

 Shell anatomy and intraspecific variability of the Spanish Lutetian podocnemidid turtle Neochelys zamorensis
Shell anatomy and intraspecific variability of the Spanish Lutetian podocnemidid turtle Neochelys zamorensis
Article number: 27.2.a28
https://doi.org/10.26879/1388
Copyright Palaeontological Association, June 2024
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 7 March 2024. Acceptance: 18 May 2024.
ABSTRACT
Two Eocene podocnemidid turtles are recognized in the Duero Basin (northwestern Spain), both exclusive to the middle Eocene record of that region: the Bartonian Neochelys salmanticensis and the Lutetian Neochelys zamorensis. Information on the shell anatomy of the former, identified at several sites in the Salamanca Province, is much greater than that available for any other representative of this exclusively European Eocene genus. However, only one specimen indisputably attributable to Neochelys zamorensis has so far been described and figured. It corresponds to a shell of a subadult individual, from the Sanzoles fossil site, in the Zamora Province. Some well-preserved partial shells of several individuals from its type locality are analyzed here, including adult specimens, so that knowledge about this hitherto poorly known taxon is markedly increased. In addition, numerous unpublished shells and plates from the same geological formation, and found in various fossil sites in the Zamora Province, are studied. They are also recognized as attributable to Neochelys zamorensis. A selection of nearly 200 specimens allows us to study in detail the shell anatomy of this taxon. Given the great availability of remains, and their generally good state of preservation, the shell intraspecific variability of the species is analyzed here (considering aspects such as the ontogeny, the dimorphism, and the individual variability), with a degree of precision much greater than that so far recognized for any other European pleurodiran representative.
Adán Pérez-García. Grupo de Biología Evolutiva, Dpto. de Física Matemática y de Fluidos, Facultad de Ciencias, UNED, Avda. Esparta s/n, 28232, Las Rozas, Madrid, Spain. (Corresponding author) a.perez.garcia@ccia.uned.es
Andrea Guerrero. Grupo de Biología Evolutiva, Dpto. de Física Matemática y de Fluidos, Facultad de Ciencias, UNED, Avda. Esparta s/n, 28232, Las Rozas, Madrid, Spain. guerbach@gmail.com
Santiago Martín de Jesús. Colección de Vertebrados Fósiles de la Cuenca del Duero (Sala de las Tortugas), Departamento de Geología, Facultad de Ciencias, Universidad de Salamanca, Salamanca, Spain. saladelastortugas@usal.es
Francisco Ortega. Grupo de Biología Evolutiva, Dpto. de Física Matemática y de Fluidos, Facultad de Ciencias, UNED, Avda. Esparta s/n, 28232, Las Rozas, Madrid, Spain. fortega@ccia.uned.es
Keywords: Paleogene; middle Eocene; Spain; Zamora Province; Pleurodira; Podocnemididae
Final citation: Pérez-García, Adán, Guerrero, Andrea, Martín de Jesús, Santiago, and Ortega, Francisco. 2024. Shell anatomy and intraspecific variability of the Spanish Lutetian podocnemidid turtle Neochelys zamorensis. Palaeontologia Electronica, 27(2):a28.
https://doi.org/10.26879/1388
palaeo-electronica.org/content/2024/5220-shell-of-neochelys-zamorensis
Copyright: June 2024 Palaeontological Association.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
 The Paleogene record of turtles from the Iberian Peninsula is mostly restricted to that of the Eocene (Pérez-García, 2017). The Eocene turtle fauna of the Duero Basin (northwestern Spain; Figure 1) is very relevant within the European context, considering both its diversity (members of Testudinidae, Carettochelyidae, Trionychidae, and Pelomedusidae being represented there) and, especially, the abundance of remains, several taxa exclusive to this region having been defined (see Ortega et al., 2022 and references therein). Among the turtle lineages identified there, the best represented are the podocnemidids, being recorded from the middle to the late Eocene (from the Lutetian, MP13 or MP14; to the Priabonian, MP19). The first representative of Podocnemididae described there was ‘Stereogenys’ salmanticensis Jiménez Fuentes, 1968 (now recognized as attributable to the genus Neochelys; see Pérez-García et al., 2023, and references therein). This species lived during the Bartonian (middle Eocene), having been exclusively recognized in the adjacent municipal terms of Cabrerizos and Aldealengua, in the Salamanca Province. Although very limited information on the shell of this species was available until recently (only nine partial specimens had been documented), the recent study of about 140 unpublished shells and shell remains has allowed it to be currently considered as the best characterized species of the abundant and diverse European Eocene genus Neochelys (Pérez-García et al., 2023). After the establishment of the species ‘Stereogenys’ salmanticensis, several forms from the Duero Basin were defined (see Pérez-García, 2017; and references therein). However, two of them, ‘Podocnemis entodermica’ Jiménez Fuentes, 1970, and ‘Podocnemis carbajosai’ Jiménez Fuentes, 1971, from the Bartonian of the Salamanca Province, are currently recognized as junior synonyms of Neochelys salmanticensis (Pérez-García et al., 2023). There is another species also defined in the Duero Basin, in older levels (Lutetian, MP13 or MP14) of the Sanzoles Municipality, in the Zamora Province (Figure 1). It is Neochelys zamorensis Jiménez Fuentes, 1993. Very limited information on this smaller podocnemidid is currently available. By contrast, very abundant material, hitherto unpublished, could potentially be attributable to it.
The Paleogene record of turtles from the Iberian Peninsula is mostly restricted to that of the Eocene (Pérez-García, 2017). The Eocene turtle fauna of the Duero Basin (northwestern Spain; Figure 1) is very relevant within the European context, considering both its diversity (members of Testudinidae, Carettochelyidae, Trionychidae, and Pelomedusidae being represented there) and, especially, the abundance of remains, several taxa exclusive to this region having been defined (see Ortega et al., 2022 and references therein). Among the turtle lineages identified there, the best represented are the podocnemidids, being recorded from the middle to the late Eocene (from the Lutetian, MP13 or MP14; to the Priabonian, MP19). The first representative of Podocnemididae described there was ‘Stereogenys’ salmanticensis Jiménez Fuentes, 1968 (now recognized as attributable to the genus Neochelys; see Pérez-García et al., 2023, and references therein). This species lived during the Bartonian (middle Eocene), having been exclusively recognized in the adjacent municipal terms of Cabrerizos and Aldealengua, in the Salamanca Province. Although very limited information on the shell of this species was available until recently (only nine partial specimens had been documented), the recent study of about 140 unpublished shells and shell remains has allowed it to be currently considered as the best characterized species of the abundant and diverse European Eocene genus Neochelys (Pérez-García et al., 2023). After the establishment of the species ‘Stereogenys’ salmanticensis, several forms from the Duero Basin were defined (see Pérez-García, 2017; and references therein). However, two of them, ‘Podocnemis entodermica’ Jiménez Fuentes, 1970, and ‘Podocnemis carbajosai’ Jiménez Fuentes, 1971, from the Bartonian of the Salamanca Province, are currently recognized as junior synonyms of Neochelys salmanticensis (Pérez-García et al., 2023). There is another species also defined in the Duero Basin, in older levels (Lutetian, MP13 or MP14) of the Sanzoles Municipality, in the Zamora Province (Figure 1). It is Neochelys zamorensis Jiménez Fuentes, 1993. Very limited information on this smaller podocnemidid is currently available. By contrast, very abundant material, hitherto unpublished, could potentially be attributable to it.
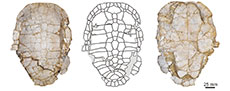
 The presence of Paleogene vertebrates in the Duero Basin was reported for the first time a century and a half ago, based on remains found in the Sanzoles Municipality, corresponding to crocodile teeth (Vilanova, 1873). Over more than a century, remains of various groups of vertebrates were found there. Among them, the discovery made in 1977 of several shell remains (including some relatively complete ones; see Figure 2) in the vicinity of the town of Sanzoles, in the here called El Escobar fossil site (see Figure 1; and the point 11 in the map B of Jiménez Fuentes, 1977), stands out. Jiménez Fuentes (1977) reported this discovery, attributing the remains found there to a small pelomedusid, without providing more information. Subsequently, Jiménez Fuentes (1982) indicated, without justification, that the Sanzoles specimens, which are older than those known for the Salamanca Province (Lutetian instead of Bartonian, see Ortega et al., 2022 and references therein), corresponded to a new, undefined species. Thus, the El Escobar specimens were referred to ‘Neochelys’ zamorensis (Arribas Moreno et al., 1984; Jiménez Fuentes, 1986) or Neochelys zamorensis (Jiménez Fuentes et al., 1985) in several works published in the following years, this putative new species not having been formally described (lacking figuration, description, diagnosis, or explicit allusion to specific specimens, so that it fails to conform to Article 13 of the International Code of Zoological Nomenclature) and, therefore, the species being recognized as a nomen nudum. Since 1987, and for 15 years, the attribution Neochelys aff. arenarum was used for some of the pelomedusids of the Zamora Province (e.g., Mulas and Alonso Gavilán, 1987; Mulas Alonso et al., 1988; Jiménez Fuentes et al., 1991, 1992), including those from El Escobar as well as others from synchronous localities (e.g., outcrops on the western margin of the Infiernos Hill, in the Monfarracinos Municipality, where the Cubillos 1 site is located; as well as those from the Valdegallina 1 and Valdegallina 2 sites, in the Valdegallina locality of the Zamora Municipality; Figure 1). Although Jiménez Fuentes et al. (1992) indicated that there was a clear geographical separation between the French species Neochelys arenarum and the Spanish form Neochelys aff. arenarum, features that differentiate both taxa were not justified. Thus, it was only indicated that the relative small size of the Spanish taxon, and the presence of a straight anterior plastral margin on it, were shared with Neochelys arenarum, a species from which it differed in the ratio between the entoplastral length and the distance of this plate to the hyo-hypoplastral suture, without having been explicitly indicated what the difference for those features are; in addition to pointing out that there were differences in other not specified parameters (Mulas Alonso et al., 1988). No specimen attributable to this Spanish turtle was described in any of those publications. Although the El Escobar specimens were not figured, only photographs corresponding to a plastron, as well as to a skull and a lower jaw from the Valdegallina locality, were published (see figures 1 and 2 in Jiménez Fuentes et al., 1991; the plastron corresponds to STUS 10609, which is here analyzed in detail). Jiménez Fuentes (1992) referred to the El Escobar taxon as ‘Neochelys zamorensis’, alluding to the absence of descriptions and figuration about it; and he published, for the first time, a photograph in dorsal view and another in ventral view of the most complete shell from that locality (see figure 4 in Jiménez Fuentes, 1992; corresponding to that in Figure 2A-B). Although Jiménez Fuentes (1993) indicated that the species Neochelys zamorensis was valid from the aforementioned work published in 1992, because a specimen was figured there for the first time, no holotype was established, nor was a description made, or a diagnosis proposed for the taxon. Therefore, we propose here that the authorship of the taxon should be that of the work of Jiménez Fuentes (1993) since it was in that publication that the act of establishment of the species occurs, following all the requirements indicated in the International Code of Zoological Nomenclature so that the name of the taxon was valid.
The presence of Paleogene vertebrates in the Duero Basin was reported for the first time a century and a half ago, based on remains found in the Sanzoles Municipality, corresponding to crocodile teeth (Vilanova, 1873). Over more than a century, remains of various groups of vertebrates were found there. Among them, the discovery made in 1977 of several shell remains (including some relatively complete ones; see Figure 2) in the vicinity of the town of Sanzoles, in the here called El Escobar fossil site (see Figure 1; and the point 11 in the map B of Jiménez Fuentes, 1977), stands out. Jiménez Fuentes (1977) reported this discovery, attributing the remains found there to a small pelomedusid, without providing more information. Subsequently, Jiménez Fuentes (1982) indicated, without justification, that the Sanzoles specimens, which are older than those known for the Salamanca Province (Lutetian instead of Bartonian, see Ortega et al., 2022 and references therein), corresponded to a new, undefined species. Thus, the El Escobar specimens were referred to ‘Neochelys’ zamorensis (Arribas Moreno et al., 1984; Jiménez Fuentes, 1986) or Neochelys zamorensis (Jiménez Fuentes et al., 1985) in several works published in the following years, this putative new species not having been formally described (lacking figuration, description, diagnosis, or explicit allusion to specific specimens, so that it fails to conform to Article 13 of the International Code of Zoological Nomenclature) and, therefore, the species being recognized as a nomen nudum. Since 1987, and for 15 years, the attribution Neochelys aff. arenarum was used for some of the pelomedusids of the Zamora Province (e.g., Mulas and Alonso Gavilán, 1987; Mulas Alonso et al., 1988; Jiménez Fuentes et al., 1991, 1992), including those from El Escobar as well as others from synchronous localities (e.g., outcrops on the western margin of the Infiernos Hill, in the Monfarracinos Municipality, where the Cubillos 1 site is located; as well as those from the Valdegallina 1 and Valdegallina 2 sites, in the Valdegallina locality of the Zamora Municipality; Figure 1). Although Jiménez Fuentes et al. (1992) indicated that there was a clear geographical separation between the French species Neochelys arenarum and the Spanish form Neochelys aff. arenarum, features that differentiate both taxa were not justified. Thus, it was only indicated that the relative small size of the Spanish taxon, and the presence of a straight anterior plastral margin on it, were shared with Neochelys arenarum, a species from which it differed in the ratio between the entoplastral length and the distance of this plate to the hyo-hypoplastral suture, without having been explicitly indicated what the difference for those features are; in addition to pointing out that there were differences in other not specified parameters (Mulas Alonso et al., 1988). No specimen attributable to this Spanish turtle was described in any of those publications. Although the El Escobar specimens were not figured, only photographs corresponding to a plastron, as well as to a skull and a lower jaw from the Valdegallina locality, were published (see figures 1 and 2 in Jiménez Fuentes et al., 1991; the plastron corresponds to STUS 10609, which is here analyzed in detail). Jiménez Fuentes (1992) referred to the El Escobar taxon as ‘Neochelys zamorensis’, alluding to the absence of descriptions and figuration about it; and he published, for the first time, a photograph in dorsal view and another in ventral view of the most complete shell from that locality (see figure 4 in Jiménez Fuentes, 1992; corresponding to that in Figure 2A-B). Although Jiménez Fuentes (1993) indicated that the species Neochelys zamorensis was valid from the aforementioned work published in 1992, because a specimen was figured there for the first time, no holotype was established, nor was a description made, or a diagnosis proposed for the taxon. Therefore, we propose here that the authorship of the taxon should be that of the work of Jiménez Fuentes (1993) since it was in that publication that the act of establishment of the species occurs, following all the requirements indicated in the International Code of Zoological Nomenclature so that the name of the taxon was valid.
The work of Jiménez Fuentes (1993) was the first publication in which drawings of the shell of Neochelys zamorensis were presented (see figure 2 in that paper), indicating the margins of both the scutes and the plates. Furthermore, that shell, recognized as the holotype of the taxon, was described for the first time. A differential diagnosis in relation to Neochelys arenarum was proposed (without indication on which specimens and from which localities in the Zamora Province it was based, apart from the holotype). In this sense, Jiménez Fuentes (1993) indicated, without justifying it, that the species could also be present in other sites in that province, in addition to the type locality. These include some fossil sites currently recognized as approximately synchronous with the type locality (the Cubillos 1, Tardemayas 1, Valdegallina 1, and Valdegallina 2 sites, whose material will be analyzed in detail in this work), but also others that are currently considered as stratigraphically younger (Madridanos and Santa Clara de Avedillo), and in which the podocnemidids represented seem to correspond to different taxa (see Ortega et al., 2022 and references therein). Therefore, that comparison between the Spanish and the French species must be taken with caution. Jiménez Fuentes (1993) considered that there were differences in the ratio between the entoplastral length and the distance between that plate and the hyo-hypoplastral suture (as in Mulas Alonso et al., 1988), but characterizing for the first time the state present in each of these taxa. In addition, he established differences based on five other features, relating to the ornamental pattern, the ratio between the entoplastral length and the distance of that plate to the pectoral-abdominal sulcus, the number of neural plates, and the general morphology of the carapace.
