, 2005), Porto Novo (SHN 002, Torres Vedras), Praia da Corva (SHN 530, Torres Vedras), Santa Rita (SHN 534, Torres Vedras), Praia Azul (SHN 533, Torres Vedras), and Cambelas (SHN 529, Torres Vedras). The systematic study of all these finds will significantly improve our knowledge of Portuguese Late Jurassic sauropod fauna. The study of other, so far unpublished, specimens will also be informative, such as those including axial and appendicular elements, in particular the basal macronarians recovered in Casal da Costa (SHN 531, Torres Vedras) and Valmitão (SHN 181, Lourinhã) and the diplodocids from Valmitão (SHN (JJS) 177, related to Diplodocinae), and Cambelas (SHN (JJS) 178, Torres Vedras). These specimens will allow a better understanding of the palaeobiodiversity of Diplodocinae and Camarasauridae in the Upper Jurassic record of the Lusitanian Basin. SHN 181, from Valmitão (Praia da Amoreira-Porto Novo Formation, late Kimmeridgian-basal Tithonian in age) is a specimen that is currently being described, and might represent a new basal macronarian, increasing the palaeobiodiversity of the Portuguese Late Jurassic sauropod fauna, which seems to be comprised of at least six taxa: one turiasaur (Zby), two different diplodocines (Dinheirosaurus and one indeterminate diplodocine); and at least three macronarians: one camarasaurid (Lourinhasaurus), one brachiosaurid (Lusotitan), and one non-camarasaurid basal macronarian (SHN 181).
CONCLUSIONS
The Upper Jurassic sequence of the central sector of the Lusitanian Basin has yielded a rich sauropod dinosaur fossil record, particularly abundant in the Bombarral and Turcifal Sub-basins. A geographic and stratigraphic context for the known sauropod record in the Lusitanian Basin is proposed, and several new specimens are herein reported for the first time. Sauropods are present throughout the continental to transitional deposits of the Lusitanian Basin, recorded in the Alcobaça, Praia da Amoreira-Porto Novo, Sobral, Freixial, and Bombarral Formations. Some areas of the Bombarral Sub-basin and Arruda Sub-basin are still poorly understood, due to scarce and incomplete material. The identified sauropod fauna are mainly comprised of non-neosauropod eusauropods (turiasaurs and indeterminate forms), diplodocids (diplodocines and indeterminate forms), basal macronarians (non-camarasaurids and camarasaurids), and titanosauriforms (some specimens with brachiosaurid affinities). No particular stratigraphic pattern was identified within the Upper Jurassic sequence, and the major clades are present throughout the lower Kimmeridgian to upper Tithonian levels. However, the detailed relationships between the different palaeoenvironments and sauropod fauna will require a systematic analysis of the geological context (formation, facies, environment) of the listed findings, which is not always easy to assure, given the uncertain provenance of some specimens. Such analysis may eventually shed some light on the spatio-temporal distribution of the fauna and its control by stratigraphic and/or palaeoenvironmental factors.
ACKNOWLEDGMENTS
This research was supported by SFRH/BD/68450/2010 PhD scholarship, financed by the "Fundação para a Ciência e Tecnologia" (Portugal); Haaga Family Postdoctoral Fellow; Departamento de Educación, Universidad, Cultura y Deporte del Gobierno de Aragón, the Departamento de Industria e Innovación and the Fondo Social Europe (FOCONTUR, Ref. E62); and the Instituto Aragonés de Fomento, the Spanish Ministerio de Economía y Competitividad (CGL2013-41295-P Project DINOTUR). This research received support from the Synthesys Project (http://synthesys3.myspecies.info/), which is financed by the European Community Research Infrastructure Action under the FP7 (FR-TAF-5072 and DE-TAF-6138). We want to thank for allow accessing specimens to M. Ramalho and R. Silva (MG, LNEG, Portugal), L. Chiappe, and M. Walsh (NHMLAC, USA), the Herrero family (MPG, Spain), L. Póvoas, V.F. dos Santos and B. Ribeiro (MUHNAC, Portugal), M. Cachão and C.M. da Silva (GeoFCUL, Portugal), M.F.C. Pereira (MDT, IST, Portugal), L. Machado (MMPM, Portugal), C. Manso (MMB, Portugal), L. Alcalá (FCPT-Dinópolis, Spain), A. Aberasturi (MUPE), D. Schwarz (HNM, Germany), S. Chapman (NHMUK, UK), and R. Allain (MNHN, France). Comments made by O.W.M. Rauhut, an anonymous reviewer, and the editor helped improve an early version of this paper.We also thank P. Dantas (MUHNAC, Portugal), F. Marcos (UCM, Spain), A. Parámo and A. Serrano (UNED, Spain), G. Ramalheiro, A. Mano and J.J. dos Santos (SHN, Portugal) for help and comments. The study was supported by a protocol between CMTV and SHN.
REFERENCES
Antunes, M.T., Becquart, D., and de Broin, F. 1988. Découverte de “Plesiochelys”, Chélonien marin-littoral, dans le Kimméridgien d’Alcobaça, Portugal. Ciências da Terra, 3:179-195.
Antunes, M.T. and Mateus, O. 2003. Dinosaurs of Portugal. Palevol, 2:77-95.
doi:10.1016/S1631-0683(03)00003-4
Araújo, R., Castanhinha, R., Martins, M.S., Mateus, O., Hendrickx, C., Beckmann, F., Schell, N., and Alves, L.C. 2013. Filling the gaps of dinosaur eggshell phylogeny: Late Jurassic Theropod clutch with embryos from Portugal. Scientific Reports, 3:1924.
doi:10.1038/srep01924
Araújo, R., Castanhinha, R., Mateus, O., and Martins, R. 2012. Late Jurassic theropod embryos from Porto das Barcas, Lourinhã formation, Portugal. Journal of Vertebrate Paleontology, Program and Abstracts, 32:57.
Azerêdo, A.C., Cabral, M.C., Martins, M.J., Loureiro, I.M., and Inês, N. 2010. Estudo estratigráfico dum novo afloramento da Formação de Cabaços (Oxfordiano) na região da Serra do Bouro (Caldas da Rainha). Comunicações Geológicas, 97:5-22.
Balbino, A.C. 2003. Upper Jurassic Hybodontidae (Selachii) from Lourinhã, Portugal. Ciências da Terra, 15:45-52.
Barco, J.L. 2009. Sistemática e implicaciones filogenéticas y paleobiogeográficas del saurópodo Galvesaurus herreroi (Formación Villar del Arzobispo, Galve, España). Unpublished PhD Thesis, Universidad de Zaragoza, Zaragoza, Spain.
Barrett, P.M., Benson, R.B.J., and Upchurch, P. 2010. Dinosaurs of Dorset: Part II, the sauropod dinosaurs (Saurischia, Sauropoda) with additional comments on the theropods. Proceedings of the Dorset Natural History and Archaeological Society, 131:113-126.
Bernardes, C. 1992. A sedimentação durante o Jurássico Superior entre o Cabo Mondego e o Baleal (Bacia Lusitana): Modelos depositionais e arquitectura sequencia. Unpublished PhD Thesis, Departamentode Geociências, Aveiro, Portugal.
Boillot, G., Dupeuble, P.A., and Muselec, P. 1978. Carta Geológica da plataforma continental portuguesa, 1/1000000. Serviços Geológicos de Portugal, Lisboa.
