

 Pteropoda (Mollusca, Gastropoda, Thecosomata) from the Eocene/Oligocene boundary interval of three cored boreholes in southern coastal Tanzania and their response to the global cooling event
Pteropoda (Mollusca, Gastropoda, Thecosomata) from the Eocene/Oligocene boundary interval of three cored boreholes in southern coastal Tanzania and their response to the global cooling event
Article number: 20.3.52A
https://doi.org/10.26879/733
Copyright Palaeontological Association, October 2017
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 18 October 2016. Acceptance: 26 September 2017
{flike id=2024}
ABSTRACT
The Eocene - Oligocene Transition was a period of major climatic and oceanographic change, resulting in widespread biotic overturning. However, the record of many marine organisms remains patchy. Planktic Mollusca (Pteropoda) from three cored boreholes spanning the Eocene/Oligocene boundary (EOB) in southern coastal Tanzania are represented by eight species, three of which are introduced as new: Heliconoides nikkieae sp. nov., Limacina tanzaniaensis sp. nov. and Limacina timi sp. nov. Three of the other species can only be identified in open nomenclature. The two most commonly occurring species, H. nikkieae and L. timi, straddle the EOB without noticeable loss in abundance. Two species, Limacina robusta (Eames, 1952) and L. tanzaniaensis disappear before the EOB. The species Bovicornu aff. eocenense Meyer, 1886 disappears shortly after the EOB. Two species were only found in a single sample each, in the Eocene part of the succession. Response to changing environmental conditions seems to be demonstrated by two or three of the pteropod species that become extinct before or shortly after the EOB.
Laura J. Cotton. Naturalis Biodiversity Center, P.O. Box 9517, 2300 RA Leiden, The Netherlands; present address: Florida Museum of Natural History, 1659 Museum Road, University of Florida, Gainesville, FL 32611, USA and Department of Geological Sciences, University of Florida, 241 Williamson Hall, Gainesville, FL32611, USA ; lcotton@flmnh.ufl.edu
Arie W. Janssen. Naturalis Biodiversity Center, P.O. Box 9517, 2300 RA Leiden, The Netherlands; ariewjanssen@gmail.com
Paul N. Pearson. School of Earth and Ocean Sciences, Cardiff University, Main Building, Park Place, Cardiff. CF10 3AT, United Kingdom; PearsonP@cardiff.ac.uk
Rens van Driel. Buys Ballotsingel 85A, 3112 JD, Schiedam, The Netherlands; rensvdriel@gmail.com
Key words: planktic Gastropoda; Altaspiratella; Heliconoides; Limacina; Bovicornu; new species; geographical distribution; vertical ranges; Eocene-Oligocene transition (EOT)
Final citation: Cotton, Laura J., Janssen, Arie W., Pearson, Paul N., and van Driel, Rens. 2017. Pteropoda (Mollusca, Gastropoda, Thecosomata) from the Eocene/Oligocene boundary interval of three cored boreholes in southern coastal Tanzania and their response to the global cooling event. Palaeontologia Electronica 20.3.52A: 1-21. https://doi.org/10.26879/733
palaeo-electronica.org/content/2017/2024-pteropoda-from-tanzania
Copyright: October 2017 Palaeontology Association.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
http://zoobank.org/2B5D7C0F-1AE0-4310-9751-97FC6FD64475
INTRODUCTION
The Eocene-Oligocene transition (EOT) was one of the most dramatic events of the Cenozoic, associated with major climatic disruption and widespread biotic turnover in both marine and terrestrial realms (see Coxall and Pearson, 2007 for review). High-resolution studies of the marine response have been carried out using planktic and benthic foraminifera, and nanno-fossils (e.g., Zachos et al., 1994; Molina et al., 1993; Diester-Haass et al., 2001; Lui et al., 2004; Wade and Pearson, 2008; Cotton and Pearson, 2011; Cotton et al., 2014). However, records of other marine groups are often patchy. Pteropods are planktic gastropods and are increasingly being used in studies of recent ocean records and climate change (Fabry, 1990; Wall-Palmer et al., 2012; Bednaršek and Ohman, 2015; references therein). Their aragonitic shells are susceptible to dissolution, making them excellent model organisms to study ocean acidification. However, palaeoecological data for older pteropod records are relatively uncommon, yet may add important information to the overall understanding of climatic impacts in the marine realm. Pteropods have a well-documented fossil record, but many regions remain understudied, and early well-preserved records are therefore exceptionally important (e.g., Janssen et al., 2016).