The species Neochelys zamorensis was cited in several subsequently published papers, generally being included in faunal lists or when the diversity of turtles from the Zamora Province, the Cuenca del Duero, or the Iberian Peninsula were considered (e.g., Jiménez Fuentes and Gil Tudanca, 1993; Pérez-García, 2017); but also when the diversity of the genus Neochelys was discussed (e.g., Pérez-García and de Lapparent de Broin, 2013, 2015). Given the lack of information about the species, Jiménez Fuentes and Gil Tudanca (1993) indicated, without justification and in contradiction to the idea previously proposed (Jiménez Fuentes, 1993), that Neochelys zamorensis could not be the taxon recorded in fossil sites as those of the Valdegallina locality or the Monfarracinos Municipality (where sites as Cubillos 1 and Tardemayas 1 are located). Thus, the only indisputably specimen attributable to Neochelys zamorensis described and figured is its holotype, and a valid diagnosis considering the current knowledge on the genus Neochelys is not available for the species. Given the presence of other well-preserved relatively complete and partial shells in its type locality, but also that of numerous unpublished shell remains coming from the aforementioned fossiliferous areas of the same Zamora Province and geological formation (comprising a quantity of remains much greater than that hitherto documented for any other species of Neochelys), the objective of this work is the detailed analysis of these shell remains. Thus, the availability of such abundant material will allow us to analyze the shell anatomy and intraspecific variability of Neochelys zamorensis based on the material from its type locality, but also to evaluate its possible presence in other localities, significantly improving the knowledge on the shell anatomy of the species, as well as on the interspecific variability within the most relevant genus of Podocnemididae for the European record.
Institutional abbreviations. MGM, Museo Geominero, Instituto Geológico y Minero de España, Madrid, Spain; MNHN.RA, Reptiles and Amphibians Collection of the Muséum national d’Histoire naturelle, Paris, France; STUS, Collection of Fossil Vertebrates of the Duero Basin (Sala de las Tortugas) of the Universidad de Salamanca, Salamanca, Spain.
Anatomical abbreviations. Ab, abdominal; An, anal; c, costal; Ex, extragular; en, entoplastron; ep, epiplastron; Fe, femoral; Gu, gular; hp, hypoplastron; Hu, humeral; hy, hyoplastron; M, marginal; ms, mesoplastron; n, neural; nu, nuchal; p, peripheral; Pc, pectoral; Pl, pleural; py, pygal; spy, suprapygal; V, vertebral; xi, xiphiplastron.
MATERIALS AND METHODS
The holotype and so far only figured material of Neochelys zamorensis (STUS 12121) from the Lutetian (middle Eocene, Entrala Formation) of El Escobar fossil site (Sanzoles Municipality, Zamora Province, Spain; Figure 1) is analyzed in detail (Figure 2A-B). Furthermore, all other podocnemidid specimens that were found in the same fossiliferous assemblage (Figure 2C-J), which remained unpublished until now, are studied here. Given that most Neochelys species are exclusively known through the shell, the analysis of this anatomical region is essential to compare Neochelys zamorensis with all other so far defined representatives of the genus. The first-hand detailed study of shell remains of all these species has been carried out by us, in addition to considering all the information about their shells available in the literature. For the shell of Neochelys arenarum see figures 10 to 12 in Broin (1977); for Neochelys laurenti see figures 5 to 10 in Pérez-García and Lapparent de Broin (2015); for Neochelys liriae see figures 2 and 3 in Pérez-García and Lapparent de Broin (2013); for Neochelys eocaenica see figures 19 to 22 in Broin (1977); for Neochelys capellinii see figures 5 to 8 and 55 in Broin (1977); for Neochelys franzeni see figures 1 to 3, and 6 in Cadena (2015); and for Neochelys salmanticensis see figures 2 to 8 in Pérez-García et al. (2023).
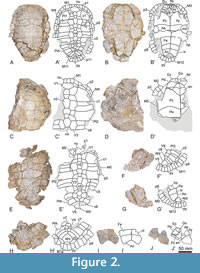
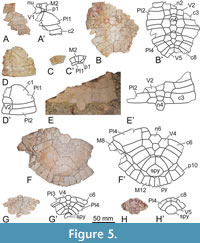
Very abundant (more than 1,300) but until now unpublished remains from the same province and formation as the type locality of Neochelys zamorensis, deposited in two Spanish scientific collections (the Museo Geominero of the Instituto Geológico y Minero de España, in Madrid; and, specially, the “Sala de las Tortugas” or Collection of Fossil Vertebrates of the Duero Basin of the Universidad de Salamanca, in Salamanca) have been analyzed by us. They come from several sites (the Valdegallina 1, Valdegallina 2, and Valdegallina 3 fossil sites, in the Valdegallina locality of the Zamora Municipality; the Cubillos 1 and Tardemayas 1 fossil sites, in the Infiernos Hill of the Monfarracinos Municipality; and the Teso de la Mora fossil site, in the Torres del Carrizal Municipality; Figure 1). A selection of the 184 most relevant specimens, both for the anatomical analysis of the shell and for the study of its variability, has been made [i.e., all available complete and partial shells (8 specimens), carapaces (16), and plastra (29); and all non-significatively deformed isolated and complete or almost complete plates informative for the analysis of both the anatomy and variability, corresponding to the nuchals (25), epiplastra (26), entoplastra (33), and xiphiplastra (47); Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9, Figure 10, Figure 11, Figure 12, Figure 13]. The comparisons with other Neochelys species made here are mostly based on the first-hand study of individuals attributable to most of them, but also considering all the information previously provided in other papers.
Quantitative analyses were conducted, employing the two-dimensional geometric morphometric method (GMM), to scrutinize shell regions exhibiting heightened morphological variability (i.e., nuchal, epiplastron, entoplastron, and xiphiplastron), due to both intraindividual variability (i.e., the nuchal, epiplastral, and entoplastral areas, see Table S1, Table S2, Table S3 in Appendix 1) and sexual dimorphism (i.e., the xiphiplastral region, Figure 14, Figure 15, Figure 16, and also Table S4, Table S5 in Appendix 1). The two-dimensional GMM has been performed following to the methodology outlined by Zelditch et al. (2012) (for more details see Guerrero and Pérez-García, 2021a, 2021b). Landmark selection has been made based on their capacity to faithfully depict geometric forms (see Table S1, Table S2, Table S3, Table S4, Table S5 in Appendix 1; Larson, 2002). Furthermore, semilandmarks were incorporated to delineate the contours and capture the shape along the curves of the shell elements (Gunz and Mitteroecker, 2013). Scale bars were employed for the scaling of each digitized specimen. The digitization process was executed using the tpsDig v.2.16 software (Rohlf, 2010, 2015), with all data collection steps performed by the same operator to eliminate interobserver variation in the data (Fruciano, 2016). All sets of landmarks underwent scaling, translation, and rotation through Generalized Procrustes Analysis (GPA; Rohlf and Slice, 1990; Zelditch et al., 2012). This procedural step was conducted in R v. 4.0.1 (R Core Team, 2020), using the gpagen function in geomorph (Adams et al., 2021). Semilandmarks were slid using the bending energy method (Gunz and Mitteroecker, 2013), chosen for its compatibility with the inherent characteristics of the study sample. To evaluate shape variations, the main maximal axes were described through Principal Component Analysis (PCA). In this instance, a PCA obtained from both Procrustes coordinates and shape data corrected for allometry has not been included, given that the allometric factor does not influence significatively in the shape variation of the study sample (see Figure 14-Figure 15 and Figure S1, Figure S2, Figure S3, Figure S4, Figure S5, Figure S6 in Appendix 1). The determination of the number of principal components (PCs) for assessment in each Principal Component Analysis (PCA) was conducted using the broken-stick method (Jackson, 1993). The PCA was executed utilizing the gm.prcomp function within the geomorph package. Allometry was statistically examined through the logarithm (base 10) of centroid size (log[CS]) values, treated as the independent variable, against the Procrustes shape coordinates, considered as the dependent variable. The allometric regression analysis was carried out using the procD.lm function from the geomorph R package. Multivariate normality for each analyzed element was tested, specifically employing the Shapiro-Wilk test in R v. 4.0.1 (R Core Team, 2020), with the mvnormtest package (Jarek, 2012).
An in-depth analysis of the xiphiplastral region was conducted to characterize the potential sexual dimorphism in Neochelys zamorensis. For the geometric morphometric analyses of this region, we initially performed quantitative analyses of the complete anal notch (i.e., considering both the left and the right sides). We observed that they yielded basically the same results as those where only a margin was considered. Taking this into account, we decided to use the analyses that involves fewer variables (i.e., the second ones) to ensure that the statistical power is not compromised by the available sample size. The evaluation of the potential sexual dimorphism in the xiphiplastral region also involved the integration of two additional statistical approaches: a Procrustes ANOVA and a box plot illustrating Centroid Size variation. Specifically, ANOVA encompassed an evaluation of sexual shape and size dimorphism based on initially defined morphotypes derived from the qualitative analysis. The analysis, which employed morphotypes as a factor with 10,000 permutations, provided insights into the sexual variations. Likewise, a cross-validation test, specifically the leave-one-out cross-validation, was executed to gauge the precision of morphotype classification. In addition, a box plot representing Centroid Size variation for each morphotype visually depicted the distribution within each group. A t-test was performed to confirm that the means were significantly different. Both analyses were executed using R v. 4.0.1 (R Core Team, 2020). Furthermore, a quantitative approximation of the sexual variation on the xiphiplastral region was conducted on various extant podocnemidid species (i.e., an adult male and an adult female of the following taxa: Erymnochelys madagascariensis, Podocnemis expansa, Podocnemis lewyana, Podocnemis unifilis, and Podocnemis vogli, see Figure S5-Figure S6 in Appendix 1). The aim of that analysis was to ascertain whether the sexual morphotypes observed in Neochelys zamorensis align with the shape of the xiphiplastron of a specific sex within the extant podocnemidids. The analyzed specimens of extant podocnemidids are housed in the Comparative Anatomy and Zoology of Reptiles and Amphibians collections, at the Muséum national d’Histoire Naturelle, in Paris (France). For the extant representatives, the same GMM protocol described above was followed.
SYSTEMATIC PALEONTOLOGY
Suborder PLEURODIRA Cope, 1864
Hyperfamily PELOMEDUSOIDES Cope, 1868
Superfamily PODOCNEMIDOIDEA Cope, 1868
Family PODOCNEMIDIDAE Cope, 1868
Subfamily ERYMNOCHELYINAE Broin, 1988
Genus NEOCHELYS Bergounioux, 1954
 Type species. Emys capellinii Zigno, 1889
Type species. Emys capellinii Zigno, 1889
Neochelys zamorensis Jiménez Fuentes, 1993
Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9, Figure 10, Figure 11, Figure 12, Figure 13
Synonymy
'Neochelys’ zamorensis Arribas Moreno et al. (1984), Jiménez Fuentes (1986) [ nomen nudum ]
Neochelys zamorensis nomen nudum Jiménez Fuentes et al. (1985) [ nomen nudum ]
Neochelys aff. arenarum Mulas and Alonso Gavilán (1987), Mulas Alonso et al. (1988), Jiménez Fuentes et al. (1991, 1992)
Neochelys zamorensis Jiménez Fuentes and Gil Tudanca (1993), Pérez-García and de Lapparent de Broin (2013, 2015), Pérez-García (2017), Ortega et al. (2022), Pérez-García et al. (2023)
Type material. The holotype, STUS 12121, corresponding to an almost complete shell (Figure 2A-B).
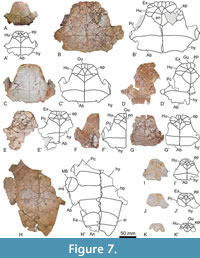
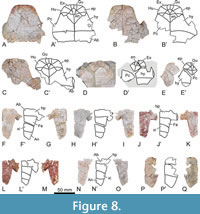
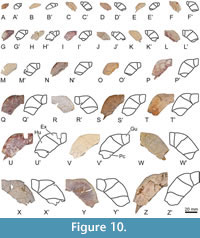
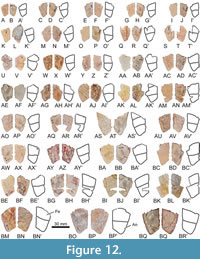
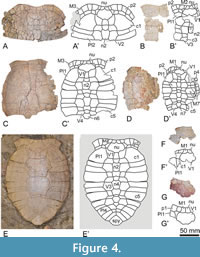 Referred material. A selection of 183 specimens, both from the type locality (Figure 2C-J) and from other outcrops of the same geological formation (i.e., the Entrala Formation) and province (Zamora Province) (Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9, Figure 10, Figure 11, Figure 12, Figure 13), corresponding to relatively complete to partial shells, and isolated plates, including: both articulated carapacial and plastral elements (Figure 2A-D, 2F-I, Figure 3A-H), partial carapaces (Figure 2E, Figure 4A-G, Figure 5A-H), complete and partial plastra (Figure 2J, Figure 6A-F, Figure 7A-K, Figure 8A-Q), nuchal plates (Figure 9A-Y), epiplastra (Figure 10A-Z), entoplastra (Figure 11A-AG), and xiphiplastra (Figure 12A-BR, Figure 13A-X). See the collection numbers in the respective figure captions.
Referred material. A selection of 183 specimens, both from the type locality (Figure 2C-J) and from other outcrops of the same geological formation (i.e., the Entrala Formation) and province (Zamora Province) (Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9, Figure 10, Figure 11, Figure 12, Figure 13), corresponding to relatively complete to partial shells, and isolated plates, including: both articulated carapacial and plastral elements (Figure 2A-D, 2F-I, Figure 3A-H), partial carapaces (Figure 2E, Figure 4A-G, Figure 5A-H), complete and partial plastra (Figure 2J, Figure 6A-F, Figure 7A-K, Figure 8A-Q), nuchal plates (Figure 9A-Y), epiplastra (Figure 10A-Z), entoplastra (Figure 11A-AG), and xiphiplastra (Figure 12A-BR, Figure 13A-X). See the collection numbers in the respective figure captions.
Type locality and horizon. El Escobar fossil site, Sanzoles Municipality, Zamora Province, Castile and Leon Autonomous Community, northwestern Spain (Figure 1). Duero Basin. Entrala Formation, MP 13 or MP 14, Lutetian, middle Eocene (see Ortega et al., 2022 and references therein).
Other localities. Valdegallina 1, Valdegallina 2, and Valdegallina 3 fossil sites, Valdegallina locality, Zamora Municipality; Cubillos 1 and Tardemayas 1 fossil sites, located respectively on the west and south margins of the Infiernos Hill, Monfarracinos Municipality; Teso de la Mora fossil site, Torres del Carrizal Municipality (Figure 1). As the type locality, all of them are located in the Zamora Province, Castile and Leon Autonomous Community, northwestern Spain. Duero Basin. Entrala Formation, MP 13 or MP 14, Lutetian, middle Eocene (see Ortega et al., 2022; and references therein).