Bonaparte, J.F. and Mateus, O. 1999. A new diplodocid, Dinheirosaurus lourinhanensis gen et sp. nov., from the Late Jurassic beds of Portugal. Revista del Museo Argentino de Ciencias Naturale, 5:13-29.
Bräm, H. 1973. Chelonia from the Upper Jurassic of Guimarota Mine (Portugal). Memórias dos Serviços Geológicos de Portugal, 22:135-141.
Broschinski, A. 2000. The lizards from the Guimarota mine, p. 59-68. In Martin, T. and Krebs, B. (eds.), Guimarota - A Jurassic Ecosystem. Verlag Dr. Friedrich Pfeil, München.
Caldwell, M.W., Nydam, R.L., Palci, A., and Apesteguía, S. 2015. The oldest known snakes from the Middle Jurassic-Lower Cretaceous provide insights on snake evolution. Nature Communications, 6:5996.
doi:10.1038/ncomms6996
Camarate França, J. and Zbyszewski, G. 1963. Notícia Explicativa da Carta Geológica de Portugal, folha 26-B (Alcobaça). Serviços Geológicos de Portugal, Lisboa.
Camarate França, J., Zbyszewski, G., and Almeida, F.M. 1960. Carta geológica de Portugal, na escala 1/50000. Notícia Explicativa da folha 26-C (Peniche). Serviços Geológicos de Portugal, Lisboa.
Carballido, J.L. and Sander, P.M. 2014. Postcranial axial skeleton of Europasaurus holgeri (Dinosauria, Sauropoda) from Upper Jurassic of Germany: implications for sauropod ontogeny and phylogenetic relationships of basal Macronaria. Journal of Systematic Paleontology, 12(3):335-387.
doi: 10.1080/14772019.2013.764935
Casanovas, M.L., Santafé, J.V., and Sanz, J.L. 2001. Losillasaurus giganteus, un nuevo saurópodo del tránsito Jurásico-Cretácico de la cuenca de «Los Serranos» (Valencia, España). Paleontologia i Evolución, 32-33:99-122.
Castanhinha, R., Araújo, R., and Mateus, O. 2009. Dinosaur eggshell and embryo localities in Lourinhã Formation, Late Jurassic, Portugal. Journal of Vertebrate Paleontology, 29(3):76A.
Castanhinha, R. and Mateus, O. 2007. Short review on the marine reptiles of Portugal: ichthyosaurs, plesiosaurs and mosasaurs. Journal of Vertebrate Paleontology, 27:57A.
Cobos, A. and Gascó, F. 2013. New vertebral remains of the stegosaurian dinosaur Dacentrurus from Riodeva (Teruel, Spain). Geogaceta, 53:17-20.
Cobos, A., Royo-Torres, R., Gascó, F., and Alcalá, L. 2011. A new giant turiasaurian specimen from Riodeva (Teruel, Spain), p. 18. In Geer, A. van der and Athanassiou, A. (eds.), Programs and Abstracts of 9th Annual Meeting of the European Association of Vertebrate Palaeontologists. Creta, Greece.
Cobos, A., Royo-Torres, R., Luque, L., Alcalá, L., and Mampel, L. 2010. An Iberian stegosaurs paradise: The Villar del Arzobispo Formation (Tithonian-Berriasian) in Teruel (Spain). Palaeogeography, Palaeoclimatology, Palaeoecology, 293:223-236.
doi:10.1016/j.palaeo.2010.05.024
Dantas, P. 1987. Contribuição ao estudo da macrofauna de vertebrados do Jurássico Superior português. O corte de Paimogo. Unpublished Estágio Científico da Licenciatura em Geologia, Faculdade de Ciências de Lisboa, Lisboa, Portugal.
Dantas, P. 1990. Dinossáurios de Portugal. Gaia, 2:17-26.
Dantas, P., Pérez-Moreno, B.P., Chure, D.J., da Silva, C.M., dos Santos, V.F., Povoas, L., Cachão, M., Sanz, J.L., Pires, C., Bruno, G., Ramalheiro, G., and Galopim de Carvalho, A.M. 1999. O dinossáurio carnívoro Allosaurus fragilis no Jurássico português. Al-Madam, 8:23-28.
Dantas, P., Sanz, J.L., and Galopim de Carvalho, A.M. 1992. Dinossáurio da Praia de Porto Dinheiro (dados preliminares). Gaia, 5:31-35.
Dantas, P., Sanz, J.L., Silva, C.M., Ortega, F., Santos, V.F., and Cachão, M. 1998. Lourinhasaurus n. gen. novo dinossáurio saurópode do Jurássico superior (Kimmeridgiano superior-Titoniano inferior) de Portugal. Actas do V Congresso de Geologia, 84:A-91-A-94.
Dantas, P., Yagüe, P., Hazevoet, C.J., Ortega, F., dos Santos, V., Sanz, J.L., Cachão, M., Galopim de Carvalho, A.M., and Santos, J.J. 2000. Estudo preliminar de um novo Iguanodontia basal do Jurássico Superior Portugués, p. 12-13. In I Congresso Ibérico de Paleontología, Livro de Resumos.
D’Emic, M.D. 2012. The early evolution of titanosauriform sauropod dinosaurs. Zoological Journal of the Linnean Society, 166:624-671. doi:10.1111/j.1096-3642.2012.00853.x
Escaso, F. 2014. Historia evolutiva de los Ornithischia (Dinosauria) del Jurásico Superior de Portugal. Unpublished PhD Thesis. Universidad Autónoma de Madrid, Madrid, Spain.
Escaso, F., Ortega, F., Dantas, P., Malafaia, E., Pimentel, N.L., Pereda-Subelbiola, X., Sanz, J.L., Kullberg, J.C., Kullberg, M.C., and Barriga, F. 2007a. New evidence of shared Dinosaur across Upper Jurassic Proto-North Atlantic: Stegosaurus from Portugal. Naturwissenschaften, 94:367-374. doi:10.1007/s00114-006-0209-8
Escaso, F., Ortega, F., Dantas, P., Malafaia, E., Silva, B., Gasulla, J.M., Mocho, P., Narváez, I., and Sanz, J.L. 2014. A new dryosaurid ornithopod (Dinosauria, Ornithischia) from the Late Jurassic of Portugal. Journal of Vertebrate Paleontology, 34:1102-1112.
doi:10.1080/02724634.2014.849715
Escaso, F., Ortega, F., Dantas, P., Malafaia, E., Silva, B., and Sanz, J.L. 2007b. Elementos postcraneales de Dacentrurus (Dinosauria: Stegosauria) del Jurásico Superior de Moçafaneira (Torres Vedras, Portugal), p. 157-171. In Cambra-Moo, O., Martimez-Pérez, C., Chamero, B., Escaso, F., De Estebar Trivigno, S., and Marugán-Lobón, J. (eds.), Cantera Paleontológica. Diputación Provincial de Cuenca, Cuenca, Spain.
Escaso, F., Ortega, F., Dantas, P., Malafaia, E., Silva, B., and Sanz, J.L. 2008. Estudio preliminar del material del estegosáurido de Vale Pombas (Portugal): nueva evidencia de Stegosaurus en el Jurásico Superior del suroeste europeo, p. 107-108. In Libro de resúmenes XXIV Jornadas de la Sociedad Española de Paleontología, Colunga, Spain.
