

 Revision of the Early Devonian psammosteids from the “Placoderm Sandstone”: Implications for their body shape reconstruction
Revision of the Early Devonian psammosteids from the “Placoderm Sandstone”: Implications for their body shape reconstruction
Article number: 22.2.36
https://doi.org/10.26879/948
Copyright Paleontological Society, June 2019
Author biography
Plain-language and multi-lingual abstracts
PDF version
Submission: 29 November 2018. Acceptance: 19 May 2019
{flike id=2593}
ABSTRACT
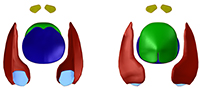
The Early Devonian psammosteid agnathans (Guerichosteus and Hariosteus) originally described by Tarlo (1964, 1965) from the so-called “Placoderm Sandstone” are revised. The newly collected material from the Holy Cross Mountains allows detailed reexamination and provides new anatomical information. The morphology and ornamentation of the plates supports the establishment of the genera Guerichosteus and Hariosteus. However, G. kotanskii, G. kulczyckii and G. lefeldi should be considered as junior synonyms of Guerichosteus kozlowskii. Hariosteus lobanowskii, erected by Tarlo (1964), is considered here as invalid (nomen dubium) and H. kotanskii is included with Hariosteus kielanae. The three-dimensional scanning of plates belonging to Guerichosteus allows for a more detailed description and reconstruction. In cross section the deep ventral part of the trunk is V-shaped in contrast to previous U-shaped restorations. Probably, the Early Devonian psammosteid, Drepanaspis gemuendenesis in particular, shared this body shape.
Marek Dec. Institute of Palaeobiology, Polish Academy of Sciences, Twarda 51/55, 00-818 Warszawa, Poland. mdec@wp.pl
Keywords: agnatha; Heterostraci; three-dimensional reconstruction
Dec, Marek. 2019. Revision of the Early Devonian psammosteids from the "Placoderm Sandstone": Implications for their body shape reconstruction. Palaeontologia Electronica 22.2.36A 1-26. https://doi.org/10.26879/948
palaeo-electronica.org/content/2019/2593-psammosteids-revision
Copyright: June 2019 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
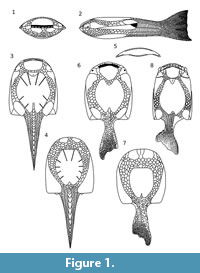 Psammosteids are an extinct group of armored agnathans occurring exclusively in the Devonian. Among these fishes, the Early Devonian Drepanaspis is best recognized. It is the only psammosteid with a nearly completely preserved exoskeleton composed of the entire armored body surface, trunk and tail (Janvier, 1996). First reconstructions of Drepanaspis (Traquair, 1899, 1903) showed the dorsal and ventral surface incorrectly, as noted by Dean (1904). This was later confirmed by Kiaer (1915) and most subsequent authors (Stensiö, 1927; Kiaer, 1928; Stetson, 1931; Obruchev, 1943; Gross, 1963). The most detailed description together with reconstructions of Drepanaspis gemuendenesis were made by Gross (1963), although many other authors also provided some reconstructions (Stenson, 1931; Obruchev, 1943; Tarlo, 1961; Tarlo, 1964; Obruchev and Mark-Kurik, 1965) (Figure 1).
Psammosteids are an extinct group of armored agnathans occurring exclusively in the Devonian. Among these fishes, the Early Devonian Drepanaspis is best recognized. It is the only psammosteid with a nearly completely preserved exoskeleton composed of the entire armored body surface, trunk and tail (Janvier, 1996). First reconstructions of Drepanaspis (Traquair, 1899, 1903) showed the dorsal and ventral surface incorrectly, as noted by Dean (1904). This was later confirmed by Kiaer (1915) and most subsequent authors (Stensiö, 1927; Kiaer, 1928; Stetson, 1931; Obruchev, 1943; Gross, 1963). The most detailed description together with reconstructions of Drepanaspis gemuendenesis were made by Gross (1963), although many other authors also provided some reconstructions (Stenson, 1931; Obruchev, 1943; Tarlo, 1961; Tarlo, 1964; Obruchev and Mark-Kurik, 1965) (Figure 1).
The large collection of psammosteids from the Lower Devonian of “Placoderm Sandstone” was described by Tarlo (1964, 1965). This author described two new genera, Guerichosteus containing four new species and Hariosteus containing two species. The material, represented by numerous fragments of plates, is preserved as natural molds in fine-grained sandstone. Many elements of the armor are perfectly preserved and show the most delicate ornamentation, which can be examined by using latex or silicone casts. Despite the abundance of Guerichosteus kozlowskii, no restoration of the species has been made until now. New material obtained during field work with Piotr Szrek and Grzegorz Niedźwiedzki at Podłazie Hill outcrop, conducted in 2011-2013 (Szrek et al., 2014), allows for a revision of the psammosteids formerly described by Tarlo (1957, 1961, 1964, 1965).
The described material is stored at the Institute of the Paleobiology of the Polish Academy of Sciences (abbreviation: ZPAL Ag. III/4), the Museum of the Earth of the Polish Academy of Sciences in Warsaw (abbreviation: MZ-VIII/Vp-430) and the Polish Geological Institute - National Research Institute (abbreviation: MUZ.PIG.1733.II.353). All current specimen numbers with reference to the numbers from the Tarlo (1964, 1965) are presented in Table 1.
MATERIAL AND GEOLOGICAL SETTING
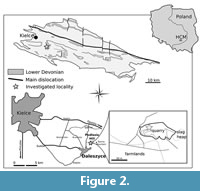 The outcrop of Podłazie Hill is located about 2 km to the north of the Daleszyce, in the southern part of the Holy Cross Mountains (Figure 2.1). The quarry is located on the southern slope of the hill (Figure 2.2). The succession dominated by thick-bedded, quartzitic sandstone is considered to be of Emsian age (Tarnowska, 1976, 1981; Szulczewski and Porębski, 2008), based on miospores and tephrocorrelation (Fijałkowska-Mader and Malec, 2011).
The outcrop of Podłazie Hill is located about 2 km to the north of the Daleszyce, in the southern part of the Holy Cross Mountains (Figure 2.1). The quarry is located on the southern slope of the hill (Figure 2.2). The succession dominated by thick-bedded, quartzitic sandstone is considered to be of Emsian age (Tarnowska, 1976, 1981; Szulczewski and Porębski, 2008), based on miospores and tephrocorrelation (Fijałkowska-Mader and Malec, 2011).
The siliciclastic, storm-originated, bone-bearing beds yield a rich vertebrate accumulation deposited in a marginal-marine (e.g., lagoon) environment. The skeletons of fish were broken and sorted, likely as a result of their redeposition in shallow water. Ichnofossils show the presence of the Cruziana ichnofacies in the section (Szrek et al., 2014).
The material from the quartzitic sandstone bone-beds consists of natural molds of since dissolved fish remains, mostly secondarily filled with clay. Therefore, bony tissues are not preserved, although the imprints reveal minute structural details, such as those of histological aspects (e.g., the boundary between parts previously occupied by cartilage and membranous bones; Tarlo, 1964). In some cases, bone tissue is preserved, but the apatite has been replaced by plumbogummite-group minerals (Kruszewski and Dec, 2018) without alteration of bone histological structures.
The specimens preserved in quartzitic sandstone filled with silt had to be mechanically cleaned and brushed in hot water with detergent. After removing silt from cavities with bone imprints, the latex casts (colored in grey) were made with Gumosill WW.
Most material is composed of broken plates, whereas any smaller plates or scales are preserved completely (Szrek et al., 2014). Most specimens lack deformation and the latex casts reflect the shape of actual bone or plate.
METHOD OF PSAMMOSTEIDS MORPHOLOGY RECONSTRUCTION
Computer methods in paleontology have developed substantially in the last decade, especially three-dimensional (3D) digitization of specimens and models achieved by different processes. Photogrammetry was a basic method used herein to produce the 3D models of specimens presented. To achieve a good resolution and models with a good accuracy (Falkingham, 2012), specific steps were followed (Mallison and Wings, 2014):
1. Photography;
2. Model calculation;
3. Polygon and texture mesh generation; and
4. Data export.
The photography is the important part of the process and has significant influence on the final effect. Therefore, the high resolution Digital Single Lens Reflex Camera (DSLR), Canon 70D (20.2-megapixels), was used. To visualize most specimens, the Sigma DC 17-50 mm 1:2.8 EX HSM lens was used (usually a 50 mm focal length and an f/10 aperture were used). For small (about 1-3 cm) specimens, a Canon EF-S 55-250 mm 1:4-5.6 IS with macro lens +4D was used (usually a 105 mm focal length and an f/10 aperture were used).