 Emended diagnosis. Neochelys zamorensis can be defined by the following exclusive character combination of shell features within the genus Neochelys: elliptical carapace; maximum carapace length about 30 cm; slightly notched to unnotched anterior carapace margin; as wide as long to generally slightly wider than long nuchal; maximum nuchal width ranging between two and three times that of its anterior margin; seven or rarely eight neurals; longer than wide to slightly wider than long second and third neurals; slightly to almost twice wider than long suprapygal; slightly longer than wide to as wide as long pygal; slightly longer than wide to wider than long first peripheral; lyre-shaped first vertebral to development of substraight lateral margins; concave, convex or substraight first vertebral posterior margin; subequal width of the second and third vertebrals, both being the widest of the vertebral series; one and a half to two times as wide as long first marginal; anterior plastral lobe ranging from slightly wider than the posterior to slightly narrower than it; trapezoidal anterior plastral lobe; epiplastral posterior margin from as large as to two times larger than the anteromedial; entoplastral length from equivalent to its distance to the pectoral-abdominal sulcus to about two times greater; subrounded to generally substraight lateral margins of the posterior plastral lobe; anal notch width between two and two and a half times less than the maximum xiphiplastral width; anal notch two to four times wider than long; relatively long overlap of the gular on the entoplastron, reaching half the length of this plate, to almost absent overlap; presence or absence of a short to long gular-pectorals contact.
Emended diagnosis. Neochelys zamorensis can be defined by the following exclusive character combination of shell features within the genus Neochelys: elliptical carapace; maximum carapace length about 30 cm; slightly notched to unnotched anterior carapace margin; as wide as long to generally slightly wider than long nuchal; maximum nuchal width ranging between two and three times that of its anterior margin; seven or rarely eight neurals; longer than wide to slightly wider than long second and third neurals; slightly to almost twice wider than long suprapygal; slightly longer than wide to as wide as long pygal; slightly longer than wide to wider than long first peripheral; lyre-shaped first vertebral to development of substraight lateral margins; concave, convex or substraight first vertebral posterior margin; subequal width of the second and third vertebrals, both being the widest of the vertebral series; one and a half to two times as wide as long first marginal; anterior plastral lobe ranging from slightly wider than the posterior to slightly narrower than it; trapezoidal anterior plastral lobe; epiplastral posterior margin from as large as to two times larger than the anteromedial; entoplastral length from equivalent to its distance to the pectoral-abdominal sulcus to about two times greater; subrounded to generally substraight lateral margins of the posterior plastral lobe; anal notch width between two and two and a half times less than the maximum xiphiplastral width; anal notch two to four times wider than long; relatively long overlap of the gular on the entoplastron, reaching half the length of this plate, to almost absent overlap; presence or absence of a short to long gular-pectorals contact.
CHARACTERIZATION OF THE SHELL OF NEOCHELYS ZAMORENSIS
Description of the Shell of Neochelys zamorensis and Qualitative Approach to Its Intraspecific Variation
 A single specimen indisputably attributable to Neochelys zamorensis had until now been described and figured: the shell STUS 12121 (Figure 2A-B), corresponding to the holotype of the taxon, from the Lutetian fossil site of El Escobar (Sanzoles Municipality, Zamora Province; Figure 1). Although the potential presence of this taxon in other localities of the Zamora Province was referred, as well as the discovery of more material from its type locality (see Introduction), the absence of the indisputable attribution of more remains to this species had not allowed to know in detail its shell.
A single specimen indisputably attributable to Neochelys zamorensis had until now been described and figured: the shell STUS 12121 (Figure 2A-B), corresponding to the holotype of the taxon, from the Lutetian fossil site of El Escobar (Sanzoles Municipality, Zamora Province; Figure 1). Although the potential presence of this taxon in other localities of the Zamora Province was referred, as well as the discovery of more material from its type locality (see Introduction), the absence of the indisputable attribution of more remains to this species had not allowed to know in detail its shell.
The carapace of this species is elliptical. The shell length of the partially crushed Neochelys zamorensis holotype is about 21 cm (Figure 2A-B). However, specimens noticeably larger than it have been recognized, including one from its type locality with a shell more than one and a half times longer (Figure 2C-D), as well as others from several of the localities considered here (Figure 5F; Figure 6A; Figure 7B; Figure 10X-Z; Figure 11AE-AG).
 The nuchal plate of the holotype is about a quarter wider than long (Figure 2A). This plate is slightly wider than long in most specimens (Figure 2C; Figure 9R-S), and recognized as being as wide as long for scarce individuals (Figure 4G; Figure 9I). The relative dimensions of the nuchal do not depend on the size of the specimens since individuals of very different dimensions share the same proportions. The maximum nuchal width is slightly more than twice the anterior margin of this plate in the holotype (Figure 2A). A relatively large range of variation is observed for this character, the maximum width ranging between approximately twice (Figure 2C; Figure 9C, F, V), two and a half (Figure 3C; Figure 9A, K, N, X), and three times (Figure 4C; Figure 9G, T) that of the anterior margin. The holotype lacks an anterior nuchal notch, showing a substraight anterior margin. This structure is recognized as absent in some specimens (Figure 9C, J, Y) but not in others (Figure 2C; Figure 9O), showing a slightly concave margin. The holotype shows seven neural plates (Figure 2A), a character shared with all other specimens from the type locality in which this information is known (Figure 2E, H), but also with most individuals from the other localities (Figure 3A, C, E; Figure 4E; Figure 5B, F). However, the presence of eight neurals is identified for one individual (Figure 5G). The maximum width of the first neural is similar to that of its anterior margin in the case of the holotype (Figure 2A). It can be recognized as one and a half times greater (Figure 2C), or even twice greater (Figure 4C) than the width of its anterior margin when other specimens are considered. The maximum width of this plate can be located in the anterior area (as in the holotype; Figure 2A), middle region (Figure 2C), or in the posterior half of the plate (Figure 4C). The anterior margin of the plate is as markedly convex in the holotype (Figure 2A). It can be recognized as substraight (Figure 2E) and as markedly concave (Figure 4C) when other individuals are analyzed. In this sense, the lateral margins of the plate range from substraight (as in the holotype; Figure 2A) to markedly curved (Figure 4E). The second to fifth neurals of the holotype are slightly longer than wide (Figure 2A). A relatively high variation is recognized for the relative dimensions of these plates. In all cases, the width/length ratio decreases throughout the neural series, from the anterior to the posterior positions. However, these plates can also be slightly wider than long (Figure 4C) or as wide as long (Figure 3C), independently of the size of the individuals. The suprapygal of the holotype is almost twice wider than long (Figure 2A). It is recognized as slightly wider than long in most individuals, from both the type locality (Figure 2F, H) and other fossil sites (Figure 3A; Figure 4F; Figure 5H). The pygal of the holotype is as wide as long (Figure 2A), as in most specimens (Figure 2H), being slightly wider than long in others (Figure 3F). The first pair of peripherals of the holotype is wider than long (Figure 2A). These plates can also be recognized as being as wide as long (Figure 4E), and even slightly longer than wide (Figure 4B), when other individuals are studied.
The nuchal plate of the holotype is about a quarter wider than long (Figure 2A). This plate is slightly wider than long in most specimens (Figure 2C; Figure 9R-S), and recognized as being as wide as long for scarce individuals (Figure 4G; Figure 9I). The relative dimensions of the nuchal do not depend on the size of the specimens since individuals of very different dimensions share the same proportions. The maximum nuchal width is slightly more than twice the anterior margin of this plate in the holotype (Figure 2A). A relatively large range of variation is observed for this character, the maximum width ranging between approximately twice (Figure 2C; Figure 9C, F, V), two and a half (Figure 3C; Figure 9A, K, N, X), and three times (Figure 4C; Figure 9G, T) that of the anterior margin. The holotype lacks an anterior nuchal notch, showing a substraight anterior margin. This structure is recognized as absent in some specimens (Figure 9C, J, Y) but not in others (Figure 2C; Figure 9O), showing a slightly concave margin. The holotype shows seven neural plates (Figure 2A), a character shared with all other specimens from the type locality in which this information is known (Figure 2E, H), but also with most individuals from the other localities (Figure 3A, C, E; Figure 4E; Figure 5B, F). However, the presence of eight neurals is identified for one individual (Figure 5G). The maximum width of the first neural is similar to that of its anterior margin in the case of the holotype (Figure 2A). It can be recognized as one and a half times greater (Figure 2C), or even twice greater (Figure 4C) than the width of its anterior margin when other specimens are considered. The maximum width of this plate can be located in the anterior area (as in the holotype; Figure 2A), middle region (Figure 2C), or in the posterior half of the plate (Figure 4C). The anterior margin of the plate is as markedly convex in the holotype (Figure 2A). It can be recognized as substraight (Figure 2E) and as markedly concave (Figure 4C) when other individuals are analyzed. In this sense, the lateral margins of the plate range from substraight (as in the holotype; Figure 2A) to markedly curved (Figure 4E). The second to fifth neurals of the holotype are slightly longer than wide (Figure 2A). A relatively high variation is recognized for the relative dimensions of these plates. In all cases, the width/length ratio decreases throughout the neural series, from the anterior to the posterior positions. However, these plates can also be slightly wider than long (Figure 4C) or as wide as long (Figure 3C), independently of the size of the individuals. The suprapygal of the holotype is almost twice wider than long (Figure 2A). It is recognized as slightly wider than long in most individuals, from both the type locality (Figure 2F, H) and other fossil sites (Figure 3A; Figure 4F; Figure 5H). The pygal of the holotype is as wide as long (Figure 2A), as in most specimens (Figure 2H), being slightly wider than long in others (Figure 3F). The first pair of peripherals of the holotype is wider than long (Figure 2A). These plates can also be recognized as being as wide as long (Figure 4E), and even slightly longer than wide (Figure 4B), when other individuals are studied.
 The first vertebral of the holotype is about one and a half times wider than long (Figure 2A), as in some other specimens (e.g., Figure 3A). This scute is identified as slightly wider than long in other individuals (e.g., Figure 4A, C). The anterior margin of the first vertebral of the holotype is substraight, being antero-medially projected (Figure 2A). That projection is absent in other individuals, with a substraight margin (e.g., Figure 2C). A medially concave and laterally convex anterior margin is identified in other individuals (e.g., Figure 4A). The lateral margins of this scute can be substraight (Figure 2C; Figure 4E) or lyre-shaped (Figure 3A; Figure 4A) sensu Pérez-García and de Lapparent de Broin (2013) (i.e., with a slight curvature). The posterior margin of the first vertebral varies among the Neochelys zamorensis specimens, being from medially convex and laterally concave (as in the holotype; Figure 2A) to almost concave (Figure 2C), but also substraight in other individuals (Figure 4A). The second and third vertebrals are the widest in the vertebral series of this taxon (Figure 2A; Figure 3A, C; Figure 4F). The second vertebral of the holotype is recognized as slightly wider than long (Figure 2A). This ratio is variable for the taxon, specimens in which the scute is about one and a half times wider than long being also identified (Figure 4E). As in the holotype (Figure 2A), the anterolateral margins of the second vertebral of most specimens are slightly anteriorly curved (Figure 4E), being substraight in others (Figure 3C). A similar condition can be recognized for the posterolateral margins, being anteriorly curved (as in the holotype and, for example, in the specimen in Figure 4E) or substraight (Figure 5B). The third vertebral ranges from as wide as long (Figure 2A, E) to one and a half times wider than long (Figure 3A). A similar range of variation relative to the dimensions of the fourth and fifth vertebral scutes can also be recognized (e.g., as long as wide fourth vertebral is present in the specimens in Figure 2E and Figure 5H; this scute being noticeably wider than long in other individuals as in the holotype or, especially, in those in Figure 3A and Figure 4E). An additional anomalous scute is developed in one of the specimens, between the two most posterior vertebrals (Figure 3C). The first marginal of the holotype is almost one and a half times wider than long (Figure 2A). The ratio between these dimensions is recognized as variable when all specimens are analyzed. Thus, it ranges from specimens in which, as in the holotype, the width is approximately one and a half times the length (Figure 2C), to others where it is approximately twice the length (Figure 4F). A relatively long overlap of the first pair of marginals on the nuchal lateral margins is observed in the holotype, being developed along half the length of that margin (Figure 2A). This overlap varies among the specimens studied here, a similar condition to that in the holotype being observed for some individuals (Figure 2C), but only affecting the anterior third in others (Figure 9Y), intermediate stages being also recognized (Figure 4C). The posterior margin of the first pair of marginals of the holotype is concave (Figure 2A), but this area is recognized as substraight for some specimens, including another from the type locality (Figure 2C). The lateral margins of the first marginals are twice as long as the medial one in the case of the holotype (Figure 2A). A relatively high range of variability is identified for this character. Specifically, both margins can be subequal (Figure 4G); or the lateral margin can be from slightly larger (Figure 3A), to more than two times larger than the medial one (Figure 4A).
The first vertebral of the holotype is about one and a half times wider than long (Figure 2A), as in some other specimens (e.g., Figure 3A). This scute is identified as slightly wider than long in other individuals (e.g., Figure 4A, C). The anterior margin of the first vertebral of the holotype is substraight, being antero-medially projected (Figure 2A). That projection is absent in other individuals, with a substraight margin (e.g., Figure 2C). A medially concave and laterally convex anterior margin is identified in other individuals (e.g., Figure 4A). The lateral margins of this scute can be substraight (Figure 2C; Figure 4E) or lyre-shaped (Figure 3A; Figure 4A) sensu Pérez-García and de Lapparent de Broin (2013) (i.e., with a slight curvature). The posterior margin of the first vertebral varies among the Neochelys zamorensis specimens, being from medially convex and laterally concave (as in the holotype; Figure 2A) to almost concave (Figure 2C), but also substraight in other individuals (Figure 4A). The second and third vertebrals are the widest in the vertebral series of this taxon (Figure 2A; Figure 3A, C; Figure 4F). The second vertebral of the holotype is recognized as slightly wider than long (Figure 2A). This ratio is variable for the taxon, specimens in which the scute is about one and a half times wider than long being also identified (Figure 4E). As in the holotype (Figure 2A), the anterolateral margins of the second vertebral of most specimens are slightly anteriorly curved (Figure 4E), being substraight in others (Figure 3C). A similar condition can be recognized for the posterolateral margins, being anteriorly curved (as in the holotype and, for example, in the specimen in Figure 4E) or substraight (Figure 5B). The third vertebral ranges from as wide as long (Figure 2A, E) to one and a half times wider than long (Figure 3A). A similar range of variation relative to the dimensions of the fourth and fifth vertebral scutes can also be recognized (e.g., as long as wide fourth vertebral is present in the specimens in Figure 2E and Figure 5H; this scute being noticeably wider than long in other individuals as in the holotype or, especially, in those in Figure 3A and Figure 4E). An additional anomalous scute is developed in one of the specimens, between the two most posterior vertebrals (Figure 3C). The first marginal of the holotype is almost one and a half times wider than long (Figure 2A). The ratio between these dimensions is recognized as variable when all specimens are analyzed. Thus, it ranges from specimens in which, as in the holotype, the width is approximately one and a half times the length (Figure 2C), to others where it is approximately twice the length (Figure 4F). A relatively long overlap of the first pair of marginals on the nuchal lateral margins is observed in the holotype, being developed along half the length of that margin (Figure 2A). This overlap varies among the specimens studied here, a similar condition to that in the holotype being observed for some individuals (Figure 2C), but only affecting the anterior third in others (Figure 9Y), intermediate stages being also recognized (Figure 4C). The posterior margin of the first pair of marginals of the holotype is concave (Figure 2A), but this area is recognized as substraight for some specimens, including another from the type locality (Figure 2C). The lateral margins of the first marginals are twice as long as the medial one in the case of the holotype (Figure 2A). A relatively high range of variability is identified for this character. Specifically, both margins can be subequal (Figure 4G); or the lateral margin can be from slightly larger (Figure 3A), to more than two times larger than the medial one (Figure 4A).