Escaso, F., Silva, B., Ortega, F., Dantas, P., Malafaia, E., Gasulla, J.M., and Sanz, J.L. 2010a. Análisis preliminar de un nuevo ejemplar de camptosáurido del Jurásico Superior de Portugal, p. 116-117. In Santos, A., Mayoral, E., Meléndez, G., Silva, C.M. da, and Cachão, M. (eds.), III Congresso Ibérico de Paleontologia/XXVI Jornadas de la SEP, Publicaciones del Seminario de Paleontología de Zaragoza, Lisboa, Portugal.
Escaso, F., Silva, B., Ortega, F., Malafaia, E., and Sanz, J.L. 2010b. A Portuguese specimen of Camptosaurus aphanoecetes (Ornithopoda: Camptosauridae) increases the dinosaurian similarity among the Upper Jurassic Alcobaça and Morrison Formations, p. 86. In 70th Meeting of the Society of Vertebrate Paleontology. Program and Abstracts. Pittsburgh, UK.
Foster, J. and Peterson, J.E. 2015. First report of Apatosaurus (Diplodocidae: Apatosaurinae) from the Cleveland-Lloyd Quarry in the Upper Jurassic Morrison Formation of Utah: Abundance, distribution, paleoecology, and taphonomy of an endemic North American sauropod clade. Palaeoworld, vol 25, pages 431-443.
doi: 10.1016/j.palwor.2015.11.006
Fürsich, F.T. 1981. Salinity-controlled benthic associations from the Upper Jurassic of Portugal. Lethaia, 14:203-223.
doi:10.1111/j.1502-3931.1981.tb01690.x
Galton, P.M. 1980. European Jurassic ornithopod dinosaurs of the families Hypsilophodontidae and Camptosauridae. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 160:73-95.
Galton, P.M. 1991. Postcranial remains of stegosaurian dinosaur Dacentrurus from Upper Jurassic of France and Portugal. Geologica et Palaeontologica, 25:299-327.
Gassner, T. 2000. The turtles from the Guimarota mine, p. 55-58. In Martin, T. and Krebs, B. (eds.), Guimarota - A Jurassic Ecosystem. Verlag Dr. Friedrich Pfeil, München.
Gilmore, C.W. 1925. A nearly complete articulated skeleton of Camarasaurus, a saurischian dinosaur from the Dinosaur National Monument, Utah. Memoirs of the Carnegie Museum. Proceedings of the US National Museum, 81:1-21.
Guéry, F. 1984. Évolution sédimentaire et dynamique du bassin marginal ouest-portugais au Jurassique (Province d’Estremadure, secteur de Caldas da Rainha, Montejunto). PhD Thesis, Université Claude Bernard, Lyon, France.
Hahn, G. 1971. The dentition of the Paulchoffatiidae (Multituberculata, Upper Jurassic). Memórias dos Serviços Geológicos de Portugal, 17:7-39.
Hahn, G. and Hahn, R. 2000. Multituberculates from the Guimarota mine, p. 97-108. In Martin, T. and Krebs, B. (eds.), Guimarota - a Jurassic Ecosystem. Verlag Dr. Friedrich Pfeil, München.
Hatcher, J.B. 1901. Diplodocus (Marsh): its osteology, taxonomy and probable habits, with a restoration of the skeleton. Memoirs of the Carnegie Museum, 1:1-63.
Hendrickx, C. and Mateus, O. 2012. Ontogenetical changes in the quadrate of basal tetanurans. ¡Fundamental!, 20:101-104.
Hendrickx, C. and Mateus, O.V. 2014. Torvosaurus gurneyi n. sp., the largest terrestrial predator from Europe, and a proposed terminology of the maxilla anatomy in nonavian Theropods. PLoS ONE, 9: e88905.
doi:10.1371/journal.pone.0088905
Hill, G. 1988. The sedimentology and lithostratigraphy of the Upper Jurassic Lourinhã Formation, Lusitanian Basin, Portugal. Unpublished Phd Thesis, Open University, Milton Keynes, UK.
Hill, G. 1989. Distal alluvial fan sediments from the Upper Jurassic of Portugal: controls on their cylclicity and channel formation. Journal Geologic Society London, 146:539-555.
doi:10.1144/gsjgs.146.3.0539
Ikejiri, T. 2004. Anatomy of Camarasaurus lentus (Dinosauria: Sauropoda) from the Morrison Formation (Late Jurassic), Thermopolis, central Wyoming, with determination and interpretation of ontogenetic, sexual dimorphic, and individual variation in the genus. Unpublished Master Thesis, Fort Hays State University, Kansas, USA.
Krebs, B. 2000. The henkelotheriids from the Guimarota mine, p. 121-142. In Martin, T. and Krebs, B. (eds.), Guimarota - A Jurassic Ecosystem. Verlag Dr. Friedrich Pfeil, München.
Krebs, B. and Schwarz, D. 2000. The crocodiles from the Guimarota mine, p. 69-74. In Martin, T. and Krebs, B. (eds.), Guimarota - A Jurassic Ecosystem. Verlag Dr. Friedrich Pfeil, München.
Kriwet, J. 2000. The fish fauna from the Guimarota mine, p. 41-50. In Martin, T. and Krebs, B. (eds.), Guimarota - A Jurassic Ecosystem. Verlag Dr. Friedrich Pfeil, München.
Krusat, G. 1980. Haldanodon exspectatus Kühne & Krusat 1972 (Mammalia, Docodonta). Memórias dos Serviços Geológicos de Portugal, 27:1-79.
Kullberg, J.C. 2000. Evolução tectónica mesozóica da Bacia Lusitaniana. Unpublished PhD Thesis, Faculdade de Ciências e Tecnologia, Universidade Nova de Lisboa, Lisboa, Portugal.
Kullberg, J.C., Rocha, R.B., Soares, A.F., Rey, J., Terrinha, P., Azerêdo, A.C., Callapez, P., Duarte, L.V., Kullberg, M.C., Martins, L., Miranda, J.R., Alves, C., Mata, J., Madeira, J., Mateus, O., Moreira, M., and Nogueira, C.R. 2013. A Bacia Lusitaniana: Estratigrafia, Paleogeografia e Tectónica, p. 989-1141. In Dias, R., Araújo, A., Terrinha, P., and Kullberg, J.C. (eds.), Geologia de Portugal no contexto da Ibéria. Escolar Editora, Lisboa.
Kullberg, J.C., Rocha, R.B., Soares, A.F., Rey, J., Terrinha, P., Callapez, P., and Martins, L. 2006. A Bacia Lusitaniana: Estratigrafia, Paleogeografia e Tectónica, p. 317-368. In Dias, R., Araújo, A., Terrinha, P., and Kullberg, J.C. (eds.), Geologia de Portugal no contexto da Ibérica. Universidade de Évora, Évora.
Lapparent, A.F. and Zbyszewski, G. 1957. Les dinosauriens du Portugal. Memórias dos Serviços Geológicos de Portugal, 2:1-63.
Leinfelder, R.R. 1993. A sequence stratigraphic approach to the Upper Jurassic mixed carbonate-siliciclastic succession of the central Lusitanian Basin, Portugal. Profil, 5:119-140.