The photographs were taken with the camera mounted on a tripod and specimen placed on rotating stage at the desired angle. To generate a mesh, photographs were taken from at least two levels, one at a low angle 10-20 degree) and the second at a high level (40-60 degree). Model creation involved a point cloud calculation in VirtualSFM, polygon and texture mesh generation in MeshLab 2016.12, and exporting object mesh to Blender 2.79 (3D modeling software) to put together the plates.
SYSTEMATIC PALEONTOLOGY
Subclass Heterostraci Lankester, 1868
Order Pteraspidiformes Berg, 1940
Family Psammosteidae Traquair, 1896
Remarks. Psammosteids are considered to be closely related to Pteraspidiformes. Different phylogenetic reconstructions recognized them either as a derived (Janvier, 1996), basal or sister (Halstead, 1973; Blieck, 1984; Pernègre and Elliott, 2008) group of Pteraspidiformes. While the precise clade position is still ambiguous, the pteraspidiform affinity seems to be uncontested (Randle and Sansom, 2017a). Therefore, at present psammosteids are considered to be the family Psammosteidae (Janvier, 1996; Pernègre and Elliott, 2008; Randle and Sansom, 2017a, 2017b), which is a position adopted herein, although currently some authors nest them in suborder Psammosteida within the order Pteraspidiformes (Elliott, Mark-Kurik, and Daeschler, 2004; Glinskiy and Mark-Kurik, 2016; Glinskiy and Nilov, 2017).
Genus GUERICHOSTEUS Tarlo, 1964
Type species. Guerichosteus kozlowskii Tarlo, 1964, by original designation.
Guerichosteus kozlowskii
Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9, FIgure 10, Figure 11, Appendix 1, Appendix 2, Appendix 3, Appendix 4, Appendix 5, Appendix 6, Appendix 7, Appendix 8, Appendix 9, Appendix 10, Appendix 11, Appendix 12, Appendix 13, Appendix 14, Appendix 15, Appendix 16, Appendix 17, Appendix 18, Appendix 19, Appendix 20, Appendix 21
See Reconstruction
v. 1957. Psammosteid type A; Tarlo 1957, p. 230, pl. 1, fig. 1.
. 1965 Guerichosteus kozlowskii; Tarlo 1965: pp. 40-50, text-figs. 5B-C, pl. X, figs. 2, 5, 7; pl. XI, figs. 6, 8-9; pl. XII, fig. 2.
v. 1965 Guerichosteus kozlowskii; Tarlo 1965: pp. 40-50, text-fig. 4, 5A, 6-8, pl. IX figs. 1A-B, 4, 5, 6; pl. X, figs. 1, 3-4, 6, 8-9; pl. XI, figs. 2-4, 7, 10-13; pl. XII figs. 1, 3-4.
. 1965 Guerichosteus kotanskii; Tarlo 1965: pp. 50, pl. XII, figs. 5-6.
v. 1965 Guerichosteus kotanskii; Tarlo 1965: pp. 50, pl. XII, fig. 7.
. 1965 Guerichosteus kulczyckii; Tarlo 1965: pp. 50-51, pl. VII, figs. 3-4, XI, fig. 15.
v. 1965 Guerichosteus lefeldi; Tarlo 1965: pp. 51-52, pl. XII, fig. 9.
. 1965 Guerichosteus lefeldi; Tarlo 1965: pp. 51-52, pl. XII, fig. 8.
Diagnosis. Ventral median plate deep, rectangular in outline with posterior notch developed as narrow and shallow longitudinal depression reaching the center of plate instead of slit. Dorsal median plate roughly circular in outline with anterior re-entrant angle present. Branchial plate long and fairly narrow. Cornual plate rectangular. Ornamentation consists of closely-packed, rounded crenulated tubercles varying in size from 7 to 16 per mm (emended after Tarlo, 1965).
Description
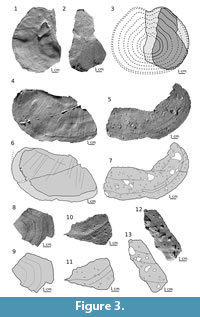 Dorsal plate. The specimen ZPAL Ag. III/4 (Figure 3.1-3; Appendix 1) described by Tarlo (1965, fig. 5A; pl. XII, fig. 3-4) as a ventral median plate is the right part of the dorsal plate. The internal side (10 cm long and 7.7 cm wide) shows an anterior margin with re-entrant angle, lateral and half of posterior margins. Along the edge an area of 2-7 mm extends without a basal layer. The anterior portion of the plate is flat while bending posteriad to the growth center creating a slight convexity (Appendix 1 in supplementary material). The anterior half of the external surface shows the ornamentation with a distinct growth lines. The plate is subcircular in outline and gets slightly constricted anteriad at the re-entrant angle level. The large posterior part (9 cm long, 13.8 cm width) of median plate ZPAL Ag. III/1 (Figure 3.4-7; Appendix 2; Tarlo, 1965, pl. X, fig. 9) shows both sides, external (Figure 3.5, 3.7) and internal (Figure 3.4, 3.6; note that Tarlo (1965) showed only the external ornamented side of the specimen), with a natural posterior and lateral margin preserved. Posterior edge, fairly straight, undulates and curves around to the lateral margin. Well-marked growth lines extend parallel to the margin. In specimens ZPAL Ag. III/4 and ZPAL Ag. III/1 there is lack of the basal layer near the edge, but in the latter specimen this area is much wider (10-18 mm).
Dorsal plate. The specimen ZPAL Ag. III/4 (Figure 3.1-3; Appendix 1) described by Tarlo (1965, fig. 5A; pl. XII, fig. 3-4) as a ventral median plate is the right part of the dorsal plate. The internal side (10 cm long and 7.7 cm wide) shows an anterior margin with re-entrant angle, lateral and half of posterior margins. Along the edge an area of 2-7 mm extends without a basal layer. The anterior portion of the plate is flat while bending posteriad to the growth center creating a slight convexity (Appendix 1 in supplementary material). The anterior half of the external surface shows the ornamentation with a distinct growth lines. The plate is subcircular in outline and gets slightly constricted anteriad at the re-entrant angle level. The large posterior part (9 cm long, 13.8 cm width) of median plate ZPAL Ag. III/1 (Figure 3.4-7; Appendix 2; Tarlo, 1965, pl. X, fig. 9) shows both sides, external (Figure 3.5, 3.7) and internal (Figure 3.4, 3.6; note that Tarlo (1965) showed only the external ornamented side of the specimen), with a natural posterior and lateral margin preserved. Posterior edge, fairly straight, undulates and curves around to the lateral margin. Well-marked growth lines extend parallel to the margin. In specimens ZPAL Ag. III/4 and ZPAL Ag. III/1 there is lack of the basal layer near the edge, but in the latter specimen this area is much wider (10-18 mm).
Several fragments of flat plates such as ZPAL Ag. III/2 (Figure 3.8-9; Appendix 3; Tarlo 1965, pl. X, fig. 1), MUZ.PIG.1733.II.13 (Figure 3.10-11; Appendix 4) and MZ-VIII/Vp-438 (Figure 3.12-13; Appendix 5), appear to be parts of a dorsal plate. ZPAL Ag. III/2 shows growth line patterns similar to ZPAL Ag. III/4. If so, it may indicate ZPAL Ag. III/2 is a fragment of the right side of the plate. In the latter specimen the tubercles are well preserved, and short radial grooves are visible on the surface (Figure 3.8). They were probably a part of the sensory canal system (Tarlo, 1965).
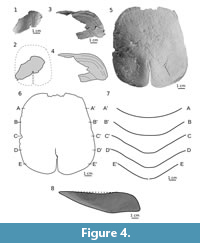 Ventral plate. The specimen MUZ.PIG.1733.II.353 (Figure 4.3-8; Appendix 6) is the largest and most complete plate. However, only the internal side (Figure 4.5-6) displays the whole surface of the plate. The external, ornamented surface is exposed only in the anterior part of the plate (Figure 4.3-4). The plate is rectangular in shape with a length of 118 mm and a width of 103 mm (length-to-width ratio is 1.15). The plate is 20 mm deep and trapezoidal in transverse section along the posterior notch (here developed as a depression instead of a slit), while anteriorly it is more rounded (Figure 4.6-7). The anterior margin is straight with a very shallow concavity in the middle. The posterior margin is smoothly rounded and arched to the medially located posterior notch. The margins of the slit are irregular, which suggests a crack more than a natural edge. On the internal side along the slit there is a slight thickening visible, so the posterior notch is developed as a low depression rather than a slit like in Tartuosteus or Schizosteus.