 The anterior plastral lobe of the holotype of Neochelys zamorensis is slightly wider than the posterior one (Figure 2B), as in some other specimens (Figure 6F). The width of both lobes is subequal in some individuals analyzed here, or even the posterior one becomes slightly wider in others (Figure 3D, Figure 6A). The anterior plastral lobe of the holotype is trapezoidal, showing a straight anterior margin (Figure 2B). A trapezoidal morphology is shared with the rest of the specimens, the anterior margin being straight or substraight in most of them (Figure 2D; Figure 6C-D), although some show a poorly developed curvature of the anterior plastral margin (Figure 6A; Figure 7C). The lateral margins of the posterior plastral lobe of most specimens analyzed here, including the holotype (Figure 2B), are substraight. However, subrounded lateral margins have also been observed for other specimens (Figure 7H).
The anterior plastral lobe of the holotype of Neochelys zamorensis is slightly wider than the posterior one (Figure 2B), as in some other specimens (Figure 6F). The width of both lobes is subequal in some individuals analyzed here, or even the posterior one becomes slightly wider in others (Figure 3D, Figure 6A). The anterior plastral lobe of the holotype is trapezoidal, showing a straight anterior margin (Figure 2B). A trapezoidal morphology is shared with the rest of the specimens, the anterior margin being straight or substraight in most of them (Figure 2D; Figure 6C-D), although some show a poorly developed curvature of the anterior plastral margin (Figure 6A; Figure 7C). The lateral margins of the posterior plastral lobe of most specimens analyzed here, including the holotype (Figure 2B), are substraight. However, subrounded lateral margins have also been observed for other specimens (Figure 7H).
 The posterior epiplastral margin is approximately twice as long as the medial one in the holotype (Figure 2B). Variations in the dimensions of this ratio are identified considering all specimens studied here. In this sense, the posterior margin can be as wide as the medial one (Figure 10E, G, P, V), one and a half times (Figure 6D; Figure 10C, I, M), or two times (Figure 7A; Figure 10F, K, R) longer than it. The entoplastron is rhomboidal. In the case of the holotype, it is slightly longer than wide, a condition shared with some other individuals (Figure 11E, R). This plate is identified as being as wide as long (Figure 8B; Figure 11A, I, V, Y, AC), or slightly wider than long (Figure 2D, J; Figure 8B; Figure 11P, Z, AD) in others. Most entoplastra present substraight posterolateral margins (Figure 6A; Figure 11Y, P), as in the case of the holotype and the other specimens from the type locality in which this plate is preserved (Figure 2B, D, J), being slightly convex in only some of the analyzed individuals (Figure 7D; Figure 11 AD). The entoplastral length ranges from being approximately the same as its distance to the pectoral-abdominal sulcus (e.g., in the holotype or in other specimens from the type locality; Figure 2B, D) to about two times greater (Figure 3D; Figure 8A). It is equivalent to approximately two-thirds of its distance to the hyo-hypoplastral suture in the holotype and other individuals (Figure 2B; Figure 6A; Figure 7C), but its length is almost equivalent to that distance in some specimens (Figure 6D). The hyoplastra are longer than the hypoplastra for all analyzed shells. The mesoplastra are slightly longer than wide. The maximum width of the xiphiplastra of the holotype is slightly less than two times the width of the anal notch, as in some other specimens (Figure 6F). It is recognized as about two (e.g., Figure 6C) or approximately two and a half (Figure 6A) times the anal notch wide in other individuals. The anal notch is observed as slightly less than two and a half (Figure 13O), than three (as in the holotype, Figure 2B, and in other specimens as for example those in Figure 2B; Figure 6A, C), or than four (Figure 6F) times wider than long. The holotype displays a U-shaped anal notch (i.e., sexual morphotype I; Figure 2B), as that observed in some specimens (Figure 2I; Figure 7H; Figure 13U, W), whereas the anal notch of other individuals is V-shaped (i.e., sexual morphotype II; Figure 3D; Figure 12X, AZ). The morphology of the anal notch of most of the smallest specimens (Figure 12A-S) is more similar to that of the second morphotype.
The posterior epiplastral margin is approximately twice as long as the medial one in the holotype (Figure 2B). Variations in the dimensions of this ratio are identified considering all specimens studied here. In this sense, the posterior margin can be as wide as the medial one (Figure 10E, G, P, V), one and a half times (Figure 6D; Figure 10C, I, M), or two times (Figure 7A; Figure 10F, K, R) longer than it. The entoplastron is rhomboidal. In the case of the holotype, it is slightly longer than wide, a condition shared with some other individuals (Figure 11E, R). This plate is identified as being as wide as long (Figure 8B; Figure 11A, I, V, Y, AC), or slightly wider than long (Figure 2D, J; Figure 8B; Figure 11P, Z, AD) in others. Most entoplastra present substraight posterolateral margins (Figure 6A; Figure 11Y, P), as in the case of the holotype and the other specimens from the type locality in which this plate is preserved (Figure 2B, D, J), being slightly convex in only some of the analyzed individuals (Figure 7D; Figure 11 AD). The entoplastral length ranges from being approximately the same as its distance to the pectoral-abdominal sulcus (e.g., in the holotype or in other specimens from the type locality; Figure 2B, D) to about two times greater (Figure 3D; Figure 8A). It is equivalent to approximately two-thirds of its distance to the hyo-hypoplastral suture in the holotype and other individuals (Figure 2B; Figure 6A; Figure 7C), but its length is almost equivalent to that distance in some specimens (Figure 6D). The hyoplastra are longer than the hypoplastra for all analyzed shells. The mesoplastra are slightly longer than wide. The maximum width of the xiphiplastra of the holotype is slightly less than two times the width of the anal notch, as in some other specimens (Figure 6F). It is recognized as about two (e.g., Figure 6C) or approximately two and a half (Figure 6A) times the anal notch wide in other individuals. The anal notch is observed as slightly less than two and a half (Figure 13O), than three (as in the holotype, Figure 2B, and in other specimens as for example those in Figure 2B; Figure 6A, C), or than four (Figure 6F) times wider than long. The holotype displays a U-shaped anal notch (i.e., sexual morphotype I; Figure 2B), as that observed in some specimens (Figure 2I; Figure 7H; Figure 13U, W), whereas the anal notch of other individuals is V-shaped (i.e., sexual morphotype II; Figure 3D; Figure 12X, AZ). The morphology of the anal notch of most of the smallest specimens (Figure 12A-S) is more similar to that of the second morphotype.
 A poorly developed gular protrusion is present in the holotype (Figure 2B). This structure ranges from more developed in some specimens (Figure 2D; Figure 7B; Figure 10X) to absent or almost absent in others (Figure 2J; Figure 6C; Figure 10V). The gular of the holotype is two times longer than wide (Figure 2B). This ratio is identified as variable when the available specimens are compared, ranging from that condition to being slightly longer than wide (Figure 7D). Thus, the gular of most individuals is approximately one and a half times longer than wide (Figure 8A). The lateral margins of the gular scute on the epiplastra are subparallel in the holotype (Figure 2B). That condition is shared with some other specimens (Figure 2J; Figure 6A, E), being anteriorly divergent in most of them, also being substraight (Figure 2D; Figure 6D; Figure 8A) or curved, so that the maximum width of the specimens displaying this last condition is not reached in the most anterior region (Figure 7B; Figure 8C; Figure 10Y). The maximum width of each extragular of the holotype is one and a half times greater than that of the gular (Figure 2B), but specimens where the extragular wide is twice as small (Figure 3B), slightly less (Figure 7A), or the same (Figure 3D) as that of the gular are identified. The extragulars of the holotype reach the antero-lateral margin of the entoplastron (Figure 2B). However, the overlapping of the extragulars on the entoplastron is also identified as variable, with one (Figure 2D; Figure 3B; Figure 11AF), both (Figure 3D; Figure 11Z), or neither (Figure 7E; Figure 11AD) of these scutes overlapping this plate but, in the latter case, reaching the entoplastral anterolateral margin. The gular scute of the holotype overlaps over half the length of the entoplastron (Figure 2B). This is variable, many intermediate stages being identified, from that referred for this specimen to the almost absent overlapping in the shell from the type locality corresponding to Figure 2D. The gular reaches the pectoral scutes in the holotype (Figure 2B), as well as in some other specimens (Figure 3D; Figure 6A; Figure 7E; Figure 11AF), the length of that contact being variable. Other specimens show a medial contact between the humerals (Figure 2D; Figure 6F; Figure 7C; Figure 11AE), also of variable length, being equivalent to that of the gular on the entoplastron in some of them (Figure 11Y) to four times less than in others (Figure 11O). The humeral-pectoral sulcus of the holotype is substraight (Figure 2B), but it is recognized as laterally curved in some other individuals (Figure 7F). The most lateral point of the humeral-pectoral sulcus can be located on the posterolateral vertex of the epiplastron, as in the holotype (Figure 2B; Figure 6C; Figure 10N), on its lateral margin but away from the suture with the hyoplastron (Figure 3D; Figure 10S), or on the hyoplastron (Figure 2D; Figure 10W). The pectoral scutes can slightly overlap the mesoplastra (Figure 3B; Figure 6C) or, as in most individuals, not reach these plates (Figure 2D; Figure 3F). The anal scutes of all analyzed specimens are relatively short, not exceeding half the medial length of the xiphiplastra (Figure 2B, Figure 12).
A poorly developed gular protrusion is present in the holotype (Figure 2B). This structure ranges from more developed in some specimens (Figure 2D; Figure 7B; Figure 10X) to absent or almost absent in others (Figure 2J; Figure 6C; Figure 10V). The gular of the holotype is two times longer than wide (Figure 2B). This ratio is identified as variable when the available specimens are compared, ranging from that condition to being slightly longer than wide (Figure 7D). Thus, the gular of most individuals is approximately one and a half times longer than wide (Figure 8A). The lateral margins of the gular scute on the epiplastra are subparallel in the holotype (Figure 2B). That condition is shared with some other specimens (Figure 2J; Figure 6A, E), being anteriorly divergent in most of them, also being substraight (Figure 2D; Figure 6D; Figure 8A) or curved, so that the maximum width of the specimens displaying this last condition is not reached in the most anterior region (Figure 7B; Figure 8C; Figure 10Y). The maximum width of each extragular of the holotype is one and a half times greater than that of the gular (Figure 2B), but specimens where the extragular wide is twice as small (Figure 3B), slightly less (Figure 7A), or the same (Figure 3D) as that of the gular are identified. The extragulars of the holotype reach the antero-lateral margin of the entoplastron (Figure 2B). However, the overlapping of the extragulars on the entoplastron is also identified as variable, with one (Figure 2D; Figure 3B; Figure 11AF), both (Figure 3D; Figure 11Z), or neither (Figure 7E; Figure 11AD) of these scutes overlapping this plate but, in the latter case, reaching the entoplastral anterolateral margin. The gular scute of the holotype overlaps over half the length of the entoplastron (Figure 2B). This is variable, many intermediate stages being identified, from that referred for this specimen to the almost absent overlapping in the shell from the type locality corresponding to Figure 2D. The gular reaches the pectoral scutes in the holotype (Figure 2B), as well as in some other specimens (Figure 3D; Figure 6A; Figure 7E; Figure 11AF), the length of that contact being variable. Other specimens show a medial contact between the humerals (Figure 2D; Figure 6F; Figure 7C; Figure 11AE), also of variable length, being equivalent to that of the gular on the entoplastron in some of them (Figure 11Y) to four times less than in others (Figure 11O). The humeral-pectoral sulcus of the holotype is substraight (Figure 2B), but it is recognized as laterally curved in some other individuals (Figure 7F). The most lateral point of the humeral-pectoral sulcus can be located on the posterolateral vertex of the epiplastron, as in the holotype (Figure 2B; Figure 6C; Figure 10N), on its lateral margin but away from the suture with the hyoplastron (Figure 3D; Figure 10S), or on the hyoplastron (Figure 2D; Figure 10W). The pectoral scutes can slightly overlap the mesoplastra (Figure 3B; Figure 6C) or, as in most individuals, not reach these plates (Figure 2D; Figure 3F). The anal scutes of all analyzed specimens are relatively short, not exceeding half the medial length of the xiphiplastra (Figure 2B, Figure 12).
 The study of all the turtle shell remains from the Spanish El Escobar fossil site (Sanzoles Municipality, Zamora Province), corresponding to the type locality of Neochelys zamorensis, shows its compatibility with a single species. In fact, all these remains share an exclusive combination of features, allowing its attribution to a single taxon. The detailed analysis of the shell material from the other localities considered here, corresponding to the same geological formation (the Lutetian Entrala Formation) and area (the Spanish Zamora Province) than the type locality, also allows its attribution to the same species (see Discussion). Numerous unpublished and generally well-preserved skulls (more than 30) from this area are also identified, on different ontogenetic stages, from juveniles to fully developed individuals. They will be prepared for its future detailed study. However, their preliminary observation allows us to confirm their attribution to a single species (i.e., Neochelys zamorensis), which is compatible with the information provided by the shells.
The study of all the turtle shell remains from the Spanish El Escobar fossil site (Sanzoles Municipality, Zamora Province), corresponding to the type locality of Neochelys zamorensis, shows its compatibility with a single species. In fact, all these remains share an exclusive combination of features, allowing its attribution to a single taxon. The detailed analysis of the shell material from the other localities considered here, corresponding to the same geological formation (the Lutetian Entrala Formation) and area (the Spanish Zamora Province) than the type locality, also allows its attribution to the same species (see Discussion). Numerous unpublished and generally well-preserved skulls (more than 30) from this area are also identified, on different ontogenetic stages, from juveniles to fully developed individuals. They will be prepared for its future detailed study. However, their preliminary observation allows us to confirm their attribution to a single species (i.e., Neochelys zamorensis), which is compatible with the information provided by the shells.
Quantitative Approach to the Neochelys zamorensis Shell Intraspecific Variation
 An evaluation of the morphological variation of the shell elements of Neochelys zamorensis recognized as more variable and which presented a sample size appropriate to perform statistical analyses has been also done, in order to visualize the main directions of the shape changes. Therefore, a Principal Component Analysis (PCA) and a linear regression have been performed for the nuchals, epiplastra, entoplastra (see Figure S1, Figure S2, Figure S3, Figure S4, Figure S5, Figure S6 in Appendix 1), and xiphiplastra (see Figure 14, Figure 15, Figure 16 for the analysis of the xiphiplastral region).
An evaluation of the morphological variation of the shell elements of Neochelys zamorensis recognized as more variable and which presented a sample size appropriate to perform statistical analyses has been also done, in order to visualize the main directions of the shape changes. Therefore, a Principal Component Analysis (PCA) and a linear regression have been performed for the nuchals, epiplastra, entoplastra (see Figure S1, Figure S2, Figure S3, Figure S4, Figure S5, Figure S6 in Appendix 1), and xiphiplastra (see Figure 14, Figure 15, Figure 16 for the analysis of the xiphiplastral region).
 The PCA of morphogeometric data for the nuchal is illustrated in the space defined by the first three components (see Figure S1 in Appendix 1), which captures 79.5% of the total shape variance for this plate. The PC1 eigenvalue (39.86% of the total shape variance) principally concerns the relative dimensions of the nuchal, varying from relatively wide and long (positive scores; e.g., STUS 12121, Figure 2A-B) to narrow and short (negative scores; e.g., STUS 8751, Figure 9W). The shape variance kept by the PC2 (27.22% of the total shape variance; see Figure S1A in Appendix 1) is associated with the degree of development of the nuchal lateral margins. The positive end scores are occupied by a nuchal with long lateral margins (e.g., STUS 12061, Figure 4C), whereas a nuchal with relatively short lateral margins is located in the negative end scores (e.g., STUS 9123, Figure 9I). The PC3 (12.42% of the total shape variance; see Figure S1B in Appendix 1) is associated with the morphology of the posterolateral nuchal margin. The postero-lateral margins are more anteriorly curved as the positive values increase (e.g., STUS 12061, Figure 4C), whereas these margins are straighter as negative values increase (e.g., STUS 9108, Figure 9V).