Leinfelder, R.R. and Wilson, R.C.L. 1989. Seismic and sedimentologic features of Oxfordian-Kimmeridgian syn-rift sediments on the eastern margin of the Lusitanian Basin. International Journal of Earth Sciences, Geologische Rundschau, 78:81-104.
doi:10.1007/BF01988355
Lull, R.S.1919. The sauropod dinosaur Barosaurus Marsh: redescription of the type specimens in the Peabody Museum, Yale University. Memoirs of the Connecticut Academy of Arts and Sciences, 6:1-42.
Maidment, S.C.R., Brassey, C., and Barrett, P.M. 2015. The postcranial skeleton of an exceptionally complete individual of the plated dinosaur Stegosaurus stenops (Dinosauria: Thyreophora) from the Upper Jurassic Morrison Formation of Wyoming, U.S.A. PLoS ONE, 10:e0138352. doi:10.1371/journal.pone.0138352
Maidment, S.C.R., Norman, D.B., Barrett, P.M., and Upchurch, P. 2008. Systematics and phylogeny of Stegosauria (Dinosauria: Ornithischia). Journal of Systematic Palaeontology, 6:367-407.
doi:10.1017/S1477201908002459
Malafaia, E., Dantas, P., Ortega, F., and Escaso, F. 2007. Nuevos restos de Allosaurus fragilis (Theropoda: Carnosauria) del yacimiento de Andrés (Jurásico Superior; Centro-Oeste de Portugal), p. 255-272. In Cambra-Moo, O., Martimez-Pérez, C., Chamero, B., Escaso, F., De Estebar Trivigno, S., and Marugán-Lobón, J. (eds.), Cantera Paleontológica. Diputación Provincial de Cuenca, Cuenca.
Malafaia, E., Mocho, P., Escaso, F., and Ortega, F. 2016. A juvenile allosauroid theropod (Dinosauria, Saurischia) from the Upper Jurassic of Portugal. Historical Biology, vol. 29, p. 654-676.
doi:10.1080/08912963.2016.1231183
Malafaia, E., Mocho, P., Escaso, F., and Ortega, F. 2017. New data on the anatomy of Torvosaurus and other remains of megalosauroid (Dinosauria, Theropoda) from the Upper Jurassic of Portugal. Journal of Iberian Geology, vol. 43 p. 33–59.
doi:10.1080/08912963.2016.1231183
Malafaia, E., Ortega, F., Escaso, F., Dantas, P., and Gasulla, J.M. 2009a. Allosaurus fragilis from the Portuguese Upper Jurassic. Journal of Vertebrate Paleontology, 29 (Supplement to Number 3):140-141A.
Malafaia, E., Ortega, F., Escaso, F., Dantas, P., Pimentel, N.L., Gasulla, J.M., Ribeiro, B., Barriga, F., and Sanz, J.L. 2010a. Vertebrate fauna at the Allosaurus fossil-site of Andrés (Upper Jurassic), Pombal, Portugal. Journal of Iberian Geology, 36:193-204.
doi:10.5209/rev_JIGE.2010.v36.n2.7
Malafaia, E., Ortega, F., Escaso, F., Dantas, P., and Silva, B. 2010b. Nova evidência de Ceratosaurus (Theropoda, Ceratosauria) from the Upper Jurassic of Lusitanian Basin (Portugal), p. 157-158. In Santos, A., Mayoral, E., Meléndez, G., Silva, C.M. da and Cachão, M. (eds.), III Congresso Ibérico de Paleontologia/XXVI Jornadas de la SEP, Publicaciones del Seminario de Paleontología de Zaragoza, 9.
Malafaia, E., Ortega, F., Escaso, F., and Silva, B. 2015. New evidence of Ceratosaurus (Dinosauria: Theropoda) from the Late Jurassic of the Lusitanian Basin, Portugal. Historical Biology: An International Journal of Paleobiology, 27:938-946.
doi:10.1080/08912963.2014.915820
Malafaia, E., Ortega, F., Escaso, F., Silva, B., Ramalheiro, G., Dantas, P., Moniz, C., and Barriga, F. 2009b. Análisis preliminar de un nuevo ejemplar de Allosaurus del Grupo Lourinhã (Jurásico Superior de Torres Vedras, Portugal), p. 243-251. In Colectivo Arqueológico y Paleontológico de Salas (ed.), Actas de las IV Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno. Salas de los Infantes, Burgos.
Malafaia, E., Ortega, F., Silva, B., and Escaso, F. 2008. Fragmento de un maxilar de terópodo de Praia da Corva (Jurásico Superior. Torres Vedras, Portugal), p. 273-279. In Esteve J. and Meléndez, G. (eds.), Palaeontologica Nova, SEPAZ (8).
Mannion, P.D. and Calvo, J.O. 2011. Anatomy of the basal titanosaur (Dinosauria, Sauropoda) Andesaurus delgadoi from the mid-Cretaceous (Albian-early Cenomanian) Río Limay Formation, Neuquén Province, Argentina: implications for titanosaur systematics. Zoological Journal of the Linnean Society, 163:155-181.
doi:10.1111/j.1096-3642.2011.00699.x
Mannion, P., Upchurch, P., Barnes, R.N., and Mateus, O. 2013. Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary history of basal titanosauriformes. Zoological Journal of the Linnean Society, 168:98-206.
doi:10.1111/zoj.12029
Mannion, P., Upchurch, P., Mateus, O., Barnes, R.N., and Jones, M.E.H. 2012. New information on the anatomy and systematic position of Dinheirosaurus lourinhanensis (Sauropoda: Diplodocoidea) from the Late Jurassic of Portugal, with a review of European diplodocoids. Journal of Systematic Palaeontology, 10:521-551.
doi:10.1080/14772019.2011.595432
Manuppella, G., Antunes, M.T., Costa Almeida, C.A., Azerêdo, A.C., Barbosa, B., Cardoso, J.L., Crispim, J.A., Duarte, L.V., Henriques, M.H., Martins, L.T., Ramalho, M.M., Santos, V.F., and Terrinha, P. 2000. Notícia Explicativa da Carta Geológica de Portugal, folha 27-A (Vila Nova de Ourém). Departamento de Geologia do Instituto Geolégico e Minerio, Lisboa.
Manuppella, G., Antunes, M.T., Pais, J., Ramalho, M.M., and Rey, J. 1999. Notícia explicativa da Folha 30-A (Lourinhã). Departamento de Geologia do Instituto Geolégico e Minerio, Lisboa.
Martin, T. 2000. The dryolestids and the primitive ‘peramurid’ from the Guimarota mine, p. 109-120. In Martin, T.and Krebs, B. (eds.), Guimarota. A Jurassic ecosystem. Verlag Dr. Friedrich Pfeil, München.
Martin, T. 2005. Postcranial anatomy of Haldanodon exspectatus (Mammalia, Docodonta) from the Late Jurassic (Kimmeridgian) of Portugal and its bearing for mammalian evolution. Zoological Journal of the Linnean Society, 145:219-248.
doi:10.1111/j.1096-3642.2005.00187.x
Martin, T. and Nowotny, M. 2000. The docodont Haldanodon from the Guimarota mine, p. 91-96. In Martin, T. and Krebs, B. (eds.), Guimarota. A Jurassic ecosystem. Verlag Dr. Friedrich Pfeil, München.