Ventral plate. The specimen MUZ.PIG.1733.II.353 (Figure 4.3-8; Appendix 6) is the largest and most complete plate. However, only the internal side (Figure 4.5-6) displays the whole surface of the plate. The external, ornamented surface is exposed only in the anterior part of the plate (Figure 4.3-4). The plate is rectangular in shape with a length of 118 mm and a width of 103 mm (length-to-width ratio is 1.15). The plate is 20 mm deep and trapezoidal in transverse section along the posterior notch (here developed as a depression instead of a slit), while anteriorly it is more rounded (Figure 4.6-7). The anterior margin is straight with a very shallow concavity in the middle. The posterior margin is smoothly rounded and arched to the medially located posterior notch. The margins of the slit are irregular, which suggests a crack more than a natural edge. On the internal side along the slit there is a slight thickening visible, so the posterior notch is developed as a low depression rather than a slit like in Tartuosteus or Schizosteus.
ZPAL Ag. III/45 is fragment of a ventral plate of a juvenile specimen (Figure 4.1-2; Appendix 7; Tarlo 1965, pl. XII, fig 9). The plate is deeply arched and resembles the anterior portion of large specimen MUZ.PIG.1733.II.353 in shape. Near the posterior broken edge a low depression marks the posterior notch (Figure 4.2).
 Branchial plate. Most specimens of branchial plates are broken or incomplete. The most complete is a small, triangular, left branchial plate, ZPAL Ag. III/1089 (Figure 5.1-2; Appendix 8), of a juvenile individual. The plate shows only a dorsal view of the anterior half of the external, ornamented, dorsal surface and the internal side of the ventral surface. The plate is 40 mm long (measured along the median margin of dorsal ornamented surface) and its maximum width is 28 mm (measured perpendicularly to the length). The dorsal ornamented surface is narrow with a straight medial margin except for the tapering of the proximal tip. The medial margin of the ventral surface is concave. The angle between the dorsal and ventral parts is about 65 degrees. The angle and shape of the last specimen resemble those of small fragments of MUZ.PIG.1733.II.185 (Figure 5.7-10) and MZ-VIII/Vp-533 (Figure 5.13-14) as well as the posterior half of the larger ZPAL Ag. III/16 (Figure 5.3-4; Appendix 9), which is 62 mm long. Although in ZPAL Ag. III/16 the anterior half is elongated and narrow, the ventral surface’s maximal width is 18 mm, and the dorsal surface’s maximal width is 12 mm. Therefore, the lateral margin and the median margin of the ventral and dorsal surface run parallel. Also the proximal part of the specimen is less angled (40 degrees). Various fragments of larger specimens showed on Figure 5 and Figure 6 (ZPAL Ag. III/7, ZPAL Ag. III/8. ZPAL Ag. III/9, ZPAL Ag. III/15, MZ-VIII/Vp-423, MUZ.PIG.1733.II.185) are incomplete. Nevertheless, all features of the plate possibly show the mid-part of the plate. Their shape (i.e., narrow anterior part and considerably expanded posterior part) is in general similar to ZPAL Ag. III/16. However, the larger ZPAL Ag. III/15 specimen (Figure 5.5-6; Appendix 10) of the right branchial plate displays a dorsal view of a distal tip, where the medial margin of the dorsal surface is slightly curved.
Branchial plate. Most specimens of branchial plates are broken or incomplete. The most complete is a small, triangular, left branchial plate, ZPAL Ag. III/1089 (Figure 5.1-2; Appendix 8), of a juvenile individual. The plate shows only a dorsal view of the anterior half of the external, ornamented, dorsal surface and the internal side of the ventral surface. The plate is 40 mm long (measured along the median margin of dorsal ornamented surface) and its maximum width is 28 mm (measured perpendicularly to the length). The dorsal ornamented surface is narrow with a straight medial margin except for the tapering of the proximal tip. The medial margin of the ventral surface is concave. The angle between the dorsal and ventral parts is about 65 degrees. The angle and shape of the last specimen resemble those of small fragments of MUZ.PIG.1733.II.185 (Figure 5.7-10) and MZ-VIII/Vp-533 (Figure 5.13-14) as well as the posterior half of the larger ZPAL Ag. III/16 (Figure 5.3-4; Appendix 9), which is 62 mm long. Although in ZPAL Ag. III/16 the anterior half is elongated and narrow, the ventral surface’s maximal width is 18 mm, and the dorsal surface’s maximal width is 12 mm. Therefore, the lateral margin and the median margin of the ventral and dorsal surface run parallel. Also the proximal part of the specimen is less angled (40 degrees). Various fragments of larger specimens showed on Figure 5 and Figure 6 (ZPAL Ag. III/7, ZPAL Ag. III/8. ZPAL Ag. III/9, ZPAL Ag. III/15, MZ-VIII/Vp-423, MUZ.PIG.1733.II.185) are incomplete. Nevertheless, all features of the plate possibly show the mid-part of the plate. Their shape (i.e., narrow anterior part and considerably expanded posterior part) is in general similar to ZPAL Ag. III/16. However, the larger ZPAL Ag. III/15 specimen (Figure 5.5-6; Appendix 10) of the right branchial plate displays a dorsal view of a distal tip, where the medial margin of the dorsal surface is slightly curved. 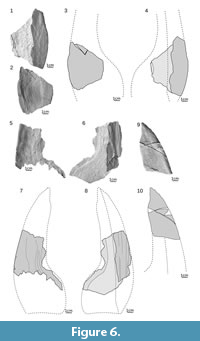 The margin tapers with its posterior corner creating the edge of a branchial opening. In ZPAL Ag. III/9 (Figure 6.5-8; Appendix 13), probably representing the biggest individual, and in ZPAL Ag. III/7 (holotype) (Figure 6.1-4; Appendix 12) the ventral surface is concave. Although the holotype is slightly crushed, the damage occurred probably post-mortem. The angle between the dorsal and ventral surface is about 35-40 degrees in ZPAL Ag. III/9 and ZPAL Ag. III/7. More than half of the space between those surfaces is filled with bone (about 15 mm in ZPAL Ag. III/7) so the plate is quite thick. The proximal tip of the left branchial plate ZPAL Ag. III/14 (Figure 6.9-10; Appendix 14) shows an undulating medial margin in ventral view. The lateral margin is arcuate near the anterior tip and almost straight posteriorly. The plate is 98 mm long and expands to the rear up to 40 mm.
The margin tapers with its posterior corner creating the edge of a branchial opening. In ZPAL Ag. III/9 (Figure 6.5-8; Appendix 13), probably representing the biggest individual, and in ZPAL Ag. III/7 (holotype) (Figure 6.1-4; Appendix 12) the ventral surface is concave. Although the holotype is slightly crushed, the damage occurred probably post-mortem. The angle between the dorsal and ventral surface is about 35-40 degrees in ZPAL Ag. III/9 and ZPAL Ag. III/7. More than half of the space between those surfaces is filled with bone (about 15 mm in ZPAL Ag. III/7) so the plate is quite thick. The proximal tip of the left branchial plate ZPAL Ag. III/14 (Figure 6.9-10; Appendix 14) shows an undulating medial margin in ventral view. The lateral margin is arcuate near the anterior tip and almost straight posteriorly. The plate is 98 mm long and expands to the rear up to 40 mm.
Cornual plate. The almost complete cornual plate ZPAL Ag. III/19 (Figure 7.1-2; Appendix 15) is rectangular and slightly convex. The specimen is 55 mm long and 53 mm wide (length to width ratio is 1.04). The postero-lateral corner of the plate is rounded posteriorly (also the area bent inside) and forms a thickening. The growth center is at the posterior margin of the plate and is displaced laterally (Figure 7.1).
Orbital plate. The trapezoidal right orbital plate is 13 mm long and 25 mm width (length-to-width ratio is 0.52). A round opening for the eye with a diameter of 3.5 mm is in the middle of the inclined lateral margin (Figure 7.3-4; Appendix 16).
Complex plate. One, almost pentagonal, left complex plate MUZ.PIG.1733.II.357 (Figure 7.8-9; Appendix 17) is 42 mm wide and 30 mm long. In transverse (sagittal) section the S-shaped curvature of the plate is visible (Figure 7.9). The anterior margin is sinusoid with the medial part convex forming a space for oral plates. The tubercles on the most prominent part of the plate are weathered.