The Eocene-Oligocene Transition (EOT) is an extended period of global oceanographic and climatic perturbation spanning between 33.5 - 34 Ma. The Eocene/Oligocene Boundary (EOB) is defined by the last occurrence ofIn addition, the site contains many the planktic foraminiferal family Hantkeninidae at the Global Boundary Stratotype-Section and Point (GSSP) at Massignano, near Ancona, on the Italian Adriatic coast (Premoli Silva and Jenkins, 1993), which occurs at 33.7 Ma on the timescale of Cande and Kent (1995). However, an unreliable magnetostratigraphy and stable-isotope stratigraphy (Bodiselitsch et al., 2004) and lack of carbonate (macro)fossils (pers. observ. AWJ, August 1992) in the Massignano section prevented global correlation with climatic events. The additional fact that the occurrence of Hantkeninidae is mostly restricted to tropical and sub-tropical environments also made long-distance correlations (e.g., to the North Sea Basin and in particular to the Priabonian stratotype) problematic. Subsequent studies of Deep Sea Drilling Project Site 522, showed that the extinction preceded the most positive oxygen isotopes of the EOT, representing the Early Oligocene Glacial Maximum (Zachos et al., 1994; Liu et al., 2004). More recently this idea has been further refined.
The Kilwa District in southern Tanzania contains an apparently complete succession through the EOT, which was recovered by three Tanzanian Drilling Project boreholes (TDP 11, 12 and 17; Nicholas et al., 2006; Pearson et al., 2008). The dominant lithology consists of hemipelagic clays interspersed with debris flow limestones (Nicholas et al. 2006). These clays contain exceptionally well-preserved calcareous micro- and nannofossils, including aragonitic preservation (Pearson et al., 2008; Wade and Pearson, 2008; Bown et al., 2008). This, coupled with the expanded nature of the sediments, allowed for high resolution stratigraphy chemo-and biostratigraphy to be carried out and precise correlation to global records. This placed, for the first time, the extinction of the Hantkeninidae between the two positive isotope shifts of the transition (Pearson et al., 2008; Wade and Pearson, 2008); the first of which is attributed to temperature change and the second largely to ice-volume (Lear et al., 2008). In addition, the site contains many other exceptionally well-preserved calcareous micro-fossils, including larger benthic foraminifera (Cotton and Pearson, 2011), bryozoans (Martino et al., 2017), dasycladaceae, ostracodes and also benthic molluscs (on which a paper is in preparation), making Tanzania an exceptionally important site for the study of comparatively less ubiquitous fossils across the EOT. Studies of the benthic molluscan record have shown that overall numbers of mollusc shells increase across the boundary. However, that study mainly examined the benthic forms. Here we present a detailed record of occurrences through the EOT interval and taxonomy of specifically the pteropod fauna from TDP 11, 12 and 17 (see Nicholas et al., 2006; Cotton and Pearson, 2011, for details on these boreholes). Though relatively few specimens were found due to the small sample size, the often excellent preservation and well-correlated nature of the record offers a unique insight into a region where no pteropod records were previously known.
MATERIAL AND METHODS
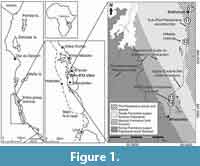 The Tanzania Drilling Project (TDP) was initiated in 2002 (Pearson et al., 2004; Nicholas et al., 2006, 2007) after preliminary field observations beginning in 1998 (Pearson et al., 2001) on numerous outcrops between the towns of Kilwa and Lindi, southern Tanzania, had demonstrated the presence of well-preserved microfossil assemblages of Cretaceous to Paleogene age. The TDP focused on generating litho- and biostratigraphic, geochemical, micropalaeontological and palaeoclimatic records, and resulted in recovery of over 40 cored boreholes, covering Cretaceous (Aptian) to Paleogene (Oligocene) sediments (Pearson et al., 2004, 2006; Nicholas et al., 2006; Jiminez-Berrucoso et al., 2010, 2012, 2015). These sediments are formally defined as the Kilwa Group and were initially subdivided into four formations: The Nangurukuru, Kivinje, Masoko, and Pande formations (Nicholas et al., 2006), to which a fifth was recently added, the Lindi formation (Jiminez-Berrucoso et al., 2015). The Pande Formation contains an apparently locally conformable succession through the EOT and was recovered in three boreholes drilled approximately along strike of each other; a minor hiatus is inferred at TDP Sites 11 and 17 but TDP 12 appears conformable (Figure 1).
The Tanzania Drilling Project (TDP) was initiated in 2002 (Pearson et al., 2004; Nicholas et al., 2006, 2007) after preliminary field observations beginning in 1998 (Pearson et al., 2001) on numerous outcrops between the towns of Kilwa and Lindi, southern Tanzania, had demonstrated the presence of well-preserved microfossil assemblages of Cretaceous to Paleogene age. The TDP focused on generating litho- and biostratigraphic, geochemical, micropalaeontological and palaeoclimatic records, and resulted in recovery of over 40 cored boreholes, covering Cretaceous (Aptian) to Paleogene (Oligocene) sediments (Pearson et al., 2004, 2006; Nicholas et al., 2006; Jiminez-Berrucoso et al., 2010, 2012, 2015). These sediments are formally defined as the Kilwa Group and were initially subdivided into four formations: The Nangurukuru, Kivinje, Masoko, and Pande formations (Nicholas et al., 2006), to which a fifth was recently added, the Lindi formation (Jiminez-Berrucoso et al., 2015). The Pande Formation contains an apparently locally conformable succession through the EOT and was recovered in three boreholes drilled approximately along strike of each other; a minor hiatus is inferred at TDP Sites 11 and 17 but TDP 12 appears conformable (Figure 1).