The PCA of morphogeometric data for the nuchal is illustrated in the space defined by the first three components (see Figure S1 in Appendix 1), which captures 79.5% of the total shape variance for this plate. The PC1 eigenvalue (39.86% of the total shape variance) principally concerns the relative dimensions of the nuchal, varying from relatively wide and long (positive scores; e.g., STUS 12121, Figure 2A-B) to narrow and short (negative scores; e.g., STUS 8751, Figure 9W). The shape variance kept by the PC2 (27.22% of the total shape variance; see Figure S1A in Appendix 1) is associated with the degree of development of the nuchal lateral margins. The positive end scores are occupied by a nuchal with long lateral margins (e.g., STUS 12061, Figure 4C), whereas a nuchal with relatively short lateral margins is located in the negative end scores (e.g., STUS 9123, Figure 9I). The PC3 (12.42% of the total shape variance; see Figure S1B in Appendix 1) is associated with the morphology of the posterolateral nuchal margin. The postero-lateral margins are more anteriorly curved as the positive values increase (e.g., STUS 12061, Figure 4C), whereas these margins are straighter as negative values increase (e.g., STUS 9108, Figure 9V).
 The PCA of morphogeometric data for the epiplastron is illustrated in the space defined by the first three components (see Figure S2 in Appendix 1), which captures 75.25% of the total shape variance for this plate. The PC1 (34.95% of the total shape variance) is associated with the relative width of the plate, varying from relatively wide (positive scores; STUS 13633, Figure 10Y) to relatively narrow (negative scores; STUS 9115, Figure 10R). The PC2 eigenvalue (25.64% of the total shape variance, Figure S2A in Appendix 1) describes the relative width of the epiplastral posterior margin. Positive values are associated with a relatively wide posterior margin (e.g., STUS 12121, Figure 2A-B), whereas negative scores are related to a relatively narrow one (e.g., STUS 5754, Figure 10V). The shape variance kept by the PC3 (14.66% of the total shape variance; see Figure S2B in Appendix 1) is associated with the morphology of the anterior epiplastral margin. The positive end scores are occupied by an epiplastron with a straight anterior margin (e.g., STUS 10420, Figure 10N), whereas an epiplastron with a curved anterior margin is located in the negative end scores (e.g., STUS 8787, Figure 10W).
The PCA of morphogeometric data for the epiplastron is illustrated in the space defined by the first three components (see Figure S2 in Appendix 1), which captures 75.25% of the total shape variance for this plate. The PC1 (34.95% of the total shape variance) is associated with the relative width of the plate, varying from relatively wide (positive scores; STUS 13633, Figure 10Y) to relatively narrow (negative scores; STUS 9115, Figure 10R). The PC2 eigenvalue (25.64% of the total shape variance, Figure S2A in Appendix 1) describes the relative width of the epiplastral posterior margin. Positive values are associated with a relatively wide posterior margin (e.g., STUS 12121, Figure 2A-B), whereas negative scores are related to a relatively narrow one (e.g., STUS 5754, Figure 10V). The shape variance kept by the PC3 (14.66% of the total shape variance; see Figure S2B in Appendix 1) is associated with the morphology of the anterior epiplastral margin. The positive end scores are occupied by an epiplastron with a straight anterior margin (e.g., STUS 10420, Figure 10N), whereas an epiplastron with a curved anterior margin is located in the negative end scores (e.g., STUS 8787, Figure 10W).
 The PCA of morphogeometric data for the entoplastron is illustrated in the space defined by the first three components (see Figure S3 in Appendix 1), which captures 95.67% of the total shape variance for this plate. The PC1 eigenvalue (39.86% of the total shape variance) principally describes the relative dimensions of the plate. The entoplastra are shorter and wider as positive values increase (e.g., STUS 10500, Figure 11AD), whereas this plate is larger and narrower as negative values increase (e.g., STUS 12121, Figure 2A-B). The PC2 (34.02% of the total shape variance; see Figure S3A in Appendix 1) principally concerns the height at which the lateral vertex of the plate is located. Thus, the positive end scores are occupied by a lateral vertex located relatively anterior in the entoplastron (e.g., STUS 8764, Figure 11V), whereas this vertex is posteriorly ubicated in the plate in the negative values (e.g., STUS 10501, Figure 11X). The PC3 eigenvalue (18.24% of the total shape variance; see Figure S3B in Appendix 1) is associated with the degree of curvature of the postero-lateral entoplastral margins. Positive values are related to entoplastra with anteriorly curved postero-lateral margins (e.g., STUS 5756, Figure 11Z), whereas negative values are associated with entoplastra with posteriorly curved postero-lateral margins (e.g., STUS 14255, Figure 7B).
The PCA of morphogeometric data for the entoplastron is illustrated in the space defined by the first three components (see Figure S3 in Appendix 1), which captures 95.67% of the total shape variance for this plate. The PC1 eigenvalue (39.86% of the total shape variance) principally describes the relative dimensions of the plate. The entoplastra are shorter and wider as positive values increase (e.g., STUS 10500, Figure 11AD), whereas this plate is larger and narrower as negative values increase (e.g., STUS 12121, Figure 2A-B). The PC2 (34.02% of the total shape variance; see Figure S3A in Appendix 1) principally concerns the height at which the lateral vertex of the plate is located. Thus, the positive end scores are occupied by a lateral vertex located relatively anterior in the entoplastron (e.g., STUS 8764, Figure 11V), whereas this vertex is posteriorly ubicated in the plate in the negative values (e.g., STUS 10501, Figure 11X). The PC3 eigenvalue (18.24% of the total shape variance; see Figure S3B in Appendix 1) is associated with the degree of curvature of the postero-lateral entoplastral margins. Positive values are related to entoplastra with anteriorly curved postero-lateral margins (e.g., STUS 5756, Figure 11Z), whereas negative values are associated with entoplastra with posteriorly curved postero-lateral margins (e.g., STUS 14255, Figure 7B).
In addition to the study of the polymorphisms relative to the shape variation, those corresponding to the size variation have also been analyzed for Neochelys zamorensis. Shapiro-Wilk tests indicated, for all the shell elements of this species analyzed (i.e., nuchals, epiplastra, and entoplastra), p-values < 0.05. Therefore, we can affirm that the data are not distributed following a normal distribution in the sample.
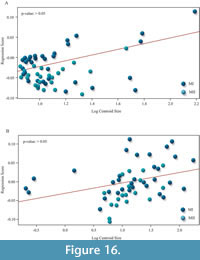
Only the epiplastron is influenced by allometry (p < 0.05; see Figure S4B in Appendix 1). Therefore, part of shape variation for this plate (i.e., 15%) can be explained by differences in size among specimens. The shape of the rest of the shell elements are not influenced by allometry (p > 0.05; see Figure S4A, S4C in Appendix 1).
Characterization of the Neochelys zamorensis Shell Sexual Dimorphism
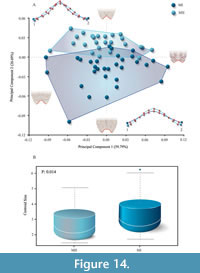
The PCA of morphogeometric data of the anal notch of Neochelys zamorensis is illustrated in the space defined by the first two components (see Figure 14A), which accounts for 86.48% of total shape variance of this region. The PC1 eigenvalue, which accounts for 59.79% of the variance, principally concerns the width of the anal notch. The width of the anal notch is higher as positive values increase, (e.g., STUS 9101, Figure 8H-I), whereas it is lower as negative scores increase (e.g., STUS 8747, Figure 13K-L). An evident distinction of the two morphotypes has not been observed along this x-axis. The shape variance kept by the PC2 (26.69% of the total shape variance) is mainly related to the morphology of the anal notch. The positive end scores are associated with a V-shaped anal notch (e.g., STUS 6107, Figure 12C-D), while negative end scores are associated with a U-shaped one (e.g., STUS 9098, Figure 13U-V). Thus, the y-axis evidence a distinction between the two sexual morphotypes described above (for more details see the section Description of the shell of Neochelys zamorensis and qualitative approach to its intraspecific variation).
The Procrustes ANOVA coordinates confirmed a significant difference regarding the anal notch area (p < 0.05) between sexes (Table 1). Also, the cross-validation test showed a high percentage of correctly classified morphotypes (87.60%).
Shapiro-Wilk tests indicated a not normal distribution (p > 0.05) for the dataset of the anal notch. Therefore, the validity of the parametric analyzes (i.e., regression) cannot be proved. The regression indicates that the anal notch of Neochelys zamorensis (Figure 16A) is not influenced by allometry (p > 0.05), where only 5.96% of the shape variation can be explained by differences in size among specimens. Likewise, this is also evidenced by Procrustes ANOVA (p > 0.05; for more details, see Table 1). The box plot graphically shows the differences in distribution between both anal notch morphotypes, being relatively separated with a low overlap (Figure 14B). T-test revealed not accentuated differences between the mean of both groups (p > 0.05).
The PCA of morphogeometric data for the xiphiplastral process is illustrated in the space defined by the first two components (Figure 15A), which captures 86.93% of the total shape variance for this structure. The PC1 (54.23% of the total shape variance) is associated with the relative width of the xiphiplastral process. The positive loadings are associated with an increase in the width of the xiphiplastral process (e.g., STUS 10063, Figure 2H-I), and vice versa (e.g., STUS 8746, Figure 12BI-BJ). This x-axis partially distinguishes the two previously established sexual morphotypes although, in this case, the overlap between the specimens of different morphotypes is higher than in the study of the anal notch. The shape variance kept by the PC2, which accounts for 32.70% of the variance, principally concerns the relative length and the degree of roundness of the xiphiplastral process. Therefore, the xiphiplastral process is larger and more pointed as positive values increase (e.g., STUS 5757, Figure 13M-N). By contrast, this structure is shorter and more rounded as negative values increase (e.g., STUS 6103, Figure 12BC, BD). There is no differentiation for the y-axis between morphotypes.
Procrustes ANOVA revealed no significant differences in shape (p > 0.05) (Table 2). Cross-validation test showed a high percentage of correctly classified morphotypes (85.43%).
As in the anterior cases, Shapiro-Wilk tests indicated a p < 0.05 for the dataset of the xiphiplastral process. Therefore, the data is not distributed following a normal distribution in the sample. The regression indicates that the xiphiplastral process of Neochelys zamorensis (Figure 16B) is not influenced by allometry (p > 0.05), where only 7.16% of the shape variation can be explained by differences in size among specimens. Procrustes ANOVA coordinates confirmed no significant difference regarding the xiphiplastral processes area (p > 0.05) between sexes (Table 2). The box plot of the centroid size variation for each sex is represented in Figure 15B. This demonstrates no accentuated differences between the mean of both morphotypes. Likewise, T-test evidenced no significant differences between both groups (p > 0.05).
DISCUSSION
Sexual dimorphism approximation to the sexual dimorphism in extant podocnemidids. The PCA of morphogeometric data of the anal notch including several species of extant podocnemidids evidenced the same shape changes that the anterior PCA of this area in Neochelys zamorensis (see Figure S6A in Appendix 1). The first two principal components account for 91.76% of the total variance in the dataset. As described before, the PC1 (76.14% of the total shape variance) is principally associated with the width of the anal notch. Unlike the specimens of Neochelys zamorensis, the x-axis evidenced a distinction between the two sexes of the analyzed species of extant podocnemidids. Thus, in most species (i.e., except for Podocnemis lewyana), males tend to have more negative scores, which is associated with a narrower anal notch. By contrast, females have more positive scores, which is related to a wider opening in the anal region.
The PC2 eigenvalue, which accounts for 15.62% of the variance, coincides with the shape differences described before for this axis (i.e., the morphology of the anal notch). As in the PC1, the two sexual morphotypes of extant podocnemidids are clearly differentiated. In this case, males have more negative values than females, which is related to a U-shaped anal notch. The only exception is in Erymnochelys madagascariensis, in which the male is located in slightly more positive values than the female. By contrast, females of all the species of Podocnemis analyzed present a more positive values than males, which is associated with a V-shaped anal notch. The combination of the PC1 and the PC2 of the specimens of extant turtles achieves a good separation of sexual dimorphs. However, in Neochelys zamorensis, this separation only occurs when considering the PC2. Despite these differences, the male specimens of the species of extant podocnemidids are more similar to the first sexual morphotype of Neochelys zamorensis, whereas the females are closer to the anal notch morphology of the second one.
The PCA of morphogeometric data of the xiphiplastral processes including several species of extant podocnemidids evidenced the similar shape changes than the PCA for this region in Neochelys zamorensis (see Figure S6B in Appendix 1). In this case, the first two principal components account for 87.97% of the total variance in the dataset. The shape variance kept by the PC1 (56.08% of the total shape variance) is mainly related to the relative width of the region. The most positive values are occupied for some of the males of extant podocnemidids (i.e., Podocnemis expansa, Podocnemis lewyana, and Podocnemis unifilis), which is related to a relatively narrow xiphiplastral process. By contrast, most females tend to present more negative values on the x-axis. In some species (e.g., in Podocnemis expansa and Podocnemis unifilis), the values between males and females are very different, both sexes being relatively separated along this axis. By contrast, in other species (e.g., Erymnochelys madagascariensis or Podocnemis vogli) the differences between them are very subtle. The PC2 (31.89% of the total shape variance) is associated with the relative length and the degree of roundness of the xiphiplastral process, as observed in the analyzes conducted on this region in Neochelys zamorensis. In this case, the males of the other species (except for Podocnemis lewyana and Podocnemis volgli) present more negative values, which are related to shorter and more rounded xiphiplastral processes. By contrast, several females of the extant podocnemidids analyzed here are located on more positive values, associated with a larger and more pointed xiphiplastral process. For this area, no obvious trend has been observed, combining the main axes of shape variation, to characterize the sexual dimorphs of extant podocnemidids.
The combination of the results of both analyses of the xiphiplastral region evidenced that three species (i.e., Podocnemis expansa, Podocnemis lewyana, and Podocnemis unifili s) coincide in that the males present a U-shaped anal notch with a narrower xiphiplastral process. The males of Podocnemis vogli also coincide with a U-shaped anal notch; however, the morphology of the xiphiplastral processes of this sexual morphotype is similar to that of females, being relatively wide. Erymnochelys madagascariensis does not present significant differences between both sexual dimorphs, the differences being located in the xiphiplastral region in this analysis. Only some of the species analyzed here (i.e., Podocnemis expansa, Podocnemis unifilis, and Erymnochelys madagascariensis) coincide with the trend of the sexual dimorphs of Neochelys zamorensis described above (i.e., the males of these three species have more negative values on the two axes, as in the morphotype I of Neochelys zamorensis).
Sexual dimorphism in Neochelys zamorensis. Sexual dimorphism in the turtle shell can manifest in several ways, the most obvious being the difference in size between adult males and females (Pritchard, 1979). In some species the degree of sexual dimorphism at the maturity may be very high (e.g., Graptemys pseudogeographica; Vetter, 2004); however, it can be barely distinguishable in others (e.g., Erymnochelys madagascariensis; Garcia, 2005). In terms of morphology, the more common sexual shell variations are related to the concavity of the plastron in males (e.g., in terrestrial species, Castellano et al., 2013), the overall shape of the carapace (e.g., it can be relatively rounded or domed in females, and elongated and more flattened in males; Keswick and Hofmeyr, 2015), or the morphology of the xiphiplastral region (Castaño-Mora et al., 2005).