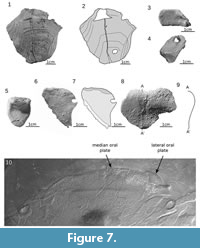 Oral plate. An almost complete and triangular median oral plate is 30 mm long and 26 mm wide (specimen, Figure 7.5; Appendix 18). In transverse section the plate has a slightly S-shaped curvature similar to a complex plate. Beside the median oral plate described by Tarlo (1965; fig. 7A-C), a newly found plate appears to be a lateral oral plate (Figure 7.6-7; Appendix 19). This nearly complete and triangular specimen is 30 mm long and 26 mm wide (Figure 7.6; Appendix 19). In transverse section the plate has a slightly S-shaped curvature similar to a complex plate. The internal side of the plate shows a narrow area with tubercles along a straight anterior margin. The internal margin of the ornamented area is sinusoidal. The anterior edge smoothly turns rearward at an almost right angle forming a concave edge. The third medial edge is not preserved, although on the basis of the overlapping area (Figure 7.7, light gray) the boundary of the plate can be stated. The external side, visible only near the edges of the plate, shows closely packed and small dome-shaped tubercles. Those two oral plates, medial and lateral, look similar to those of Drepanaspis gemuendenesis presented by Tarlo in 1965 (pl VI, figure 1) and to a specimen stored in the National Museum of Natural History in Paris (Figure 7.10) showed by Lehman (1967; planche 1).
Oral plate. An almost complete and triangular median oral plate is 30 mm long and 26 mm wide (specimen, Figure 7.5; Appendix 18). In transverse section the plate has a slightly S-shaped curvature similar to a complex plate. Beside the median oral plate described by Tarlo (1965; fig. 7A-C), a newly found plate appears to be a lateral oral plate (Figure 7.6-7; Appendix 19). This nearly complete and triangular specimen is 30 mm long and 26 mm wide (Figure 7.6; Appendix 19). In transverse section the plate has a slightly S-shaped curvature similar to a complex plate. The internal side of the plate shows a narrow area with tubercles along a straight anterior margin. The internal margin of the ornamented area is sinusoidal. The anterior edge smoothly turns rearward at an almost right angle forming a concave edge. The third medial edge is not preserved, although on the basis of the overlapping area (Figure 7.7, light gray) the boundary of the plate can be stated. The external side, visible only near the edges of the plate, shows closely packed and small dome-shaped tubercles. Those two oral plates, medial and lateral, look similar to those of Drepanaspis gemuendenesis presented by Tarlo in 1965 (pl VI, figure 1) and to a specimen stored in the National Museum of Natural History in Paris (Figure 7.10) showed by Lehman (1967; planche 1).
 Ridge scales. A few incomplete ridge scales (Figure 8) vary in length from 15 mm (MUZ.PIG.1733.II.13, Figure 7.10-11) to 69 mm (ZPAL Ag. III/27A, Figure 7.1-2; Appendix 20). The maximal width is from 8 mm (MUZ.PIG.1733.II.13) to 24 mm (ZPAL Ag. III/27A and MZ-VIII/Vp-463). Most specimens are long and slender apart from MZ-VIII-Vp-463 (Figure 7.14-15), which is relatively wider and shorter. Ridge scales are roundly curved dorsally in the small specimens (ZPAL Ag. III/26, Figure 7.7-9) and became more triangular in the larger specimens (MZ-VIII/Vp-335, Figure 7.12-13 and ZPAL Ag. III/27A, Figure 7.3). The maximal high of the ridge scale is 9 mm, and the maximal thickness is 4 mm (MZ-VIII/Vp-335). The tubercles cover the entire external side and the visceral surface, without its proximal unornamented part (light gray in Figure 7.2, 7.5, 7.15) that was in contact with body tissues.
Ridge scales. A few incomplete ridge scales (Figure 8) vary in length from 15 mm (MUZ.PIG.1733.II.13, Figure 7.10-11) to 69 mm (ZPAL Ag. III/27A, Figure 7.1-2; Appendix 20). The maximal width is from 8 mm (MUZ.PIG.1733.II.13) to 24 mm (ZPAL Ag. III/27A and MZ-VIII/Vp-463). Most specimens are long and slender apart from MZ-VIII-Vp-463 (Figure 7.14-15), which is relatively wider and shorter. Ridge scales are roundly curved dorsally in the small specimens (ZPAL Ag. III/26, Figure 7.7-9) and became more triangular in the larger specimens (MZ-VIII/Vp-335, Figure 7.12-13 and ZPAL Ag. III/27A, Figure 7.3). The maximal high of the ridge scale is 9 mm, and the maximal thickness is 4 mm (MZ-VIII/Vp-335). The tubercles cover the entire external side and the visceral surface, without its proximal unornamented part (light gray in Figure 7.2, 7.5, 7.15) that was in contact with body tissues.
Trunk scales. In the studied material several trunk scales are recognized. The largest scale, ZPAL Ag. III/27B, is 28 mm long and 17 mm wide and shows a narrow (3 mm) overlapping area in the proximal margin (Figure 8.16-17). ZPAL Ag. III/33 resembles the latter specimen, but is poorly preserved. Its length is 23 mm and width is 10 mm (Figure 8.18). This specimen also has a row of longitudinal tubercles arranged perpendicularly to the margin of the scale. ZPAL Ag. III/28 and ZPAL Ag. III/33 represent more proximal trunk scales. A more distal scale is ZPAL Ag. III/22, which is 25 mm long and 17 mm wide (Figure 8.19; Appendix 21). The central part of the scale is elevated (about 1 mm). The last scale was probably placed in the tail area. A small and rhomboidal scale MZ-VIII/Vp-536 is 17 mm long and 13 mm wide (Figure 8.20). In contrast to ZPAL Ag. III/27B and ZPAL Ag. III/33, which are flat and thin, this scale is convex and thick.
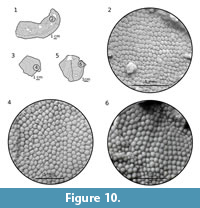
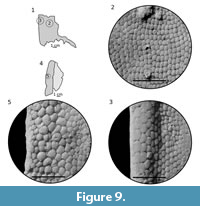 Ornamentation. The ornamentation consists of closely packed, rounded, irregular to pentagonal tubercles (Figure 9, Figure 10). All tubercles have well-marked crenulation at the base. On branchial plates the largest tubercles (7-8 per mm) are located along the lateral margin (Figure 9.3, 9.5) and most of them are low and pentagonal in shape. Toward the medial part of the plate, the size of tubercles decreases from 10-12 per mm (Figure 9.3) to 15-16 per mm (Figure 9.2). The tubercles are mostly low, almost flat and irregular to pentagonal in shape. Nevertheless, in some parts the tubercles may be higher and more dome-shaped (Figure 9.5). Lower, more closely packed and less varied are tubercles on the dorsal (Figure 10) and ventral plates whose shape is pentagonal or irregular. The difference is in the size of tubercles; on the dorsal plate tubercles are larger (12-14 per cm; Figure 10.2, 10.4) than those on the ventral plate (18-20 per cm). The cornual plate possesses dome-shaped tubercles (9-13 per cm) with a well-visible base crenulation (Figure 10.6).
Ornamentation. The ornamentation consists of closely packed, rounded, irregular to pentagonal tubercles (Figure 9, Figure 10). All tubercles have well-marked crenulation at the base. On branchial plates the largest tubercles (7-8 per mm) are located along the lateral margin (Figure 9.3, 9.5) and most of them are low and pentagonal in shape. Toward the medial part of the plate, the size of tubercles decreases from 10-12 per mm (Figure 9.3) to 15-16 per mm (Figure 9.2). The tubercles are mostly low, almost flat and irregular to pentagonal in shape. Nevertheless, in some parts the tubercles may be higher and more dome-shaped (Figure 9.5). Lower, more closely packed and less varied are tubercles on the dorsal (Figure 10) and ventral plates whose shape is pentagonal or irregular. The difference is in the size of tubercles; on the dorsal plate tubercles are larger (12-14 per cm; Figure 10.2, 10.4) than those on the ventral plate (18-20 per cm). The cornual plate possesses dome-shaped tubercles (9-13 per cm) with a well-visible base crenulation (Figure 10.6).
Remarks. Revision of psammosteid collection described by Tarlo (1964, 1965) shows that some plates were incorrectly identified. The studied collection lacks several specimens of Guerichosteus described by Tarlo (1964, 1965): D.5.U.W., D.6.U.W., D.10.U.W., D.11.U.W., D.12.U.W., D.18.U.W, D.20.U.W., D.21.U.W., D.31.U.W., D.32.U.W., D.34.U.W., D.37.U.W., D.40.U.W., D.41.U.W, D.42.U.W., D.43.U.W., D.44.U.W., D.82.U.W., D.84.U.W.