Martinius, A. and Gowland, S. 2011. Tide-influenced fluvial bedforms and tidal bore deposits (Late Jurassic Lourinhã Formation, Lusitanian Basin, western Portugal). Sedimentology, 58:285-324.
doi:10.1111/j.1365-3091.2010.01185.x
Mateus, O. 1998. Lourinhanosaurus antunesi, a new Upper Jurassic allosauroid (Dinosauria: Theropoda) from Lourinhã, Portugal. Memórias da Academia das Ciências de Lisboa, Classe Ciências, 37:111-124.
Mateus, O. 2005. Dinossauros do Jurássico Superior de Portugal, com destaque para os saurísquios. Unpublished PhD Thesis. Universidade Nova de Lisboa, Lisboa, Portugal.
Mateus, O. 2006. Late Jurassic dinosaurs from the Morrison Formation, the Lourinhã and Alcobaça Formations (Portugal), and the Tendaguru Beds (Tanzania): a comparison. New Mexico Museum of Natural History and Science, 36:223-231.
Mateus, O. 2009. The sauropod Turiasaurus riodevensis in the Late Jurassic of Portugal. Journal of Vertebrate Paleontology, 29:144A.
Mateus, O. and Antunes, M.T. 2000a. Ceratosaurus sp. (Dinosauria: Theropoda) in the Late Jurassic of Portugal. In Abstract volume of the 31st International Geological Congress, Rio de Janeiro, Brazil.
Mateus, O. and Antunes, M.T. 2000b. Torvosaurus sp. (Dinosauria: Theropoda) in the Late Jurassic of Portugal, p. 115-117. In Livro de Resumos do I Congresso Ibérico de Paleontologia.
Mateus, O. and Antunes, M.T. 2001. Draconyx loureiroi, a new Camptosauridae (Dinosauria, Ornithopoda) from the Late Jurassic of Lourinhã. Annales de Paleontologie, 87:61-73.
doi:10.1016/S0753-3969(01)88003-4
Mateus, O., Dinis, J., and Cunha, P.P. 2013. Upper Jurassic to Lowermost Cretaceous of the Lusitanian Basin, Portugal - landscapes where dinosaurs walked. Ciências da Terra, número especial VIII, 34:618-634.
Mateus, O., Maidment, S.C.R., and Christiansen, N.A. 2009. A new long-necked ‘sauropod-mimic’ stegosaur and the evolution of the plated dinosaurs. Proceedings of the Royal Society of London Series B, 276:1815-1821.
doi:10.1098/rspb.2008.1909
Mateus, O., Mannion, P.D., and Upchurch, P. 2014. Zby atlanticus, a new turiasaurian sauropod (Dinosauria, Eusauropoda) from the Late Jurassic of Portugal. Journal of Vertebrate Paleontology, 34:618-634.
doi:10.1080/02724634.2013.822875
Mateus, I., Mateus, H., Antunes, M.T., Mateus, O., Taquet, P., Ribeiro, V., and Manuppella, G. 1997. Couveé, oeufs et embryons d'un Dinosaure Théropode du Jurassique de Lourinhã (Portugal). Comptes Rendus de l'Académie des Sciences Paris; Sciences de la terre et des planets, 325:71-78.
Mateus, O., Walen, A., and Antunes, M.T. 2006. The large theropod fauna of the Lourinhã Formation (Portugal) and its similarity to the Morrison Formation, with a description of a new species of Allosaurus. New Mexico Museum of Natural History and Science, 36:223-231.
McIntosh, J.S. 1990a. Sauropoda, p. 345-401. In Weishampel, D.B., Dodson, P., and Osmólska, H. (eds.), The Dinosauria. University of California Press, Berkeley, CA.
McIntosh, J.S. 1990b. Species determination in sauropod dinosaurs with tentative suggestions for their classification, p. 53-69. In Carpenter, K. and Currie, P.J. (eds.), Dinosaur systematics: approaches and perspectives. Cambridge University Press, Cambridge.
McIntosh, J.S. 2005. The genus Barosaurus Marsh (Sauropoda, Diplodocidae), p. 38-77. In Tidwell, V. and Carpenter, K. (eds.), Thunder-Lizards: the sauropodomorph dinosaurs. Indiana University Press, Bloomington.
McIntosh, J.S., Miles, C.A., Cloward, K.C., and Parker, J.R. 1996b. A new nearly complete skeleton of Camarasaurus. Bulletin Gunma Museum of Natural History, 1:1-87.
McIntosh, J.S., Miller, W.E., Stadtman, K.L., and Gillette, D.D. 1996a. The osteology of Camarasaurus lewisi (Jensen, 1988). Brigham Young University Geology Studies, 41:73-115.
Mocho, P., Ortega, F., and Royo-Torres, R. 2012. Morphological variation of Turiasauria-like teeth and their stratigraphic distribution in Portuguese Upper Jurassic. ¡Fundamental!, 20:161-163.
Mocho, P., Royo-Torres, R., Malafaia, E., Escaso, F., Narváez, I., and Ortega, F. 2017. New data on Late Jurassic sauropods of central and northern sectors of the Bombarral Sub-basin (Lusitanian Basin, Portugal). Historical Biology: An International Journal of Paleobiology, 29:151-169. doi:10.1080/08912963.2015.1137912
Mocho, P., Royo-Torres, R., Malafaia, E., Escaso, F., and Ortega, F. 2014a. A preliminary evaluation of Diplodocidae record from the Upper Jurassic of Lusitanian Basin (W, Portugal), p. 85-88. In Pereira, I., Amaral, F., and Vinhas, A. (eds.), IV CJIG, LEG 2014, Livro de Actas. Pólo de Estremoz de UÉvora, Évora.
Mocho, P., Royo-Torres, R., Malafaia, E., Escaso, F., and Ortega F. 2016a. Systematic review of Late Jurassic sauropods from the Museu Geológico collections (Lisboa, Portugal). Journal of Iberian Geology, 42:227-250.
Mocho, P., Royo-Torres, R., Malafaia, E., Escaso, F., Silva, B., and Ortega, F. 2016b. Turiasauria-like teeth from the Upper Jurassic of the Lusitanian Basin, Portugal. Historical Biology: An International Journal of Paleobiology, 28:861-880.
doi:10.1080/08912963.2015.1049948
Mocho, P., Royo-Torres, R., and Ortega, F. 2013a. New insights for basal macronarians of Portuguese Upper Jurassic, p. 171-174. In Moreira, N., Pereira, I., Couto, F., and Silva, H. (eds.), III Congresso Jovens Investigadores em Geociências, LEG 2013 & 6th Meeting of Post-Graduation in Physics and Earth Sciences of UÉ, 04-08 outubro 2013, Livro de Actas. Pólo de Estremoz da UÉvora, Évora.
Mocho, P., Royo-Torres, R., and Ortega, F. 2014b. Phylogenetic reassessment of Lourinhasaurus alenquerensis, a basal Macronaria (Sauropoda) from the Upper Jurassic of Portugal. Zoological Journal of the Linnean Society, 170:875-916.
doi:10.1111/zoj.12113
Mocho, P., Royo-Torres, R., and Ortega, F. 2016c. New data of the Portuguese brachiosaurid Lusotitan atalaiensis (Sobral Formation, Upper Jurassic). Historical Biology: An International Journal of Paleobiology, vol. 29 p. 789-817.
doi:10.1080/08912963.2016.1247447
Mocho, P., Royo-Torres, R., Ortega, F., and Silva, B. 2011. "Euhelopodidae" (Sauropoda) teeth from the Upper Jurassic of Portugal, p. 160. In 71st SVP Symposium of Vertebrate Paleontology (SVP), Las Vegas, USA 2nd-5th November. Program and Abstract.