The evaluation of the sexual shape dimorphism (SShD) in Neochelys zamorensis has been performed through the analysis of the morphological differences of the xiphiplastral area, since other dimorphic features observed in extant turtles (e.g., the plastron concavity) have not been included due to the nature of the sample (see materials and methods for more details). Thus, as in previous studies of extinct (e.g., Cadena et al., 2013; Limaverde et al., 2020) and extant turtles (e.g., Djordjevic et al., 2011; Sepúlveda-Seguro et al., 2020), the depth, the width, and the morphology of the anal notch, but also the morphology of the xiphiplastral processes, have been analyzed to infer the sexual dimorphism in Neochelys zamorensis.
The application of geometric morphometry improved the visualization and the distinction of the sexual morphotypes previously identified through the qualitative analyses. Although several intermediate forms are present in the sample, two main morphotypes (identified as the sexual morphotypes) can be distinguished on the basis of the morphology of the anal notch. As reported before (Guerrero and Pérez-García, 2021a), the study of extant pleurodires evidenced that both the direction and the magnitude of the sexual shape dimorphism are variable at generical level. Thus, the anal notch of representatives of different clades, but also of different genera of the same clade, belonging to the same sex, can be V-shaped (e.g., females of the genus of Podocnemididae Podocnemis; Páez et al., 2013; González-Zárate, 2014; Gallego-García and Forero-Medina, 2014; Sepúlveda-Seguro et al., 2020) or U-shaped (e.g., females of the genus of Chelidae Chelodina Fitzinger, 1826 and of that of Pelomedusidae Pelusios, Host, 2006; Gerlach, 2008). Regarding extant podocnemidids, but not in the two other mentioned clades (i.e., Chelidae and Pelomedusidae), the direction of the sexual shape dimorphism of the anal notch is maintained among all the representatives of the clade. Specifically, this region has been described in males as U-shaped and in females as V-shaped (i.e., in Peltocephalus, Parra-Henao et al., 2019; and in Podocnemis, González-Zárate, 2014; Páez et al., 2013; Gallego-García and Forero-Medina, 2014; Sepúlveda-Seguro et al., 2020). In this context, the U-shaped of the males of extant podocnemidids coincides with the first sexual morphotype described for Neochelys zamorensis, whereas the V-shaped of females is similar to the second morphotype. Regarding the xiphiplastral processes, no significant differences have been evidenced between the two sexual forms of Neochelys zamorensis; consequently, in this case, this character cannot be used to distinguish morphotypes.
Sexual size dimorphism (SSD) is also present the extant turtles ecologically similar to Neochelys (i.e., freshwater taxa; Rueda-Almonacid et al., 2007). As occurs with SShD, the direction and magnitude of this factor varies without following a phylogenetic pattern (e.g., being opposite in the chelids Platemys Wagler, 1830 and Emydura Bonaparte, 1836; Ernst, 1986; Trembath and Freier, 2004; and the pelomedusids Pelomedusa and Pelusios; Pritchard, 1979; Bour, 1983; Strydom, 2001; Broadley and Boycott, 2009; Böhm, 2010; Bour et al., 2016). Unlike the SShD, sexual size dimorphism is not maintained in the clade Podocnemididae. Specifically, in the podocnemidids Peltocephalus Duméril and Bibron, 1835,, and Erymnochelys Baur, 1888 the males get larger sizes than the females (i.e., males-biased; Garcia, 2005; Gentil et al., 2021), whereas females in Podocnemis reach larger sizes (e.g., females-biased; Rueda-Almonacid et al., 2007; González-Zárate et al., 2014).
The specimens that reach greater sizes in Neochelys zamorensis are those that correspond to the morphotype I. This data has been evidenced through the qualitative analysis (e.g., Figure 13W-X) but also quantitatively (i.e., with the box plots). The combination of the results previously obtained (i.e., SShD) with those related to SSD demonstrate that the morphotype I of Neochelys zamorensis coincides with the males of some representatives of Podocnemididae. Specifically, the morphotype I of Neochelys zamorensis presents the same SSD direction that Peltocephalus and Erymnochelys turtles, since the male sex has larger dimensions (Garcia, 2005; Gentil et al., 2021). By contrast, the morphotype I differs in the direction of the SSD from the genus Podocnemis, where the males are smaller than the females (Rueda-Almonacid et al., 2007).
As justified anteriorly (Guerrero and Pérez-García, 2021a), define sexual morphotypes in extinct species becomes relatively complex, since the direction but also the magnitude of the sexual dimorphism varies without a clear phylogenetic pattern. Therefore, caution must be used when inferring sexual dimorphs in extinct turtles, especially in the most basal taxa. As a consequence, on the basis of the comparisons of the SShD and the SSD of extant podocnemidids, the sexual morphotype I of Neochelys zamorensis has been identified as putative males and the sexual morphotype II as putative females. However, it is an attribution that cannot be confirmed with total certainty.
Sexual dimorphism in the genera Neochelys. The sexual dimorphism in the genera Neochelys has been poorly explored due to the limited sample size of most of the species (e.g., Neochelys eocaenica or Neochelys liriae). Thus, this typology of intraspecific variation has been only evaluated in Neochelys salmanticensis (Pérez-García et al., 2023), where a significant sexual dimorphism has not been identified. Although it has not been described and analyzed in detail, two clearly defined morphotypes of anal notch are present in several species of Neochelys (i.e., Neochelys arenarum, Neochelys eocaenica, Neochelys franzeni, and Neochelys laurenti), coincident with those reported here for Neochelys zamorensis and, therefore, with those of extant podocnemidids. For the moment, two clear anal notch morphotypes have not been observed in Neochelys capellini, Neochelys liriae, and Neochelys salmanticensis. However, this could vary in some of them (i.e., in the poorly represented Neochelys capellini and, especially, Neochelys liriae) with an increase in the sample size.
Most Neochelys species with two potential sexual dimorphs (i.e., Neochelys eocaenica, Neochelys franzeni, and Neochelys laurenti) does not present significant differences in the size between both morphotypes. The morphotype with a U-shaped morphology of the anal notch are slightly larger than the other form in Neochelys arenarum. Therefore, this coincides with the direction of both SShD and SSD of Neochelys zamorensis. However, a greater sample size of most Neochelys species is needed to confirm this idea. Consequently, based on the presently available specimens of the genus Neochelys, two similar morphotypes to those described in Neochelys zamorensis have been identified in some Neochelys species. This similarity encompasses attributes like the morphology of the anal notch and the size of the shell, with the latter displaying no significant variation.
Ontogeny of Neochelys Zamorensis
The variation result of the ontogenetic variability has also been evaluated due to the availability of numerous specimens, of different sizes, of Neochelys zamorensis. In this context, the features commonly dependent on the ontogenetic development in other Neochelys members (e.g., Neochelys franzeni, Cadena, 2015; or Neochelys salmanticensis, Pérez-García et al., 2023), as well as in extant podocnemidids (e.g., in the genus Podocnemis), have been analyzed in detail for Neochelys zamorensis. As occurs in other Neochelys species with the presence of juvenile individuals (i.e., Neochelys franzeni and Neochelys salmanticensis), the ontogenetical morphological variation in Neochelys zamorensis does not correspond to the principal factor of variation among the specimens. In fact, there is not a significant variation in the shell elements among the individuals of different sizes. Thus, other features identified in other Neochelys species as dependent on the ontogeny (i.e., the relative dimensions of vertebral scutes or the presence of a medial keel; Cadena, 2015; Pérez-García, 2015; Pérez-García et al., 2023), as well as in extant podocnemidids (e.g., Rueda-Almonacid et al., 2007; Gallego-García and Forero-Medina, 2014) are invariable among the specimens of Neochelys zamorensis. The lack of an ontogenetic trend in the plates and scutes in Neochelys zamorensis suggest an early ontogenetic development of the shell structures of the juveniles, relatively early reaching the adult status. Therefore, the sole determinant that distinguishes ontogenetic stages is the size of the individuals. Thus, both juvenile and adult individuals are recognized. The identification of intermediate ontogenetic stages (e.g., subadult individuals) for Neochelys zamorensis proves challenging due to the complexity introduced by the size differences, which may also be influenced by the sexual size dimorphism. This is the case of the holotype (Figure 2A-B) whose size is approximately two-thirds that of some other specimens of Neochelys zamorensis, including individuals also from the type locality (e.g., Figure 2C-D). This difference in size could be the result of the ontogenetic variation, the holotype being, therefore, a not fully developed adult. However, the anal notch of the holotype aligns with that of the second sexual morphotype recognized in this work, corresponding to the smaller one. In this context, the xiphiplastral length in the larger specimen recorded for this morphotype (Figure 13E-F) is approximately 35% greater than that of the holotype (Figure 2A-B). Consequently, this evidence suggests that the holotype of Neochelys zamorensis did not reach the maximum size observed for this morphotype, its relatively small size being due, at least in part, to ontogenetic factors.
Interspecific Variation for the Genus Neochelys
Generalities of the shell. As indicated above (see Introduction), Jiménez Fuentes (1993) proposed a differential diagnosis for Neochelys zamorensis in relation to Neochelys arenarum. The analysis of the abundant material of Neochelys zamorensis analyzed here, together with the first-hand study of the holotype and of all the previously published Neochelys arenarum shell remains (see Broin, 1977), as well as that of all the other unpublished specimens from its type locality (Rians, in France) deposited in the Muséum national d’Histoire naturelle (Paris, France) allow the evaluation of the features proposed by Jiménez Fuentes (1993). Jiménez Fuentes (1993) indicated that there were differences in the bone surface ornamentation pattern of both taxa, Neochelys zamorensis being smooth but costulations being present in the French species. Differences relative to this aspect are not recognized between both taxa: probably the costulations referred by Jiménez Fuentes (1993) for Neochelys arenarum correspond to irregularities in the shell due to the growth, and they are also recognized for Neochelys zamorensis (Figure 4A, Figure 5F). Jiménez Fuentes (1993) considered the length of the entoplastron of Neochelys zamorensis as approximately equivalent to its distance to the pectoral-abdominal sulcus, being at least twice it in Neochelys arenarum. As has been indicated above (see Description), the analysis of the unpublished remains of Neochelys zamorensis allows us to recognize a wide range of variability for this character, the condition indicated by Jiménez Fuentes (1993) for Neochelys arenarum being compatible with that in some individuals of the Spanish species. Jiménez Fuentes (1993) indicated that the length of the entoplastron was lower than that between that plate and the hyo-hypoplastral suture in the Spanish species, both lengths being subequal in the French species. However, the entoplastral length represents approximately two-thirds of the distance between this plate and the hyo-hypoplastral suture in the holotypes of both Neochelys arenarum and Neochelys zamorensis. As indicated above (see Description), variability is recognized for the Spanish species, the entoplastral length being almost equivalent to that distance in some Neochelys zamorensis individuals. Jiménez Fuentes (1993) characterized the morphology of the carapace of the French species as more rectangular than that of the Spanish form, this difference not being supported here. Jiménez Fuentes (1993) pointed out that the number of neural plates was different between both taxa, the French species having six and the Spanish having seven. Although the study of new material allows us to recognize variability for the number of neurals of Neochelys zamorensis, they vary between seven (in most specimens) and eight, this number of plates being, therefore, greater than that until now recognized for Neochelys arenarum. Therefore, this last character does allow us to characterize both diachronic species as different. But, in addition, other different character states between both taxa are recognized here. Thus, Neochelys zamorensis is compared in detail below, not only with that French representative, but also with all known species for this genus.
As recently reported by Pérez-García et al. (2023), the carapace morphology of almost all Neochelys species is elliptical, except in Neochelys liriae, which is subpentagonal. That general condition coincides with the carapace of Neochelys zamorensis. The maximum length observed among the carapaces of Neochelys zamorensis individuals, about 30 cm (e.g., Figure 3A), is much smaller than that reported for the other representative from the Spanish Duero Basin (i.e., Neochelys salmanticensis, reaching 50 cm), and smaller than those of Neochelys capellinii, Neochelys eocaenica, and Neochelys laurenti, whose reported maximum length is larger than 35 cm (Pérez-García and de Lapparent de Broin, 2013). Only Neochelys arenarum, Neochelys liriae, and Neochelys franzeni shows a carapace length shorter than that of Neochelys zamorensis, being about 25 cm (Pérez-García and de Lapparent de Broin, 2013).
The presence of the anterior plastral lobe of some individuals of Neochelys zamorensis slightly narrower than the posterior one is shared with Neochelys capellinii and Neochelys laurenti. An anterior plastral lobe slightly wider than the posterior one, as that observed in other specimens of Neochelys zamorensis, is shared with Neochelys salmanticensis, Neochelys arenarum, Neochelys franzeni, and Neochelys liriae. The presence of a trapezoidal anterior plastral lobe in Neochelys zamorensis is exclusively shared with Neochelys arenarum. As mentioned by Pérez-García et al. (2023) based on the holotype and so far only known individual of the species, the lateral margins of the posterior plastral lobe in Neochelys zamorensis are straight (e.g., Figure 3D). However, subrounded lateral margins have also been observed for several specimens of this taxon in the present contribution (e.g., Figure 7H), a condition which coincides with that in all other species, except in some individuals of Neochelys laurenti.
Carapacial plates. Neochelys zamorensis exhibits some variation in the relative dimensions of the nuchal plate (this plate ranging from slightly wider than long to as wide as long), as in almost all Neochelys representatives (i.e., Neochelys arenarum, Neochelys capellinii, Neochelys eocaenica, Neochelys franzeni, and Neochelys laurenti). The only exceptions are Neochelys salmanticensis, whose nuchal can also be slightly longer than wide (Pérez-García et al., 2023); and Neochelys liriae, in which this plate is exclusively recognized as one and a half times wider than long. The variation observed in the maximum nuchal width relative to its anterior margin in Neochelys zamorensis (i.e., the former ranging from about two to three times larger than that of the anterior margin) is less than that observed in Neochelys salmanticensis (i.e., ranging from one and a half to more than four times greater). However, the range of variability of this feature in Neochelys zamorensis surpasses that so far recognized for all other species (i.e., Neochelys arenarum, Neochelys capellinii, Neochelys eocaenica, Neochelys franzeni, Neochelys laurenti, and Neochelys liriae), in which the maximum nuchal width is always less than two and a half times that of the anterior margin. As previously reported (see Pérez-García et al., 2023, and references therein), some specimens of Neochelys salmanticensis present a well-developed notch in the anterior margin of the nuchal. The presence of this structure occurs in Neochelys zamorensis (e.g., Figure 9O); however, it is less pronounced than that developed in some individuals of Neochelys salmanticensis. This latter condition (i.e., a not significant nuchal notch) is shared with the other of Neochelys species. As described before, the anterior margin of the nuchal plate of Neochelys zamorensis can be substraight or slightly concave. This first condition coincides with that observed in Neochelys arenarum, Neochelys franzeni, and some specimens of Neochelys capellinii, Neochelys eocaenica, and Neochelys laurenti. By constrast, Neochelys zamorensis shares a slightly concave anterior margin of the nuchal plate with some individuals of Neochelys capellinii, Neochelys eocaenica, and Neochelys laurenti. Neochelys liriae is the only species in which this character is unknown due to the preservation of the holotype and only known specimens.
As in Neochelys capellinii, Neochelys laurenti, Neochelys liriae, Neochelys franzeni, and most Neochelys salmanticensis specimens (for more details see Pérez-García et al., 2023), most Neochelys zamorensis individuals present seven plates in its neural series. However, variations have been documented in Neochelys zamorensis, where the neural series can extend to eight plates, as observed in Neochelys eocaenica and certain specimens of Neochelys salmanticensis. Neochelys zamorensis presents a larger variation in the relative dimensions of the neural plates, specifically from the second to the fifth, than that identified for the other species of the genus. Thus, the presence of slightly wider than long or as wide as long second and third neurals is shared with Neochelys salmanticensis, Neochelys arenarum, Neochelys eocaenica, and Neochelys franzeni; but they can also be slightly longer than wide in the species studied here, as in Neochelys capellinii. By contrast, the second and third neurals of Neochelys laurenti and Neochelys liriae are significantly wider than long.