 ZPAL Ag. III/4, originally described by Tarlo (1965; pl. XII, figs. 3-4) as the right half of the posterior end of a ventral plate, is in fact the right half of the dorsal plate (Figure 3.1-3). The edge considered by Tarlo (1965) as the margin of the posterior notch is irregular in outline, which has been likely formed by breaking the plate fragment.
ZPAL Ag. III/4, originally described by Tarlo (1965; pl. XII, figs. 3-4) as the right half of the posterior end of a ventral plate, is in fact the right half of the dorsal plate (Figure 3.1-3). The edge considered by Tarlo (1965) as the margin of the posterior notch is irregular in outline, which has been likely formed by breaking the plate fragment.
The part of the ZPAL Ag. III/1 dorsal plate illustrated in Figure 3.4-7 is recognized as the lateral edge rather than a posterior one as described by Tarlo (1965). Well visible growth lines with an almost straight posterior edge and roundly curved lateral margin resemble ZPAL Ag. III/4.
During the investigation of the collection an additional tip of the ZPAL Ag. III/14 branchial plate was found (presented in Tarlo, 1965: pl. X, fig. 6). It shows larger, anterior portion of the ventral and external ornamented surface (Figure 6.9-10).
The ZPAL Ag. III/22 pineal plate (Figure 8.19), recognized by Tarlo (1965), is more likely a scale as it resembles scales illustrated by Mark-Kurik (1965).
Neither the Gurichosteus kotanskii holotype D.37.U.W nor two other specimens (D.39.U.W and D.40.U.W) were found in the studied collection. Only ZPAL Ag. III/38 (in Tarlo (1965) D.38.U.W) shows the ornamentation similar to G. kozlowskii. According to Tarlo (1965), the tubercles of G. kotanskii are extremely large, but ZPAL Ag. III/38 has 8-10 tubercles per cm (diameter 1-1.25 mm), such as those observed in the G. kozlowskii branchial plate. The measurements of specimens not currently in the collection, but presented in Tarlo (1965), were taken from figures. So the holotype had approximately 7-10 tubercles per cm and specimen D.40.U.W had 5-9 tubercles per cm. The last specimen shows slightly larger tubercles than G. kozlowskii. However, this is insufficient to support the creation of the new species Guerichosteus kotanskii.
The ZPAL Ag. III/45 holotype of Guerichosteus lefeldii (Tarlo, 1965: pl. XII, fig. 9) is the antero-median part of the ventral plate with a visible posterior notch (Figure 4.1-2). Tarlo (1965) distinguished this genus based on ornamentation of minute, closely packed, rounded tubercles, approximately 0.3 mm in diameter. This study shows (similar to G. kozlowskii) ornamentation of small tubercles, approximately 0.5 mm (18-20 per cm). The tubercles of MUZ.PIG.1733.II.252 and ZPAL Ag. III/1080 G. kozlowskii specimens are slightly bigger than those of ZPAL Ag. III/45, albeit it may be due to the closer position of the tubercles to the growth center instead of marginal within former specimens. It should be classified as G. kozlowskii, such as a body scale of ZPAL Ag. III/47 (Tarlo 1965: pl. XII, fig. 8). Therefore, Guerichosteus lefeldii is considered here as a junior synonym of Guerichosteus kozlowskii.
No Guerichosteus kuczycki specimens (D.42.U.W., D.43.U.W, D.44.U.W.) were found in Tarlo’s collection, even the holotype (D.41.U.W.). The investigation of descriptions and figures suggests the affinity of G. kozlowskii. Tarlo decided to distinguish the species G. kulczyski from other species of Guerichosteus by its fairly small (diameter - 0.5 mm) and clearly separated tubercles. However, it resembles the ornamentation observed on the ventral plate, especially the small and pointed tubercles, of ZPAL Ag.III/45 (Figure 4.1).
The species erected by Tarlo (1965, 1966), G. kulczyckii, G. lefeldi and G. kotanskii, are in the range of variability of Guerichosteus kozlowskii and are here included in this species. Several specimens (ZPAL Ag. III/17, ZPAL Ag. III/24, ZPAL Ag. III/25, ZPAL Ag. III/26) included by Tarlo in Guerichosteus are moved to Hariosteus (see description below).
 The ventral plate of Guerichosteus displays the intermediate stage of development between Drepanaspis and all other members of the group (except Psammosteus, in which the posterior notch was reduced). The plate has a shallow and narrow depression rather than a notch. Whereas Drepanaspis lacks a posterior notch, it is well developed among other psammosteids. The improvement of this structure might be the result of the increase in body size and can be related to ecological adaptation.
The ventral plate of Guerichosteus displays the intermediate stage of development between Drepanaspis and all other members of the group (except Psammosteus, in which the posterior notch was reduced). The plate has a shallow and narrow depression rather than a notch. Whereas Drepanaspis lacks a posterior notch, it is well developed among other psammosteids. The improvement of this structure might be the result of the increase in body size and can be related to ecological adaptation.
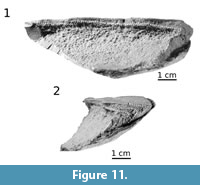
Apart from growth lines visible on the external surface, one branchial plate (ZPAL Ag. III/7) reveals a layered pattern of internal growth of the plate. The broken surface of the dorsal part exposes layers of the bone (Figure 11, hatched line). In median view, the middle vascular layer shows thin bone layers obliquely arranged to the dorsal surface of the plate and proximally thinned-out (Figure 11.1). At the posterior broken margin of the plate, the layers correspond to the shape of the branchial plate surface (Figure 11.2). The bone layers show forward and inward ossification, which also caused an increase of the plate thickness. Therefore, it is consistent with the medial direction of ossification, expressed by the presence of growth lines on ornamented external surface of the branchial plate.
Occurrence. Lower Devonian (Emsian), Placoderm Sandstone, Podłazie Hill near Daleszyce, Holy Cross Mountains, Poland.
Genus HARIOSTEUS Tarlo, 1964
Type species. Hariosteus kielanae Tarlo, 1964, by original designation.
Hariosteus kielanae
Figure 12, Figure 13, Figure 14, Figure 15; Appendix 22, Appendix 23, Appendix 24, Appendix 25, Appendix 26, Appendix 27, Appendix 28, Appendix 29
 . 1957. Psammosteid type B; Tarlo 1957, p. 230, pl. 1, fig. 2.
. 1957. Psammosteid type B; Tarlo 1957, p. 230, pl. 1, fig. 2.
. 1965. Hariosteus kielanae; Tarlo 1965: pp. 52-55, pl. XI, fig. 14, pl. XIII, figs. 3, 6-10.
v. 1965. Hariosteus kielanae; Tarlo 1965: pp. 52-55, text-figs. 9, 10, pl. XIII, figs. 1-2, 4, 11.
v. 1965. Guerichosteus kozlowskii; Tarlo 1965: pp. 47, fig. 7D-E, pl. IX, fig. 2A-B, pl. XI, figs. 1, 5.
Diagnosis. Branchial plate wide and short, ornamentation composed of well separated, crenulated tubercles with smaller accessory tubercles between them. Presence of alternate bands of larger and smaller ornamentation, giving distinguishing growth zones, especially on the dorsal median plate (modified from Tarlo, 1965).
Description
Dorsal plate. Apparent left posterior fragment (ZPAL Ag. III/48) of a dorsal plate (Figure 12.1-12.3, S22) preserves both sides, external surface with tubercles and visceral. The fragment is flat, 44 mm long, 63 mm wide and 4 mm thick. Well-recognizable growth lines reflect the posterior margin. The visceral surface is smooth with several grooves and ribs, mostly concentrated near the plate margin (Figure 12.2). They both mirror the growth zones marked in external, superficial ornamentation. Two other thin and flat specimens, MZ-VIII/Vp-549 (Figure 12.4) and ZPAL Ag. III/50 (Figure 12.5, 12.6), are possibly fragments of the dorsal median (Figure 12.4-5). The latter one is the largest specimen and shows very well-preserved tubercles characteristic for Hariosteus.