Mocho, P., Royo-Torres, R., Ortega, F., and Silva, B. 2013b. Macronarian record from the Upper Jurassic of Portugal, p. 178. In Maxwell, E., Miller-Camp, J., and Anemone, R. (eds.), 73st SVP Symposium of Vertebrate Paleontology (SVP), Los Angeles, USA 30th Octobre-2nd November. Program and Abstract.
Oliveira, J.T., Pereira, H., Ramalho, M., and Antunes, M.T. 1992. Carta Geológica de Portugal, na escala 1:500000. Serviços Geológicos de Portugal, Lisboa.
Ortega F., Dantas, P., Escaso, F., Gasulla, J.M., Malafaia, E., and Ribeiro, B. 2006. Primera cita de reptiles esfenodontos en el Jurásico Superior de la Península Ibérica, p. 152-153. In Fernández-Martínez, E. (eds.), Libro de resúmenes XXII Jornadas de la Sociedad Española de Paleontología.
Ortega, F., Escaso, F., Gasulla, J.M., Dantas, P., and Sanz, J.L. 2006. Dinosaurios de la Península Ibérica. Estudios Geológicos, 62:219-240.
Ortega, F., Escaso, F., Malafaia, E., Mocho, P., and Pérez-García, A. 2013. Paleobiogeografia dos répteis peri-Atlânticos do Jurássico Superior português: resposta a padrões de vicariância ou dispersão, p. 17-20. In Moreira, N., Pereira, I., Couto, F., and Silva, H. (eds.), Livro de Actas III Congresso Jovens Investigadores em Geociéncias LEG 2013 & 6th Meeting of Post-Graduatíon in Physics and Earth Sciences of University of Évora. Universidade de Évora, Évora.
Ortega, F., Malafaia, E., Escaso, F., Pérez-García, A., and Dantas, P. 2009. Faunas de répteis do Jurássico superior de Portugal. Paleolusitana, 1:43-56.
Ortega, F., Royo-Torres, R., Gascó, F., Escaso, F., and Sanz, J.L. 2010. New evidences of the sauropod Turiasaurus from the Portuguese Upper Jurassic, p. 62. In Abstract Volume 8th Annual Meeting of the European Association of Vertebrate Palaeontologists.
Osborn, H.F. 1899. A skeleton of Diplodocus. Memoirs of the American Museum of Natural History, 1:191-214.
Osborn, H.F. and Mook, C.C. 1921. Camarasaurus, Amphicoelias and other sauropods of Cope. Memoirs of the American Museum of Natural History, new series, 3:247-387.
Ostrom, J.H. and McIntosh, J.S. 1966. Marsh’s dinosaurs. The collection from Como Bluff. Yale University Press, New Haven.
Ouyang, H. and Ye, Y. 2002. The First Mamenchisaurian Skeleton with Complete Skull, Mamenchisaurus youngi. Sichuan Publishing House of Science and Technology, Chengdu. [Chinese].
Pena dos Reis, R.P.B., Dinis, J.L., Cunha, P.M.R., and Trincão, P. 1996. Upper Jurassic sedimentary infill and tectonics of the Lusitanian Basin (western Portugal), p. 377-386. In Riccardi, A.C. (eds.), Advances in Jurassic Research, GeoResearch Forum, Vols 1-2. Zurich.
Pena dos Reis, R., Pimentel, N.L., and Garcia, A.J.V. 2011. The Lusitanian Basin, stratigraphic analysis and geodynamic evolution. Boletim de Geociencias Petrobras, 19:23-52.
Pena dos Reis, R.P.B., Proença Cunha, C.P., Dinis, J.L., and Trincão, P.R. 2000. Geologic evolution of the Lusitanian Basin (Portugal) during the late Jurassic. GeoResearch Forum, 6:345-356.
Pereda-Suberbiola, X., Dantas, P., Galton, P.M., and Sanz, J.L. 2005. Autopodium of the holotype of Dracopelta zbyszewskii (Dinosauria, Ankylosauria) and its type horizon and locality (Upper Jurassic: Tithonian, western Portugal). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 235:175-196.
Pérez-García, A. 2015. Revision of the British record of Tropidemys (Testudines, Plesiochelyidae) and recognition of its presence in the Late Jurassic of Portugal. Journal of Iberian Geology, 41:11-20.
doi:10.5209/rev_JIGE.2015.v41.n1.48651
Pérez-García, A. and Ortega, F. 2011. Selenemys lusitanica, gen. et sp. nov., a new pleurosternid turtle (Testudines: Paracryptodira) from the Upper Jurassic of Portugal. Journal of Vertebrate Paleontology, 31:60-69.
doi:10.1080/02724634.2011.540054
Pérez-García, A. and Ortega, F. 2013. A new species of the turtle Hylaeochelys (Eucryptodira) outside its known geographic and stratigraphic ranges of distribution. Comptes Rendus Palevol, 13:183-188.
doi:10.1016/j.crpv.2013.10.009
Pérez-García, A., Ortega, F., Murelaga, X., and Dantas, P. 2008. Plesiochelys sp. (Testudines; Eucryptodira) de la Fm. Freixial (Jurásico Superior) en Ulsa (Torres Vedras, Portugal). Publicaciones del Seminario de Paleontología de Zaragoza, 8:331-344.
Pérez-Moreno, B.P., Chure, D.J., Pires, C., Silva, C.M., Santos, V.F., Dantas, P., Póvoas, L., Cachão, M., Sanz, J.L., and Galopim de Carvalho, A.M. 1999. On the presence of Allosaurus fragilis (Theropoda, Carnosauria) in the Upper Jurassic of Portugal: first evidence of an intercontinental dinosaur species. Journal of the Geological Society, 156:449-452.
doi:10.1144/gsjgs.156.3.0449
Rasmussen, E.S., Lomholt, S., Andersen, C., and Vejbaek, O.V. 1998. Aspects of the structural evolution of the Lusitanian Basin in Portugal and the shelf and slope area offshore Portugal. Tectonophysics, 300:199-225.
doi:10.1016/S0040-1951(98)00241-8
Rauhut, O.W.M. 2000. The dinosaur fauna of the Guimarota mine, p. 75-82. In Martin, T. and Krebs, B. (eds.), Guimarota - A Jurassic Ecosystem. Verlag Dr. Friedrich Pfeil, München.
Rauhut, O.W.M. 2001. Herbivorous dinosaurs from the Late Jurassic (Kimmeridgian) of Guimarota, Portugal. Proceedings of the Geologists' Association, 112:275-283.
Rauhut, O.W.M. 2003. A tyrannosauroid dinosaur from the Upper Jurassic of Portugal. Palaeontology, 46:903-910.
Rauhut, O.W.M. and Fechner, R. 2005. Early development of the facial region in a non-avian theropod dinosaur. Proceedings of the Royal Society B, 272:1179-1183.
Rauhut, O.W.M., Remes, K., Fechner, R., Cladera, G., and Puerta, P. 2005. Discovery of a short-necked sauropod dinosaur from the Late Jurassic period of Patagonia. Nature, 435:670-672.
doi:10.1038/nature03623
Remes, K. 2006. Revision of the Tendaguru sauropod dinosaur Tornieria Africana (Fraas) and its relevance for sauropod paleobiogeography. Journal of Vertebrate Paleontology, 26:651-669.
doi: 10.1671/0272-4634(2006)26[651:ROTTSD]2.0.CO;2
Rey, J. 1999. Lower Cretaceous Depositional Sequences in the Cascais Area, p. 1-57. In European Palaeontological Association Workshop, Field trip A.