The presence of a suprapygal plate about two times wider than long in some specimens of Neochelys zamorensis is shared with Neochelys arenarum and Neochelys franzeni, those recognized as slightly wider than long in other individuals of this Spanish form being shared with Neochelys capellinii, Neochelys eocaenica, Neochelys laurenti, Neochelys liriae, and Neochelys salmanticensis.
The relative dimensions of the pygal plate of some individuals of Neochelys zamorensis, being as long as wide, coincide with those in Neochelys liriae. The identification of a slightly longer than wide pygal in other individuals of Neochelys zamorensis is shared with Neochelys salmanticensis and some specimens of Neochelys franzeni.
The slightly longer than wide first peripheral plate identified in some specimens of Neochelys zamorensis is shared with almost all Neochelys species. The presence of a wider than long first peripheral is exclusively shared with Neochelys liriae. However, the identification of this plate as being as wide as long in other individuals represents a so far unknown condition for this genus.
Carapacial scutes. The almost straight anterior margin of the first vertebral scute of some Neochelys zamorensis specimens, with anteriorly diverging margins, represents a condition shared with some specimens of Neochelys capellinii, Neochelys eocaenica, and Neochelys laurenti. The presence of a substraight margin in other individuals is shared with Neochelys arenarum. The condition recognized for other carapaces of Neochelys zamorensis, where this margin is medially concave and laterally convex, is shared with Neochelys liriae and Neochelys franzeni, but also with some Neochelys capellinii, Neochelys eocaenica, and Neochelys laurenti specimens. The presence of substraight lateral margins reported for the first vertebral of some individuals of Neochelys zamorensis is shared with Neochelys capellinii and some specimens of Neochelys franzeni. The condition observed in other individuals of Neochelys zamorensis, with lyre-shaped lateral margins, is shared with Neochelys arenarum, Neochelys eocaenica, Neochelys laurenti, Neochelys liriae, Neochelys salmanticensis, and some specimens of Neochelys franzeni. The condition recognized for some specimens of Neochelys zamorensis, with a substraight posterior margin of the first vertebral scute, is shared with Neochelys salmanticensis, Neochelys capellinii, Neochelys eocaenica, Neochelys liriae, and some specimens of Neochelys laurenti; that corresponding to a concave margin is like that in Neochelys arenarum, Neochelys franzeni, and some specimens of Neochelys laurenti; and that represented by a convex margin is the same as in some specimens of Neochelys franzeni. As reported by Pérez-García and de Lapparent de Broin (2013), and Pérez-García et al. (2023), a slightly wider than long first vertebral is recognized for all Neochelys species. Neochelys capellinii is the only species of Neochelys in which the first vertebral can also be slightly longer than wide or as wide as long. Therefore, the condition observed for other individuals of Neochelys zamorensis in which this scute is much wider (i.e., one and a half times) than long, had never been documented for this genus. The identification of the second and third vertebrals as the widest in the vertebral series is shared with all other Neochelys representatives except Neochelys eocaenica and Neochelys liriae, in which the widest vertebral is the last one.
As reported by previous authors, the relative dimensions of the first marginal scute are variable among the representatives of Neochelys, ranging from as wide as long to three times wider than long (for more details see Pérez-García and de Lapparent de Broin, 2013; Pérez-García et al., 2023). The relative dimensions of the first marginal in some Neochelys zamorensis individuals coincide with those in Neochelys arenarum and some specimens of Neochelys laurenti, the scute being about one and a half times wider than long. The condition identified for other specimens of Neochelys zamorensis, in which this scute was two times wider than long, coincides with that in Neochelys eocaenica, Neochelys capellinii, Neochelys franzeni, and some carapaces of Neochelys laurenti. As documented by Pérez-García et al. (2023), the ratio width/length of this scute in Neochelys salmanticensis is lower than that in the other Neochelys species, being as wide and as long or slightly wider than long. Neochelys liriae is the only species in which this character is unknown. As described above, a relatively high variation has been observed in the relative dimensions of the lateral margin of the first marginal compared with the medial one in Neochelys zamorensis. In this sense, the variation of this feature in other Neochelys representatives falls within the range of variation observed in Neochelys zamorensis. Specifically, the medial margin of this scute in the other species was reported as longer than the lateral one (i.e., in Neochelys capellinii, Neochelys eocaenica, Neochelys franzeni, Neochelys liriae, and some Neochelys laurenti specimens), or subequal to it (i.e., in Neochelys arenarum, Neochelys salmanticensis, and some Neochelys laurenti specimens; for more details see Pérez-García and de Lapparent de Broin, 2013). As previously described (i.e., see Pérez-García et al., 2023) the nuchal length in relation to the antero-posterior overlap of the first pair of marginals is quite variable among the Neochelys species. In this regard, this value can be about two-and-a-half times the medial margin of the marginals, as in some specimens of Neochelys salmanticensis; about three times, as in Neochelys arenarum, Neochelys liriae, and some Neochelys laurenti and Neochelys salmanticensis specimens; about five times, as in Neochelys capellinii, Neochelys eocaenica, Neochelys franzeni; and about six times, as in some specimens of Neochelys laurenti. In this context, the variation in this character in most species of Neochelys falls within the relative dimensions recognized for Neochelys zamorensis, in which the nuchal length ranges from about two and a half times the medial margin of the marginals to about five times (for more details see Pérez-García and de Lapparent de Broin, 2013; Pérez-García et al., 2023). The only exception are some Neochelys laurenti specimens, in which the nuchal length can be six times that of the overlapping of the first marginal scute on this plate.
Plastral plates. Some individuals of Neochelys zamorensis shares with the other species of Neochelys a slightly wider than long rhomboidal entoplastron. However, both the conditions of displaying an as wide as long entoplastron and a slightly longer than wide plate recognized for other specimens of this Spanish form are not shared with any other representative of the genus. The presence of substraight entoplastral posterolateral margins is shared with Neochelys arenarum, Neochelys eocaenica, Neochelys franzeni, Neochelys liriae, and some specimens of Neochelys laurenti and Neochelys salmanticensis; those slightly convex recognized for other carapaces of Neochelys zamorensis being shared with Neochelys capellini, and other individuals of Neochelys laurenti and Neochelys salmanticensis. The presence of an entoplastral length about two times the distance of this plate to the pectoral-abdominal sulcus in some individuals of Neochelys zamorensis is shared with Neochelys arenarum, Neochelys liriae, Neochelys capellinii, and Neochelys laurenti. By contrast, the entoplastral length is less than twice that distance in Neochelys eocaenica, Neochelys franzeni, and Neochelys salmanticensis. All other representatives of Neochelys present less variation than that in Neochelys zamorensis regarding the dimensions of the epiplastral posterior margin in relation to the anteromedial one. Thus, the range of variation in Neochelys zamorensis allows to recognize individuals with the posterior margin as wide as the anteromedial one (a condition shared with some specimens of Neochelys salmanticensis), slightly larger (as in Neochelys eocaenica and some individuals of Neochelys arenarum, Neochelys capellinii and Neochelys salmanticensis), one and a half times larger (as in Neochelys franzeni, Neochelys laurenti, and some specimens of Neochelys arenarum, Neochelys capellinii and Neochelys salmanticensis), but also others in which the posterior margin is two times larger than the anteromedial, this condition being recognized as exclusive for the species. The width of the anal notch about two times less than the maximum width of both xiphiplastra is shared with Neochelys franzeni, Neochelys laurenti, Neochelys liriae, and Neochelys salmanticensis; the condition displayed for other individuals of Neochelys zamorensis, with an anal notch between two or two and a half times less than the maximum width of both xiphiplastra, being similar to that in Neochelys arenarum, Neochelys capellinii, and Neochelys eocaenica. In most of the Neochelys species (i.e., all of them except Neochelys salmanticensis), the relative dimensions of the anal notch fall within a range of two to three times wider than its length. However, in Neochelys zamorensis, a broader variability is evident, with proportions extending from two up to four times wider than its length. Notably, Neochelys salmanticensis exhibits a distinct pattern in the relative dimensions of the anal notch, ranging from four times wider than long (i.e., a characteristic shared with certain specimens of Neochelys zamorensis) to six times wider. This latter condition is exclusive to Neochelys salmanticensis.
Plastral scutes. The gular scute in Neochelys can be slightly wider than each extragular (generally being slightly wider, this condition being recognized in Neochelys capellinii, Neochelys franzeni, Neochelys liriae, and in some specimens of Neochelys eocaenica, Neochelys salmanticensis, and Neochelys zamorensis); the width of these scutes can be subequal (as can be observed in some plastra of Neochelys eocaenica, Neochelys salmanticensis, and Neochelys zamorensis); the extragulars can be the widest, each extragular being slightly wider (in Neochelys arenarum and some individuals of Neochelys laurenti), one and a half times wider (exclusively recognized for some specimens of Neochelys zamorensis), or about two times wider than the gular (a condition known for some specimens of Neochelys laurenti). As described in Pérez-García et al. (2023), the variation in the entoplastral length in relation to the overlapping of the gular is highly variable among the Neochelys representatives. Specifically, the entoplastron can be about two times longer than the overlapping of the gular on this plate (i.e., Neochelys arenarum, Neochelys eocaenica, Neochelys franzeni, and some Neochelys arenarum and some Neochelys salmanticensis specimens), two-and-a-half times (i.e., Neochelys franzeni), three times (i.e., Neochelys capellinii, Neochelys liriae, and some Neochelys arenarum and other Neochelys salmanticensis specimens), four times (i.e., Neochelys capellinii and Neochelys laurenti), and six times (i.e., Neochelys laurenti). This state is identified as highly variable for Neochelys zamorensis, all these cases, but also specimens with hardly any overlap of the gular on this plate, being identified. Neochelys zamorensis presents the same hight range of variation than in Neochelys salmanticensis relative to the overlapping of the extragulars on the entoplastron, which can also have one (as in some specimens of Neochelys arenarum), both (as in some specimens of Neochelys arenarum and Neochelys laurenti), or none (as in Neochelys capellinii, Neochelys eocaenica, Neochelys franzeni, and Neochelys liriae) of these scutes overlapping the plate. Neochelys zamorensis also shares with Neochelys salmanticensis a range of variation relative to the contact of the gular-pectoral scute, higher than in the other members of Neochelys. In this sense, this contact can be absent, as in Neochelys capellinii, Neochelys laurenti, Neochelys liriae, and some individuals of Neochelys arenarum, or, by contrast, can be developed, being short (as in some specimens of Neochelys arenarum and Neochelys eocaenica) or long (as in Neochelys franzeni and some plastra of Neochelys eocaenica).
CONCLUSIONS
Several lineages of turtles are identified in the Eocene levels of the Spanish Duero Basin: Testudinidae, Carettochelyidae, Trionychidae, and Pelomedusidae. The most abundant and diverse are the podocnemidids. Two representatives, recognized as exclusive from that area, are identified: the Bartonian Neochelys salmanticensis (from the Salamanca Province) and the Lutetian Neochelys zamorensis (from the Zamora Province). The shell anatomy Neochelys salmanticensis is well-known, but not that of Neochelys zamorensis. Thus, the presence of this last form has been exclusively confirmed in its type locality: El Escobar fossil site, in the Sanzoles Municipality. In fact, only its holotype has so far been described and figured. However, it does not correspond to an adult individual. Fortunately, several well-preserved partial shells from that locality, and attributable to the same taxon, were found, but all of them remained until now as unpublished. Among these remains are some adult individuals, with a shell size one and a half times larger than that of the holotype. All these remains are analyzed here, so that knowledge about the shell anatomy of this species is notably increased, and an updated diagnosis, considering the other representatives of this diverse European genus, can be proposed. To meet this end, numerous unpublished specimens from the same geological formation (i.e., the Entrala Formation), but from other fossil sites in the Zamora Province (the Valdegallina 1, Valdegallina 2, and Valdegallina 3 fossil sites, in the Valdegallina locality of the Zamora Municipality; the Cubillos 1 and Tardemayas 1 fossil sites, in the Infiernos Hill of the Monfarracinos Municipality; and the Teso de la Mora fossil site, in the Torres del Carrizal Municipality), are studied, a selection of nearly 200 specimens being performed. All of them are identified as attributable to Neochelys zamorensis. This large set of well-preserved specimens, in different ontogenetic stages, also allows us the detailed analysis of the intraspecific variability of Neochelys zamorensis, with a much higher degree of precision than known for any other European pleurodire. The future study of cranial material a priori attributable to this Spanish species, coming from the same fossil levels as the shell remains analyzed here, will allow a more detailed characterization of the taxon.
ACKNOWLEDGMENTS
This research was funded by the Spanish Ministerio de Ciencia e Innovación (research project PID2019-111488RB-I00) and a FPI UNED fellowship. The authors thank M.V. Quiralte and S. Menéndez (MGM) for access to some of the specimens analyzed here; F. Alferez (UCM) and J.C. Gutiérrez-Marco (CSIC-UCM) for the efforts that have allowed this material to become part of a public institution; F. Collantes (Palencia) for the donation of the specimen STUS 17944; and the editor Z. Barkaszi, and anonymous reviewer, and E.A. Cadena for comments and suggestions. The authors highlight the work carried out by E. Jiménez for the establishment of the collection deposited in the STUS.
REFERENCES
Adams, D.C., Collyer, M.L., Kaliontzopoulou, A., and Balken, E. 2021. Geomorph: Software for geometric morphometric analyses. R package version 3.3.2.
https://cran.r-project.org/package=geomorph
Arribas Moreno, A., Polo Díez, V., and Jiménez Fuentes, E. 1984. La “Enfermedad de la Piedra” en la Arenisca de Villamayor. Diagnóstico, tratamiento y conservación. Caja de Ahorros y M.P. Salamanca, Serie Monografías, 2:5–169.
Baur, G. 1888. Osteologische Notizen über Reptilien. Fortsetzung III. Zoologischer Anzeiger, 11:417–424.
Bergounioux, F.M. 1954. Les Ché1oniens fossiles des terrains tertiaires de la Vénétie. Memorie degli Istituti di geologia e mineralogia dell’Università di Padova, 18:1–115.
Böhm, S. 2010. Ecology of chelid turtles Platemys platycephala, Mesoclemmys gibba and Mesoclemmys nasuta in French Guyana. With notes on short term migrations and dietary spectrum of Platemys platycephala in the Nouragues Field Reserve, French Guyana. Unpublished PhD Thesis, Universität Wien, Viena.
Bonaparte, C.L. 1836. Cheloniorum Tabula Analytica. Rome.
Bour, R. 1983. Trois populations endémiques du genre Pelusios (Reptilia, Chelonii, Pelomedusidae) aux îles Seychelles; relations avec les espèces africaines et malgaches. Bulletin du Muséum national d’Histoire naturelle, 5:343–381.