 Branchial plate. There are branchial plates of two specimens present in the material, ZPAL Ag. III/17 (Figure 13.1-3; Appendix 23) and MUZ.PIG.1733.II.185 (Figure 13.4-5; Appendix 24); specimens are in order, right and left branchial plate. ZPAL Ag. III/17 preserves both sides of the plate (dorsal and ventral). The plate is 49 mm wide and 57 mm long. The plate is dorsally convex. Its lateral margin is strongly arched, and the posterior margin creates a slight sinus. The posterior part of the lateral margin and the posterior margin form a right angle. The ventral side of the plate is weathered, and only the most anterior and posterior parts of the lateral edge have preserved tubercles (Figure 13.2). The dorsal surface with tubercles is triangular with a concave posterior margin and a waving medial edge. The Branchial plate of MUZ.PIG.1733.II.185 is similar to the one of ZPAL Ag. III/17 where only the dorsal side is preserved. The overall shape of the plates is comparable. However, the lateral margin is more arched than in ZPAL Ag. III/17, and the posterior edge of the dorsal ornamented surface is deeper (Figure 13.4). The plate is 55 mm wide and 45 mm long. Its thickness, measured anteriorly along the broken antero-lateral edge, is 7 mm and gradually increases to 10 mm. ZPAL Ag III/56 shows a distal tip of the ventral side of the branchial plate. The plate preserves a fragment of the posterior and lateral margin (Figure 13.6).
Branchial plate. There are branchial plates of two specimens present in the material, ZPAL Ag. III/17 (Figure 13.1-3; Appendix 23) and MUZ.PIG.1733.II.185 (Figure 13.4-5; Appendix 24); specimens are in order, right and left branchial plate. ZPAL Ag. III/17 preserves both sides of the plate (dorsal and ventral). The plate is 49 mm wide and 57 mm long. The plate is dorsally convex. Its lateral margin is strongly arched, and the posterior margin creates a slight sinus. The posterior part of the lateral margin and the posterior margin form a right angle. The ventral side of the plate is weathered, and only the most anterior and posterior parts of the lateral edge have preserved tubercles (Figure 13.2). The dorsal surface with tubercles is triangular with a concave posterior margin and a waving medial edge. The Branchial plate of MUZ.PIG.1733.II.185 is similar to the one of ZPAL Ag. III/17 where only the dorsal side is preserved. The overall shape of the plates is comparable. However, the lateral margin is more arched than in ZPAL Ag. III/17, and the posterior edge of the dorsal ornamented surface is deeper (Figure 13.4). The plate is 55 mm wide and 45 mm long. Its thickness, measured anteriorly along the broken antero-lateral edge, is 7 mm and gradually increases to 10 mm. ZPAL Ag III/56 shows a distal tip of the ventral side of the branchial plate. The plate preserves a fragment of the posterior and lateral margin (Figure 13.6).
The smaller ZPAL Ag III/24 specimen, described as an oral plate by Tarlo (1965, fig. 7D-E), seems to be in fact a distal tip of the branchial plate. The dorsal side of the plate is visible (Figure 13.7; Appendix 25). The lateral margin is arched similarly to two larger specimens, ZPAL Ag. III/17 and MUZ.PIG.1733.II.185. Also the thickness (8 mm) is comparable. However, whether the specimen represents a small individual or if it is just a broken fragment of a plate is inconclusive, albeit the former interpretation is likely more probable. The tubercles along the medial margin of the plate seem to be natural, at least in the anterior half, which supports the assumption that the specimen is a small individual branchial plate.
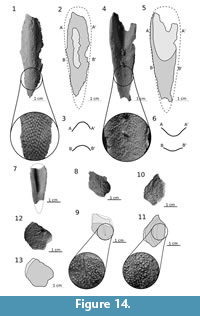 Ridge scales. Three specimens (ZPAL Ag. III/25, ZPAL Ag. III/28, MZ-VIII/Vp-67) include ridge scales. An incomplete ridge scale is ZPAL Ag III/28 (Figure 14.1-3; Appendix 27). The external surface covered with tubercles is visible, although the most prominent part lacks tubercles (probably due to post-mortem abrasion). The specimen is 60 mm long, the maximum width is 15 mm and its height is 10 mm. The scales are relatively high and U-shaped in transverse section. The more complete ZPAL Ag. III/28 shows the internal, posterior and right lateral part of the external surface of the scale (Figure 14.4-6; Appendix 26). Its length is 65 mm and maximum width is 20 mm. More than a half of the specimen (35 mm) has an unornamented proximal part which was connected with soft tissue. In the smaller MZ-VIII/Vp-67 (ZPAL Ag. III/29) the proximal smooth part reaches more than a half of the specimen (it is 21 mm of about 38 mm of the estimated length of this specimen; Figure 14.7). The ridge scales show high and slightly elongated tubercles on the external surface (ZPAL Ag. III/25; Figure 14.1), while near the lateral edges tubercles are more elongated (ZPAL Ag. III/28; Figure 14.2). The internal surface shows radially propagating, loosely dense tubercles. Tubercles change from being round in front and long and thin in back parts. The same pattern is visible on MZ-VIII/Vp-67 (Figure 14.7; Appendix 28), which is 25 mm long and 14 mm of its maximum width.
Ridge scales. Three specimens (ZPAL Ag. III/25, ZPAL Ag. III/28, MZ-VIII/Vp-67) include ridge scales. An incomplete ridge scale is ZPAL Ag III/28 (Figure 14.1-3; Appendix 27). The external surface covered with tubercles is visible, although the most prominent part lacks tubercles (probably due to post-mortem abrasion). The specimen is 60 mm long, the maximum width is 15 mm and its height is 10 mm. The scales are relatively high and U-shaped in transverse section. The more complete ZPAL Ag. III/28 shows the internal, posterior and right lateral part of the external surface of the scale (Figure 14.4-6; Appendix 26). Its length is 65 mm and maximum width is 20 mm. More than a half of the specimen (35 mm) has an unornamented proximal part which was connected with soft tissue. In the smaller MZ-VIII/Vp-67 (ZPAL Ag. III/29) the proximal smooth part reaches more than a half of the specimen (it is 21 mm of about 38 mm of the estimated length of this specimen; Figure 14.7). The ridge scales show high and slightly elongated tubercles on the external surface (ZPAL Ag. III/25; Figure 14.1), while near the lateral edges tubercles are more elongated (ZPAL Ag. III/28; Figure 14.2). The internal surface shows radially propagating, loosely dense tubercles. Tubercles change from being round in front and long and thin in back parts. The same pattern is visible on MZ-VIII/Vp-67 (Figure 14.7; Appendix 28), which is 25 mm long and 14 mm of its maximum width.
Trunk scales. In the studied material several trunk scales are recognized. Scale MUZ.PIG.1733.II.309 is 21 mm long,15 mm high and shows a wide (about 7 mm) overlapping area (Figure 14.8-9; Appendix 29). Near the center of the scale extends a groove of lateral line, which ends with a minute pore. A larger body scale MZ-VIII/Vp-410 (Figure 14.10-11) is rather high (24 mm) and short (17 mm). The last one (MUZ.PIG.1733.II.184) is more pentagonal in shape, 17 mm wide, 21 mm long and 3 mm thick (Figure 14.12-13). The proximal margin is convex with narrow unornamented area.
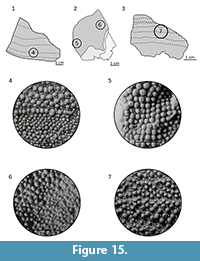 Ornamentation. Tubercles vary from 0.1 to 1.2 mm diameter. The larger ones (on branchial plates) are placed near the edge and decrease in size medially. Between larger, dome-shaped tubercles there are low and cone-shaped accessory tubercles (Figure 15.4), which may be absent on branchial plates (Figure 15.5). Narrow, marginal cusplets reach the cupola-shaped apex of both tubercles and accessory tubercles (Figure 15.6). The characteristic growth zones of alternate bands of larger and smaller tubercles are visible best on the median dorsal plate (Figure 15.7), while near the branchials margin there is a lack of them or they are barely visible (Figure 15.5). The accessory tubercles, variously arranged, may fill the space between larger tubercles in disarray or they can encircle larger tubercles.
Ornamentation. Tubercles vary from 0.1 to 1.2 mm diameter. The larger ones (on branchial plates) are placed near the edge and decrease in size medially. Between larger, dome-shaped tubercles there are low and cone-shaped accessory tubercles (Figure 15.4), which may be absent on branchial plates (Figure 15.5). Narrow, marginal cusplets reach the cupola-shaped apex of both tubercles and accessory tubercles (Figure 15.6). The characteristic growth zones of alternate bands of larger and smaller tubercles are visible best on the median dorsal plate (Figure 15.7), while near the branchials margin there is a lack of them or they are barely visible (Figure 15.5). The accessory tubercles, variously arranged, may fill the space between larger tubercles in disarray or they can encircle larger tubercles.
Remarks. The studied collection lacks several specimens of Hariosteus described by Tarlo (1964, 1965): D.52.U.W, D.54.U.W, D.55.U.W, D.58.U.W, D.59.U.W, D.61.U.W, D.62.U.W.