Ribeiro, A., Antunes, M.T., Ferreira, M.P., Rocha, R.B., Soares, A.E., Zbyszewski, G., Moitinho de Almeida F., Carvalho, D.E., and Monteiro, J.H. 1979. Introduction á la géologie générale du Portugal. Serviços Geológicos de Portugal, Lisboa.
Ribeiro, A., Silva, J.B., Cabral, J., Dias, R., Fonseca, P., Kullberg, M.C., Terrinha, P., and Kullberg, J.C. 1996. Tectonics of the Lusitanian Basin. Final Report, Proj. MILUPOBAS, Contract nº JOU-CT94-0348, ICTE/GG/GeoFCUL, Lisboa.
Ribeiro, V. and Mateus, O. 2012. Chronology of the Late Jurassic dinosaur faunas, and other reptilian faunas, from Portugal. Journal of Vertebrate Paleontology, Program and Abstracts, 32:161-161.
Rocha, R.B. (coord.), Marques, B.L., Kullberg, J.C., Caetano, P.C., Lopes, C., Soares, A.F., Duarte, L.V., Marques, J.F., and Gomes, C.R. 1996. The 1 st and 2 nd rifting phases of the Lusitanian Basin: stratigraphy, sequence analysis and sedimentary evolution. Final Report C. E. C. Proj. MILUPOBAS, Lisboa.
Rocha, R.B. and Soares, A.F. 1984. Algumas reflexões sobre a sedimentação jurássica na orla meso-cenozóica ocidental de Portugal. Memórias e Notícias, 97:133-142.
Royo-Torres, R., Cobos, A., and Alcalá, L. 2006. A giant european dinosaur and a new sauropod clade. Science, 314:1925-1927.
doi:10.1126/science.1132885
Royo-Torres, R., Cobos, A., Gascó, F., Mocho, P., Ortega, F., and Alcalá, L. 2014a. Geographic and Stratigraphic distribution of the sauropod Turiasaurus and Turiasauria clade, p. 74-75. In Kirkland, J., Foster, J., Hunt-Foster, R., Liggett, G.A., and Trujillo, K. (eds.), Mid-Mesozoic: The Age of Dinosaurs in Transition, April 30 - May 5, 2014, Fruita, Colorado & Green River, Utah. Utah.
Royo-Torres, R., Cobos, A., Luque, L., Aberasturi, A., Espílez, E., Fierro, I., González, A., Mampel, L., and Alcalá, L. 2009. High European sauropod dinosaur diversity during Jurassic-Cretaceous transition in Riodeva (Teruel, Spain). Palaeontology, 52:1009-1027.
doi:10.1111/j.1475-4983.2009.00898.x
Royo-Torres, R. and Upchurch, P. 2012. The cranial anatomy of the sauropod Turiasaurus riodevensis and implications for its phylogenetic relationships. Journal of Systematic Palaeontology, 10:553-583.
doi:10.1080/14772019.2011.598577
Royo-Torres, R., Upchurch, P., Mannion, P.D., Mas, R., Cobos, A., Gascó, F., Alcalá, L., and Sanz, J.L. 2014b. The anatomy, phylogenetic relationships and stratigraphic position of the Tithonian-Berriasian Spanish sauropod dinosaur Aragosaurus ischiaticus. Zoological Journal of the Linnean Society, 71:623-655.
doi:10.1111/zoj.12144
Russo, J., Mateus, O., Balbino, A., and Marzola, M. 2014b. Crocodylomorph eggs and eggshells from the Lourinhã Fm. (Upper Jurassic), Portugal. Comunicações Geológicas, 101(Especial I):563-566.
Russo, J., Mateus, O., Marzola, M., and Balbino, A. 2014a. Eggs and eggshells of Crocodilomorpha from the Late Jurassic of Portugal, p. 218. In Maxwell, E. and Miller-Camp, J. (eds.), 74th SVP Symposium of Vertebrate Paleontology (SVP), Berlin, USA 5th- 8th November. Program and Abstract.
Russo, J., Mateus, O., Marzola, M., and Balbino, A. 2017. Two new ootaxa from the late Jurassic: The oldest record of crocodylomorph eggs, from the Lourinhã Formation, Portugal. PLoS ONE, 12(3):e0171919.
doi:10.1371/journal.pone.0171919
Salgado, L., Coria, R.A., and Calvo, J.O. 1997. Evolution of titanosaurid sauropods. I: phylogenetic analysis based on the postcranial evidence. Ameghiniana, 34:3-32.
Sander, M.P., Mateus, O., Laven, T., and Knötschke, N. 2006. Bone histology indicates insular dwarfism in a new Late Jurassic sauropod dinosaur. Nature, 441:739-741.
Sauvage, H.E. 1897-98. Vértebrés fossiles du Portugal. Contribution à l’étude des poissons et des reptiles du Jurassique et du Crétacé. Mémoires de la Direction des Travaux Géologiques du Portugal.
Schneider, S., Fürsich, F.T., and Werner, W. 2009. Sr-isotope of the Upper Jurassic of central Portugal (Lusitanian Basin) based on oyster shells. International Journal of Earth Sciences, Geologische Rundschau, 98:1949-1970.
doi:10.1007/s00531-008-0359-3
Schudack, M. 2000. Ostracodes and charophytes from the Guimarota beds, p. 33-36. In Martin, T. and Krebs, B. (eds.), Guimarota. A Jurassic ecosystem. Verlag Dr. Friedrich Pfeil, München.
Schwarz, D. and Fechner, R. 2004. Lusitanisuchus, a new generic name for Lisboasaurus mitracostatus (Crocodylomorpha: Mesoeucrocodylia), with a description of new remains from the Upper Jurassic (Kimmeridgian) and Lower Cretaceous (Berriasian) of Portugal. Canadian Journal of Earth Sciences, 41:1259-1271.
doi:10.1139/e04-059
Schwarz, D. and Salisbury, S.W. 2005. A new species of Theriosuchus (Atoposauridae, Crocodylomorpha) from the Late Jurassic (Kimmeridgian) of Guimarota, Portugal. Géobios, 38:779-802.
doi:10.1016/j.geobios.2004.04.005
Seiffert, J. 1973. Upper Jurassic lizards from central Portugal. Memórias dos Serviços Geológicos de Portugal, 22:7-85.
Sereno, P.C., Beck, A.L., Dutheil, D.B., Larsson, H.C.E., Lyon, G.H., Moussa, B., Sadleir, R.W., Sidor, C.A., Varricchio, D.J., Wilson, G.P., and Wilson, J.A. 1999. Cretaceous sauropods from the Sahara and the uneven rate of skeletal evolution among dinosaurs. Science, 282:1342-1347. doi:10.1126/science.286.5443.1342
Taylor, A.M., Gowland, S., Leary, S., Keogh, K.J., and Martinius, A.W. 2014. Stratigraphical correlation of the Lourinhã Formation in the Consolação Sub-basin (Lusitanian Basin), Portugal. Geological Journal, 49:143-162.
doi:10.1002/gj.2505
Teixeira, C., Camarate França, J., Zbyszewski, G., Veiga Ferreira, O., Matos, M., Oliveira, J., Rodrigues, L., Rodrigues, A., and Manuppella, G. 1966. Notícia Explicativa da Carta Geológica de Portugal, folha 23-C (Leiria). Serviços Geológicos de Portugal, Lisboa.