Bour, R., Luiselli, L., Petrozzi, F., Hoinsoudé Segniagbeto, G., and Chirio, L. 2016. Pelusios castaneus (Schweigger 1812) - West African Mud Turtle, Swamp Terrapin. Chelonian Research Monographs, 5:095.1–095.11.
https://doi.org/10.3854/crm.5.095.castaneus.v1.2016
Broadley, D.G. and Boycott, R.C. 2009. Pelusios sinuatus (Smith1838) - Serrated Hinged Terrapin. Chelonian Research Monographs, 5:036.1–036.5.
https://doi.org/0.3854/crm.5.036.sinuatus.v1.2009
Cadena, E.A. 2015. A global phylogeny of Pelomedusoides turtles with new material of Neochelys franzeni Schleich, 1993 (Testudines, Podocnemididae) from the middle Eocene, Messel Pit, of Germany. PeerJ, 3:e1221.
https://doi.org/10.7717/peerj.1221
Cadena, E.A., Jaramillo, C.A., and Bloch, J.I. 2013. New material of the platychelyid turtle Notoemys zapatocaensis from the Early Cretaceous of Colombia; implications for understanding Pleurodira evolution, p. 105–120. In Brinkman, D.B., Holroyd, P.A., and Gardner J.D. (eds.), Morphology and evolution of turtles. Springer, the Netherlands.
https://doi.org/10.1007/978-94-007-4309-0_8
Castaño-Mora, O.V., Cárdenas-Arévalo, G., Gallego-García, N., and Rivera-Díaz, O. 2005. Protección y Conservación de los Quelonios Continentales en el departamento de Córdoba. Convenio No. 28. Universidad Nacional de Colombia, Instituto de Ciencias Naturales, Corporación Autónoma Regional de los Valles del Sinú y San Jorge CVS, Bogotá, Colombia.
Castellano, C.M., Rhodin, A.G.J., Ogle, M., Mittermeier, R.A., Randriamahazo, H., Hudson, R., and Lewis, R.E. 2013. Turtles on the Brink in Madagascar: Proceedings of Two Workshops on the Status, Conservation, and Biology of Malagasy Tortoises and Freshwater Turtles. Chelonian Research Monographs, 6:1–184.
Cope, E.D. 1864. On the limits and relations of the Raniformes. Publications of the Academy of Natural Sciences of Philadelphia, 16:181–183.
Cope, E.D. 1868. On the origin of genera. Publications of the Academy of Natural Sciences of Philadelphia, 20:242–300.
de Broin, F. 1977. Contribution à l'étude des Chéloniens. Chéloniens continentaux du Crétacé et du Tertiaire de France. Mémoires du Muséum National d’Histoire Naturelle, C38:1–366.
de Broin, F. 1988. Les tortues et le Gondwana. Examen des rapports entre le fractionnement du Gondawana et la dispersion géographique des tortues pleurodires à partir du Crétacé. Studia Geologica Salmanticensia, Studia Palaeocheloniologica, 2:103–142.
de Zigno, A. 1889. Chelonii scoperti nei terreni cenozoici delle prealpi Venete. Memorie del Reale Istituto Veneto di Scienze, Lettere ed Arti, 23:119–129.
Djordjevic, S., Bonnet, X., Tomovíc, L., Ajtíc, R., Golubovíc, A., and Djurakíc, M. 2011. Sexual body size and body shape dimorphism of Testudo hermanni in central and eastern Serbia. Amphibia-Reptilia, 32:445–458.
Duméril, A. M.C. and Bibron, G. 1835. Erpétologie Générale ou Histoire Naturelle Complète des Reptiles. Tome second. Librairie Encyclopédique de Roret, Paris.
Ernst, C.H. 1986. Platemys, Platemys platycephala. Catalogue of American Amphibians and Reptiles, 405:1–4.
Fitzinger, L.J. 1826. Neue Classification der Reptilien, nach ihren Natürlichen Verwandtschaften nebst einer Verwandtschafts-Tafel und einem Verzeichnisse der Reptilien-Sammlung des k.k. Zoologischen Museum, Wien.
Fruciano, C. 2016. Measurement error in geometric morphometrics. Development Genes and Evolution, 226:139–158.
https://doi.org/10.1007/s00427-016-0537-4
Gallego-García, N. and Forero-Medina, G. 2014. Plan de manejo para la tortuga de río Podocnemis lewyana en la cuenca del río Sinú. Corporación Autónoma Regional de los Valles del Sinú y del San Jorge, Empresa Urrá S.A. E.S.P., Wildlife Conservation Society, Turtle Survival Alliance y Conservación Internacional, Montería, Colombia.
Garcia, G. 2005. Ecology, human impact and conservation for the Madagascan side-necked turtle (Erymnochelys madagascariensis Grandidier, 1867) at Ankarafantsika National Park, Madagascar. Unpublished PhD Thesis, University of University of Kent, Canterbury, Great Britain.
Gentil, E., De Medeiros, L.A., Vogt, R.C., and Barnett, A.A. 2021. Biology of the Big-headed Amazon River Turtle, Peltocephalus dumerilianus (Schweigger, 1812) (Testudines, Pleurodira): the basal extant Podocnemididae species. Herpetozoa, 34:207–222.
https://doi.org/10.3897/herpetozoa.34.e67807
Gerlach, J. 2008. Pelusios castanoides intergularis Bour 1983 - Seychelles Yellow-Bellied Mud Turtle, Seychelles Chestnut-Bellied Terrapin. Chelonian Research Monographs, 5:010.2–010.4.
https://doi.org/10.3854/crm.5.010.intergularis.v1.2008
González-Zárate, A., Montenegro, O., Castaño-Mora, O.V., and Vargas-Ramírez, M. 2014. Abundancia, estructura poblacional y conservación de Podocnemis lewyana (Podocnemididae) en el río Prado, Colombia. Acta Biológica Colombiana, 19:351–361.
https://doi.org/10.15446/abc.v19n3.41565
Guerrero, A. and Pérez-García, A. 2021a. Analysis of the sexual variability in the shell of the Spanish Upper Cretaceous turtle Dortoka vasconica (Pan-Pleurodira, Dortokidae). Spanish Journal of Palaeontology, 36:149–162.
https://doi.org/10.7203/sjp.36.2.21018
Guerrero, A. and Pérez-García, A., 2021b. Morphological variability and shell characterization of the European uppermost Jurassic to lowermost Cretaceous stem turtle Pleurosternon bullockii (Paracryptodira, Pleurosternidae). Cretaceous Research, 125:104872.
https://doi.org/10.1016/j.cretres.2021.104872
Gunz, P. and Mitteroecker, P. 2013. Semilandmarks: a method for quantifying curves and surfaces. Hystrix, the Italian Journal of Mammalogy, 24:103–109.
https://doi.org/10.4404/hystrix-24.1-6292
Host, B.M. 2006. Husbandry Manual for Eastern Snake Necked Turtle, Chelodina longicollis, Reptilia: Chelidae. Western Sydney Institute of TAFE, Richmond.
Jackson, D. A. 1993. Stopping rules in principal components analysis: a comparison of heuristical and statistical approaches. Ecology, 74:2204–2214.
https://doi.org/10.2307/1939574
Jarek, S. 2012. Mvnormtest: Normality test for multivariate variables. R package version 0.1-9.
Jiménez Fuentes, E. 1968. Stereogenys salmanticensis nov. sp., quelonio eocénico del Valle del Duero. Estudios Geológicos, 24:191–203.
Jiménez Fuentes, E. 1970. Nuevos fragmentos específicos de pelomedúsidos lutecienses del Valle del Duero. Boletín de la Real Sociedad Española de Historia Natural, 68:243–257.
Jiménez Fuentes, E. 1971. Los reptiles fósiles del Valle del Duero: Podocnemis carbajosai, nov. sp., del Eoceno de Salamanca. Estudios Geológicos, 27:85–93.
Jiménez Fuentes, E. 1977. Sinopsis sobre los yacimientos fosilíferos paleógenos de la provincia de Zamora. Boletín Geológico y Minero, 88:357–364.
Jiménez Fuentes, E. 1982. Quelonios y cocodrilos fósiles de la Cuenca del Duero. Ensayo de biozonación del Paleógeno del a Cuenca del Duero. I Ciclo de Conferencias de Paleontología, Salamanca, p.125–127.
Jiménez Fuentes, E. 1986. Síntesis del Paleógeno continental de la Cuenca del Duero, p. 103–108. In Comba, J.A. (coord.), Geología de España II, Libro jubilar J.M. Ríos. Instituto Geológico y Minero de España, Madrid.
Jiménez Fuentes, E. 1992. Quelonios fósiles de Castilla y León, p. 71–100. In Jiménez Fuentes, E. and Civis Llovera, J. (eds.), Vertebrados fósiles de Castilla y León. Junta de Castilla y León, Salamanca.
Jiménez Fuentes, E. 1993. Aclaraciones sobre el status de Neochelys zamorensis, pelomedúsido (Reptilia, Chelonii) de pequeña talla del Eoceno de Zamora (España). Studia Geologica Salmanticensia, 28:141–153.
Jiménez Fuentes, E., Civis, J., González Delgado, A., and Valle, M.F. 1985. Investigaciones salmantinas en Paleontología. Revista Provincial de Estudios, 15:25–34.
Jiménez Fuentes, E., Martín de Jesús, S., and Ortega Coloma, F.J. 1991. Excavaciones paleontológicas en Zamora durante 1989. Anuario Inst. Estudios Zamoranos Florián de Ocampo, 1989:211–215.
Jiménez Fuentes, E., Martín de Jesús, S., Fincias San Martín, B., del Prado Jurado, J., and Mulas Alonso, E. 1992. La Herpetofauna paleógena de la Cuenca del Duero: Distribución cronoestratigráfica y consideraciones paleoecológicas, Acta Zoológica Lilloana, 41:371–380.
Jiménez Fuentes, E. and Gil Tudanca, S. 1993. Vertebrados fósiles de Zamora. Anuario Inst. Estudios Zamoranos Florián de Ocampo, 1993:15–29.
Keswick, T. and Hofmeyr, M.D. 2015. Sexual dimorphism and geographic variation in the morphology of a small southern African tortoise Psammobates oculifer. Amphibia-Reptilia, 36:55–64.
Larson, P.M. 2002. Chondocranial development in larval Rana sylvatica (Anura: Ranidae): morphometric analysis of cranial allometry and ontogenetic shape change. International Journal of Morphology, 252:131–144.
https://doi.org/10.1002/jmor.1095
Limaverde, S., Pêgas, R.V., Damasceno, R., Villa, C., Oliveira, G., Bonde, N., and Leal, M.E.C. 2020. Interpreting character variation in turtles: Araripemys barretoi (Pleurodira: Pelomedusoides) from the Araripe Basin, Early Cretaceous of Northeastern Brazil. PeerJ, 6:e27262v1.
https://doi.org/10.7287/peerj.preprints.27262v1
Mulas, E. and Alonso Gavilán, G. 1987. Facies lacustres de Cubillos (Zamora). Studia Geologica Salmanticensia, 24:10–14.
Mulas Alonso, E., Martín de Jesús, S., and Jiménez Fuentes, E. 1988. Sobre la herpetofauna pa1eógena de algunos yacimientos al N. del Duero (Zamora). Studia Geologica Salmanticensia, 25:181–185.
Ortega, F., Armenteros, I., de Celis, A., Escaso, F., Huerta, P., Martín de Jesús, S., Narváez, I., Pérez-García, A., and Sanz, J.L. 2022. Crocodyliformes and Testudines from the Eocene of the Duero Basin (northwestern Spain): an update of their diversity and stratigraphic context. Historical Biology, 34:1560–1581.
https://doi.org/10.1080/08912963.2022.2051503
Páez, V.P., Restrepo, A., Vargas Ramírez, M., Bock, B.C., and Gallego García, N. 2013. Podocnemis lewyana (Duméril 1852). Tortuga del río Magdalena, tortuga de río. Asociación Colombiana de Herpetología, 1:1–5.
Parra-Henao, K.D., Páez, V.P., Morales-Betancourt, M., and Lasso, C.A. 2019. A pilot study of habitat use and population characteristics of the Big-headed Amazon River Turtle, Peltocephalus dumerilianus (Schweigger, 1812), during the dry season in the Bojonawi Nature Reserve, Vichada, Colombia. Herpetology Notes, 12:1113–1120.
Pérez-García, A. 2015. Nuevos datos sobre la tortuga pleurodira del Eoceno medio de Alemania Neochelys franzeni Schleich 1993: implicaciones derivadas del hallazgo de un ejemplar juvenil. Geogaceta, 57:71–74.
Pérez-García, A. 2017. The Iberian fossil record of turtles: an update. Journal of Iberian Geology, 43:155–191.
https://doi.org/10.1007/s41513-017-0016-4
Pérez-García, A. and de Lapparent de Broin, F. 2013. A new species of Neochelys (Chelonii, Podocnemididae) from the Ypresian (early Eocene) of the south of France. Comptes Rendus Palevol, 12:269–277.
https://doi.org/10.1016/j.crpv.2013.05.011
Pérez-García, A. and de Lapparent de Broin, F. 2015. New insights into the anatomy and systematic of ‘Papoulemys’ laurenti, a representative of Neochelys (Chelonii, Podocnemididae) from the early Eocene of the south of France. Paläontologische Zeitschrift, 89:901–923.
https://doi.org/10.1007/s12542-015-0259-3
Pérez-García, A., Guerrero, A., Martín de Jesús, S., and Ortega, F. 2023. Shell characterization of the youngest valid species of the European Eocene genus Neochelys (Pleurodira, Podocnemididae): The Spanish Bartonian Neochelys salmanticensis. The Anatomical Record, 2023:1–20.
https://doi.org/10.1002/ar.25225
Pritchard, P.C.H. 1979. Encyclopedia of turtles. TFH Publications, Neptune.
R Core Team. 2020. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
Rohlf, F.J. 2010. tpsDig v. 2.16. State University of New York at Stony Brook, New York.
Rohlf, F.J. 2015. The tps series of software. Hystrix, the Italian Journal of Mammalogy, 26:9–12.
https://doi.org/10.4404/hystrix-26.1-11264
Rohlf, F.J. and Slice, D. 1990. Extensions of the Procrustes method for the optimal superimposition of landmarks. Systematic Zoology, 39:40–59.
https://doi.org/10.2307/2992207
Rueda-Almonacid, J.V., Mittermeier, C., Carr, J.L., Mittermeier, R.A., Malecha, J.V.R., Mast, R., Vogt, R.C., Rhodin, A.G.J., de la Ossa-Velásquez, J., Rueda, J.N., and Mittermeier, C.G. 2007. Las tortugas y los cocodrilianos de los países andinos del trópico. Serie de guías tropicales de campo Nº 6. Conservación Internacional. Editorial Panamericana, Formas e Impresos. Bogotá, Colombia.
Sepúlveda-Seguro, A.M., Páez, V.P., Gómez-Rincón, M.T., Morales-Betancourt, M.A., and Lasso, C.A. 2020. Evaluación de las características de las poblaciones y el uso de los hábitats de Podocnemis vogli (Müller 1935) durante una temporada reproductiva en la reserva natural privada de Bojonawi (Departamento de Vichada, Colombia). Revista Latinoamericana de Herpetología, 3:83–94.
https://doi.org/10.22201/fc.25942158e.2020.2.139
Strydom, A.V. 2001. Seasonal Reproduction and Sexual Size Dimorphism of the African Helmeted Turtle, Pelomedusa subrufa (Family Pelomedusidae). Unpublished PhD Thesis, University of Stellenbosch, South Africa.
Trembath, D. and Freier, D. 2004. Sexual-size dimorphism in Emydura krefftii (Testudines: chelidae) from Ross River, Townsville, Australia. Herpetological Review, 35:31–34.
Vetter, H. 2004. Terralog: Turtles of the World Vol. 2 - North America. Edition Chimaira, Frankfurt am Main, and Verlag ACS GmbH, Rodgau.
Vilanova, J. 1873. Noticias de vertebrados hallados en Sanzoles (Zamora). Acta de la Sociedad Española de Historia Natural, 2:41–52.
Wagler, J.G. 1830. Natürliches System der Amphibien, mit vorangehender Classification der Säugthiere und Vögel. Ein Beitrag zur vergleichenden Zoologie. J. G. Cotta'schen Buchhandlung, München.
Zelditch, M.L., Swiderski, D.L., and Sheets, H.D. 2012. A Practical Companion to Geometric Morphometrics for Biologists: Running analyses in freely-available software. Academic Press, USA.