Tarlo (1964, 1965) described two species of the genus Hariosteus kielanae and Hariosteus lobanowskii. The holotype of Hariosteus lobanowskii (ZPAL Ag. III/60) is poorly preserved, especially tubercles. The ornamentation may vary depending on a specific plate or part of the plate where the tubercles are located. Therefore, a small number of the available specimens may not reveal full variability among members of the Hariosteus genus. For example, in Guerichosteus, one specimen reveals slightly different ornamentation. Also ZPAL Ag. III/61 and ZPAL Ag. III/62 are missing and are not illustrated in the Tarlo monograph (1965). Therefore, the species H. lobanowskii erected by Tarlo is considered here as invalid.
Tarlo (1965) interpretation of ZPAL Ag. III/51 as a postorbital seems to be doubtful. The plate is incomplete, and the most characteristic feature (except the ornamentation) are almost straight growth lines extended perpendicular to the bent edge (Figure 12.7-8). Such bending is not restricted to postorbital plates. In Psammolepis venyukovi and Ganosteus stelatus, the proximal ventral surface of the branchial plate looks similar (Obruchev and Mark-Kurik, 1965). Moreover, the branchial plate of Hariosteus is thick. This is why this fragment is probably more a proximal part of a larger individual in which the edge is not as rounded as in ZPAL Ag. III/17 or MUZ.PIG.1733.II.353.
ZPAL Ag. III/83, considered as Gurichosteus kozlowskii by Tarlo (1965), is a fragment of a branchial plate of Hariosteus kielanae. The edge of the plate is rounded with nearly parallel dorsal and ventral surfaces similar to the one of Hariosteus, in contrast to the more triangular and angled edge of Guerichosteus. Also the ornamentation shows several accessory tubercles like in Hariosteus.
The studied material of Hariosteus is insufficient to make an overall restoration of the shields and body. The most complete branchial plate with overall proportions resembles those of Pycnosteidae (sensu Obruchev and Mark Kurik, 1965). Both width and length of the plate is nearly the same and resembles the young stage of Schizosteus spendens and Tartuosteus giganteus. This is what distinguishes it from Guerichosteus kozlowskii, having the branchial plate long and narrow as in Drepanaspis.
In recent papers, Hariosteus is considered as a junior synonym of Guerichosteus (Glinsky, 2018; Glinskiy and Pinakhina, 2018). However, apart from differences in branchial plates, the ornamentation is also clearly different in these two genera. Moreover, Hariosteus features may suggest its affinity to younger genera such as Pycnosteus or Schizosteus. Thus, if we consider the taxonomic division by Tarlo (1964, 1965) or Glinsky (2018), this genus should be excluded from the Guerichosidae and placed in Pycnosteidae. The ornamentation is also suggestive for the latter group of psammosteids.
Occurrence. Lower Devonian (Emsian), Placoderm Sandstone, Daleszyce, near Kielce, the Holy Cross Mountains, Poland.
RECONSTRUCTION
The knowledge of an animal’s appearance is the key to any considerations regarding not only its taxonomic status, but also its biomechanical and ecological aspects. The reconstruction of the psammosteids can mostly be based on Drepanaspis gemuendenesis as it is the best known representative of the group. But specimens that consist of complete armor are flattened, while those preserved with no deformation are mostly isolated plates. Also, preservation of naturally connected plates is rare (as in the case of the specimens from Holy Cross Mountains). The new findings of the complete ventral plate, along with the posterior tip of the branchial plate, orbital plate and complex plate together (and fragments of plates), as well as the application of computer methods, reveal not only a more accurate view of Guerichosteus kozlowskii, but they also provide new insights into Drepanaspi s and other psammosteid morphology. In this study only Guerichosteus kozlowskii may be reconstructed, as the Hariosteus material is insufficient.
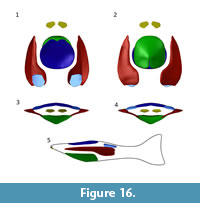 The first step of Guerichosteus kozlowskii reconstruction was a single plate restoration. Most plates (orbital plate, postorbital plate, ventral median plate, cornual plate and complex plate) are represented by at least one slightly damaged or complete plate; the median dorsal plate and branchial plate (beside one small/juvenile specimen) are broken fragments. Next all 3D reconstructed plate models were imported to the Blender software. Because the Guerichosteus remains came from specimens of all sizes, they had to be scaled (reduced or enlarged). The reference size was the (MUZ.PIG.1733.II.353) ventral plate. The size relation of individual plates was based on the reconstructions of Drepanaspis gemuendenensis by Gross (1963). Then, the individual plates were fitted together in the proper configuration (Figure 16). In ventral and lateral views, the curvature of the lateral margin of the ventral plate reflects the inner margin of the ventral part of the branchial plates. Furthermore, the transition between the plates (looking at the transverse section) is smooth, and the inclination of the ventral plate corresponds to the concave ventral side of the branchial plate (Figure 16.3-4). In dorsal view, the external surface of the branchial plate changes from flat (in the posterior part of small specimens) to slightly concave (in the anterior part of larger specimens). The ventral surface of the plate is concave. It widens in the anterior tip and extends parallel to the lateral margin. At the midpoint of the plate length it expands strongly and forms a broad postero-medial projection. In the smallest specimen, ZPAL Ag.III/1089, the ventral surface is triangular.
The first step of Guerichosteus kozlowskii reconstruction was a single plate restoration. Most plates (orbital plate, postorbital plate, ventral median plate, cornual plate and complex plate) are represented by at least one slightly damaged or complete plate; the median dorsal plate and branchial plate (beside one small/juvenile specimen) are broken fragments. Next all 3D reconstructed plate models were imported to the Blender software. Because the Guerichosteus remains came from specimens of all sizes, they had to be scaled (reduced or enlarged). The reference size was the (MUZ.PIG.1733.II.353) ventral plate. The size relation of individual plates was based on the reconstructions of Drepanaspis gemuendenensis by Gross (1963). Then, the individual plates were fitted together in the proper configuration (Figure 16). In ventral and lateral views, the curvature of the lateral margin of the ventral plate reflects the inner margin of the ventral part of the branchial plates. Furthermore, the transition between the plates (looking at the transverse section) is smooth, and the inclination of the ventral plate corresponds to the concave ventral side of the branchial plate (Figure 16.3-4). In dorsal view, the external surface of the branchial plate changes from flat (in the posterior part of small specimens) to slightly concave (in the anterior part of larger specimens). The ventral surface of the plate is concave. It widens in the anterior tip and extends parallel to the lateral margin. At the midpoint of the plate length it expands strongly and forms a broad postero-medial projection. In the smallest specimen, ZPAL Ag.III/1089, the ventral surface is triangular.
There were some problems with setting and reconstructing the dorsal plate properly, as several pieces represent distinct parts of the plate, although the dorsal plates show clear growth lines reflecting the subsequent ontogenetic stages. As it is shown in Figure 16 (Appendix 30), the lateral curvature of the plate is not smoothly arched as it was shown in the previous reconstruction of the psammosteid Drepanaspis, but it is slightly S-shaped and resembles more pteraspidids, such as Pteraspis or Errivaspis. Therefore, similar to these specimens the anterior margin of the plate is set slightly back in relation to the ventral plate, while the posterior edge of plates reaches the same point (Figure 16.1-2, 16.5).
The cornual plate is placed similar to Drepanaspis (Gross, 1963). The most anterior part of the body is limited by a complex plate. The dorsal margin of the plate is located at the point of the lateral margin of branchial plates, similarly to the Gross (1963) restoration of Drepanaspis. It is unknown how far the plate reached forward; here (in lateral view) it is placed in the extension of the antero-ventral surface of the ventral plate (Figure 16.5).
CONCLUSIONS
Investigation of the morphology, with consideration of ornamentation features, of psammosteid remains from “Placoderm Sandstone” supports the establishment of the genera Guerichosteus and Hariosteus by Tarlo (1964, 1965). The reexamination of the studied material reveals a new interpretation of some specimens such as ZPAL. Ag. III/4. Tarlo (1965) described it as a ventral plate; however, it seems to be the dorsal plate. Detailed examination of the Guerihosteus species G. kotanskii, G. kulczyckii and G. lefeldi revealed that they were junior synonyms of the revised species Guerichosteus kozlowskii Tarlo, 1965. The analysis of ornamentation and plate morphology of Hariosteus kotanskii indicates their resemblance to Hariosteus kielanae. Thus H. kotanskii should be included in the species Hariosteus kielanae Tarlo, 1965. Hariosteus lobanowskii is considered here as invalid (nomen dubium).