Tennant, J.P. and Mannion, P.D. 2014. Revision of the Late Jurassic crocodyliform Alligatorellus, and evidence for allopatric speciation driving high diversity in western European atoposaurids. PeerJ, 2:e599.
doi: 10.7717/peerj.599
Thulborn, R.A. 1973. Teeth of ornithischian dinosaurs from the Upper Jurassic of Portugal. Memórias dos Serviços Geológicos de Portugal, 22:89-134.
Tschopp, E. and Mateus, O. 2012. A sternal plate of a large-sized sauropod dinosaur from the Late Jurassic of Portugal. ¡Fundamental!, 20:263-266.
Tschopp, E. and Mateus, O. 2013. The skull and neck of a new flagellicaudatan sauropod from the Morrison Formation and its implication for the evolution and ontogeny of diplodocid dinosaurs. Journal of Systematic Palaeontology, 11:853-888.
doi:10.1080/14772019.2012.746589
Tschopp, E., Mateus, O., and Benson, R. 2015. A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda). Peer J, 3:e857.
doi: 10.7717/peerj.857
Upchurch, P., Barrett, P.M., and Dodson, P. 2004. Sauropoda, p. 259-322. In Weishampel, D.B., Dodson, P., and Osmólska, H. (eds.), The Dinosauria II. University Of California Press, Berkeley.
Wedel, M.J. 2003. Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs. Paleobiology, 29:243-255.
doi:10.1666/0094-8373(2003)029<0243:VPASAT>2.0.CO;2
Wedel, M.J. and Taylor, M.P. 2013. Caudal pneumaticity and pneumatic hiatuses in the sauropod dinosaurs Giraffatitan and Apatosaurus. PLoS ONE, 8(10):e78213.
doi:10.1371/journal.pone.0078213
Weigert, A. 1995. Isolierte Zähne von cf. Archaeopteryx sp. aus dem oberen Jura der Kohlengrube Guimarota (Portugal). Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, Mh 1995(9):562-576.
Whitlock, J.A. 2011. A phylogenetic analysis of Diplodocoidea (Saurischia: Sauropoda). Zoological Journal of the Linnean Society, 161:872-915. doi:10.1111/j.1096-3642.2010.00665.x
Wiechmann, M.F. 2000. The albanerpetontids from the Guimarota mine, p. 51-54. In Martin, T. and Krebs, B. (eds.), Guimarota - A Jurassic Ecosystem. Verlag Dr. Friedrich Pfeil, München.
Wiechmann, M.F. and Gloy, U. 2000. Pterosaurs and urvogels from the Guimarota Mine, p. 83-86. In Martin, T. and Krebs, B. (eds.), Guimarota - A Jurassic Ecosystem. Verlag Dr. Friedrich Pfeil, München.
Wilson, J.A. 1999. A nomenclature for vertebral laminae in sauropods and other saurischian dinosaurs. Journal of Vertebrate Paleontology, 19:639-653.
Wilson, J.A. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society, 136:217-276.
doi:10.1046/j.1096-3642.2002.00029.x
Wilson, J.A. 2006. Anatomical nomenclature of fossil vertebrates: standardized terms or lingua franca? Journal of Vertebrate Paleontology, 26:511-518.
Wilson, J.A. 2012. New vertebral laminae and patterns of serial variation in vertebral laminae of sauropod dinosaurs. Contributions from the Museum of Paleontology, 32:91-110.
Wilson, J.A., D'Emic, M.D., Ikejiri, T., Moacdieh, E.M., and Withlock, J.A. 2011. A nomenclature for vertebral fossae in sauropods and other saurischian dinosaurs. PLoS One, 6(2):e17114.
doi: 10.1371/journal.pone.0017114.
Wilson, J.A. and Sereno, P.C. 1998. Early evolution and higher level phylogeny of sauropod dinosaurs. Journal of Vertebrate Paleontology, 18(Suppl. 2):1-68.
doi:10.2307/3889325
Wilson, J.A. and Upchurch, P. 2009. Redescription and reassessment of the phylogenetic affinities of Euhelopus zdanskyi (Dinosauria: Sauropoda) from the Early Cretaceous of China. Journal of Systematic Palaeontology, 7:199-239.
doi:10.1017/S1477201908002691
Wilson, R.C.L. 1988. Mesozoic development of the Lusitanian basin, Portugal. Revista de la Sociedad Geológica de España, 1(3-4):393-407.
Yagüe, P., Dantas, P., Ortega, F., Cachão, M., Santos, F.A.M., Gonçalves, R., and Lopes, S. 2006. New sauropod material from the Upper Jurassic of Areia Branca (Lourinha, Portugal). Neues Jahrbuch ftir Geologie und Paliiontologie Abhandlungen, 240:313-342.
Zbyszewski, G. 1946. Les ossements d' Omosaurus découverts près de Baleal (Peniche). Comunicações dos Serviços Geológicos de Portugal, 28:135-144.
Zbyszewski, G. and Ferreira, O.V. 1978. Carta geológica de Portugal na escala de 1/50.000: notícia explicativa da folha 23A-Pombal. Serviços Geológicos de Portugal, Lisboa.
Zbyszewski, G., Ferreira, O.V., Manuppella, G., and Torres de Assunção, C. 1966. Carta geológica de Portugal na escala de 1/50.000: notícia explicativa da folha 30-B, Bombarral: micropaleontologia: rochas eruptivas. Serviços Geológicos de Portugal, Lisboa.
Zbyszewski, G., Manuppella, G., Ferreira, O.V., Mouterde, R., Perrot, C.R., and Torres de Assunção, C. 1974. Carta geológica de Portugal na escala de 1/50.000: notícia explicativa da folha 27-A, Vila Nova de Ourém: rochas eruptivas. Serviços Geológicos de Portugal, Lisboa.
Zbyszewski, G. and Moitinho de Almeida, F. 1960. Carta geológica de Portugal na escala de 1:50.000: notícia explicativa da folha 26-D Caldas da Rainha. Serviços Geológicos de Portugal, Lisboa.
Zbyszewski, G., Moitinho de Almeida, F., and Torres de Assunção, C. 1955. Carta geológica de Portugal na escala de 1/50.000: notícia explicativa da folha 30-C, Torres Vedras: rochas eruptivas. Serviços Geológicos de Portugal, Lisboa.
Zbyszewski, G. and Torre de Assunção, C. 1965. Carta geológica de Portugal na escala de 1:50.000: Notícia explicativa da carta geológica de Portugal, 1:50000, folha 30-D, Alenquer. Serviços Geológicos de Portugal, Lisboa.
Zinke, J. 1998. Small theropod teeth from the Upper Jurassic coal mine of Guimarota (Portugal). Paläontologische Zeitschrift, 72:179-189. doi:10.1007/BF02987825
Zinke, J. and Rauhut, O.W.M. 1994. Small theropods (Dinosauria, Saurischia) from the Upper Jurassic and Lower Cretaceous of the Iberian Peninsula. Berliner Geowissenschaftliche Abhandlungen Reihe E, 13:163-177.