Guerichosteus kozlowskii lacks the typically developed posterior notch. Instead of having a slit, it has a low depression. Also the Early Devonian Drepanaspis gemuendenensis seems to have a similarly developed posterior notch as Guerichosteus, although in the former species the analysis and revision is problematic due to taphonomic issues. The development of the posterior notch from a depression might have resulted from enlargement of the psammosteids body during evolution. The ventral plate of Drepanaspis gemuendenensis resembles the one of Guerichosteus kozlowskii. Therefore, it is probably deeper and not so roundly arched as shown by Gross (1963). The three-dimensional restoration of Guerichosteus kozlowskii demonstrates its more streamlined body shape in comparison to the previous Drepanaspis gemuendenensis reconstruction.
ACKNOWLEDGMENTS
The material obtained during fieldwork in 2011-2013 was financed by the Ministry of Science and Higher Education (grant No. IP2010 041470 awarded to Piotr Szrek). P. Szrek and G. Niedźwiedzki for cooperation and all support. I would like to thank B. Studencka, C. Krawczyński and D. Nast, the Museum of the Earth of the Polish Academy of Sciences in Warsaw employees, for their support during collection studying. I also thank P. Gorzelak (Institute of Paleobiology, Polish Academy of Sciences) and S. Tasakowski for the linguistic correction of the manuscript. Furthermore, I thank three anonymous reviewers for their helpful comments and linguistic improvement.
REFERENCES
Berg, L.S. 1940. Classification of fishes, both recent and fossil. Trudy Zoologicheskogo Institua Akademii Nauk SSSR, 5:85-517.
Blieck ARM. 1984. Hétérostracés, Pteraspidiformes: systemastique, phylogenie, biostratigraphie et biogeographie, p. 1-205. In Cahiers de Paléontologie (section Vertébrés). Paris: Editions du CNRS.
Dean, 1904. Fishes of Gemunden. Science, 19:64-65.
Elliott, D.K., Mark-Kurik, E., and Daeschler, E. B. 2004. A revision of Obruchevia (Psammosteida: Heterostraci) and a description of a new obrucheviid from the Late Devonian of the Canadian Arctic. Acta Universitatis Latviensis, 679:22-45.
Falkingham, P.L. 2012. Acquisition of high resolution 3D models using free, open-source, photogrammetric software. Palaeontologia Electronica Vol. 15, Issue 1; 1T:15p; https://doi.org/10.26879/264
palaeo-electronica.org/content/issue-1-2012-technical-articles/92-3d-photogrammetry
Fijałkowska-Mader, A. and Malec, J. 2011. Biostratigraphy of the Emsian to Eifelian in the Holy Cross Mountains (Poland). Geological Quarterly, 55:109-138.
Glinskiy, V. 2018. Phylogenetic relationships of psammosteid heterostracans (Pteraspidiformes), Devonian jawless vertebrates. Biological Communications, 62:219-243. https://doi.org/10.21638/11701/spbu03.2017.402
Glinskiy, V.N. and Pinakhina, D.V. 2018. New data on psammosteid heterostracans (Pteraspidomorpha) and acanthodians (Acanthodii) from the Pärnu Regional Stage (Lower Eifelian, Middle Devonian) of Estonia. Estonian Journal of Earth Sciences, 67:76. https://doi.org/10.3176/earth.2018.05
Gross, W. 1963. Drepanaspis gemuendenensis Schlüter, Neuuntersuchung. Palaeonogreaphica, 121A:133-155, Stuttgart.
Halstead, L.B. 1973. August. The heterostracan fishes. Biological Reviews, 48:279-332. https://doi.org/10.1111/j.1469-185X.1973.tb01005.x
Janvier, P. 1996. Early Vertebrates. Oxford, Oxford Science Publications, Clarendon Press.
Kiaer, J. 1915. Upper Devonian Fish remains from Ellesmere Land, with remarks on Drepanaspis. Report of the Second Norwegian Arctic expedition in the "Fram" 1898-1902, 33:1-72. Kristania.
Kiaer, J. 1928. The structure of the mouth if the oldest know vertebrates, pteraspis and cephalaspis. Palaeobiologica, 1:117-134, Vienna.
Kruszewski, Ł. and Dec, M. 2018. Plumbogummite group minerals in Lower Devonian placoderm sandstones from Podłazie Hill, Holy Cross Mountains, Poland. Geological Quarterly, 62:353–360. https://doi.org/10.7306/gq.1410
Mallison, H. and Wings, O. 2014. Photogrammetry in paleontology - a practical guide. Journal of Paleontological Techniques, 12:1-31.
Lankester E. 1868. A monograph of the fishes of the Old Red Sandstone of Britain. Part 1. The Cephalaspidae. Palaeontological Society, London, p. 1-33.
Obruchev, D. 1943. A new restoration of Drepanaspis. The Proceedings of the USSR Academy of Sciences, 41:268-271.
Obruchev, D. and Mark-Kurik, E. 1965. Psammosteids of the Devonian of the USSR. Akademiya Nauk Estonskoy SSR, Institut Geologii, Trudy. Tallinn, Estonia.
Pernègre, V.N. and Elliott, D.K. 2008. Phylogeny of the Pteraspidiformes (Heterostraci), Silurian-Devonian jawless vertebrates. Zoologica Scripta, 37:391-403. https://doi.org/10.1111/j.1463-6409.2008.00333.x
Randle, E. and Sansom, R.S. 2017a. Exploring phylogenetic relationships of Pteraspidiformes heterostracans (stem-gnathostomes) using continuous and discrete characters. Journal of Systematic Palaeontology, 15:583-599, https://doi.org/ 10.1080/14772019.2016.1208293
Randle, E. and Sansom, R.S 2017b. Phylogenetic relationships of the ‘higher heterostracans’ (Heterostraci: Pteraspidiformes and Cyathaspididae), extinct jawless vertebrates. Zoological Journal of the Linnean Society, 181:910-926. https://doi.org/10.1093/zoolinnean/zlx025
Stensiö, E.A. 1927. The Downtonian and Devonian Vertebrates of Spitsbergen. I. Family Cephalaspidae. Skrifter om Svalbard og Nordishavet, 12:1-391.
Stetson, H.C. 1931. Studies on the morphology of the Heterostraci. The Journal of Geology, 39:141-154. https://doi.org/10.1086/623803
Szrek, P., Niedźwiedzki, G., and Dec, M. 2014. Storm origin of bone-bearing beds in the Lower Devonian placoderm sand stone from Podłazie Hill (Holy Cross Mountains, central Poland). Geological Quarterly, 58:795-806. https://doi.org/10.7306/gq.1191
Szulczewski, M. and Porębski, S. 2008. Stop 1 - Bukowa Góra, Lower Devonian, p. 18-37. In Pieńkowski, G. and Uchman, A (ed.). Ichnological Sites of Poland. The Holy Cross Mountains and the Carpathian Flysch. The Second International Congress on Ichnology. Cracow, Poland, August 2-September 8. The Pre-Congress and Post-Congress Field Trip Guidebook, Warszawa.
Tarnowska, M. 1976. Lithological correlation of the Lower Devonian in the eastern part of the Holy Cross Mountains (in Polish with English summary). Biuletyn Instytutu Geologicznego, 296:75-115.
Tarnowska, M., 1981. Lower Devonian in the central part of the Holy Cross Mountains (in Polish with English summary). p. 57-67 In Przewodnik 53 Zjazdu Polskiego Towarzystwa Geologicznego, Kielce. Instytut Geologiczny, Warszawa.
Tarlo, L.B. 1957. A preliminary note on new ostracoderms from the Lower Devonian (Emsian) of central Poland. Acta Palaeontologica Polonica, 2: 225-233.
Tarlo, L.B. 1961. Psammosteids from the Middle and Upper Devonian of Scotland. The Quarterly Journal of the Geological Society of London, 117:367-402.
Tarlo, L.B. 1964.Psammosteiformes (Agnatha) - a review with descriptions of new material from the Lower Devonian of Poland. I - general part. Palaeontologia Polonica, 13:1-135.
Tarlo, L.B. 1965. Psammosteiformes (Agnatha) - a review with descriptions of new material from the Lower Devonian of Poland. II - systematic part. Palaeontologia Polonica, 15:1-168.
Traquair, R.H. 1896. The extinct vertebrata of the Moray Firth area, p. 235-285. In Harvie-Brown, J.H. and Buckley, T.E. (eds.), Vertebrate Fauna of the Moray Basin. D. Douglas, Edinburgh.
Traquair, R.H. 1899. Report on fossil fishes collected by the geological survey of Scotland in the Silurian rocks of the South of Scotland. Transitions of the Royal Society of the Edinburgh, 39:827-864.
Traquair, R.H. 1903. The Lower Devonian fishes of Gemünden. Transitions of the Royal Society of the Edinburgh, 40:723-739.