TDP 11. South of Stakishari (Tanzania, Kilwa region), coordinates UTM 37L 560250 8983211; pteropod specimens are available from four samples TDP11.26.2, 64-74 cm, to TDP11.33.2, 74-84 cm.
TDP 12. South of Stakishari (Tanzania, Kilwa region), coordinates UTM 37L 560222 8981309; pteropods are available from 16 samples TDP12.11.4, 20-26 cm, to TDP12.29.2, 20-30 cm.
TDP 17. Stakishari (Tanzania, Kilwa region), coordinates UTM 37L 560539 8984483; pteropod specimens available from 36 samples 17.15.1, 50-63 cm, to 17.41.3, 0-15 cm.
Based on micropalaeontological event correlations the three sections were given “composite depths" to enable direct comparison of results between the three sites (Pearson et al., 2008). These composite depths are specified in the distribution tables following the systematical part.
The meters added are, for TDP 17 cores 1-35: 0 m; for TDP17 cores 36-42: 12 m; for TDP11 cores 1-28: 17 m, and for TDP11 cores 29-36: 20 m.
The Eocene Tanzanian shelf is thought to have been narrow, much like today, and the three boreholes are estimated to be approximately 50 km from the palaeo-shoreline (Kent et al., 1971; Nicholas et al., 2006, 2007). Based on the sedimentary facies and smaller benthic foraminiferal assemblage the palaeo-depth is estimated to be approximately 300-500 m, although it is possible it is deeper than this (Nicholas et al., 2006; 2007). The dominant sediment is dark green-grey clay with < 10% CaCO3.
The half-round core-samples of the hemi-pelagic clays, varying in length from 1.5 to 15 cm, were washed through a 63 μm sieve and the residues dried (Lear et al., 2008; Pearson et al., 2008; Wade and Pearson, 2008). Pteropods were then picked using a paintbrush under a binocular microscope.
The Kilwa Group is characterised by excellent preservation of its carbonate microfossils, this extends to the pteropods, which is related to the high clay content and shallow maximum burial depth (Pearson et al., 2001; Van Dongen et al., 2006; Bown et al., 2008). However, the condition of pteropod specimens varies from perfect shell preservation to shells filled with pyrite and specimens preserved as pyritic internal moulds with partly or completely dissolved aragonitic shell material. Local dissolution may be related to pyritisation. Some specimens are badly deteriorated and indeterminate. Specimens are housed in the fossil holoplanktic mollusc collection of Naturalis Biodiversity Center, Leiden (The Netherlands), registered with RGM-registration numbers.
Scanning Electron Microscopy
SEM micrographs were made by Renate Helwerda from uncoated specimens at 1.000 kv with a Jeol Field emission scanning electron microscope type: JSM-7600F at Naturalis Biodiversity Center, Leiden, The Netherlands.
Micro-Computed Tomography
Micro-computed tomography (Micro-CT or X-ray microtomography) scanning of the three holotypes (RGM 777415a, RGM 777428b and RGM 1007748b) was carried out by Dirk van der Marel, of Naturalis Biodiversity Center in Leiden, the Netherlands, using a Bruker SkyScan 1172 micro-CT scanner. 1601 projections of 3 -15 sec exposure were collected with a 2000x1336 detector, using a source voltage of between 80 and 140 kV. No filtration was used, and the scan provided a reconstructed dataset with 0.8 - 1.4 μm voxels.
The pteropod shells were segmented in Avizo 9.0.0. and surface files exported as.wrl. The files were subsequently edited in Meshlab 1.3.3. Laplacian smoothing was applied and Quadric Edge collapse Decimation used, reducing the polygon count by approximately a factor of 10. Files were saved as.ply and u3d for viewing as three dimensional PDFs.
SYSTEMATICS (AWJ)
Phylum MOLLUSCA Linnaeus, 1758
Class GASTROPODA Cuvier, 1795
Subclass HETEROBRANCHIA Burmeister, 1837
Superorder PTEROPODA Cuvier, 1804
Order THECOSOMATA de Blainville, 1824
Suborder EUTHECOSOMATA Meisenheimer, 1905
Superfamily LIMACINOIDEA Gray, 1847
Family LIMACINIDAE Gray, 1847
Genus ALTASPIRATELLA Korobkov, 1966 (= Plotophysops Curry, 1982)
Type species. “Limacina elongatoides" [sic] (Aldrich), by original designation of Korobkov (1966, p. 74) = Physa elongatoidea Aldrich, 1887 = Altaspiratella elongatoidea (Aldrich, 1887) (Eocene, early Ypresian, Wilcox Group, Hatchetigbee Formation, Bashi Member; Zone NP 10).
Altaspiratella bearnensis (Curry, 1982)
Figure 2.1-2
*v 1982 Plotophysops bearnensis Curry, p. 40, pl. 1, figure 9a-c.
 v. (1986) Spiratella tutelina Curr. - Merle, p. 43 (non Curry).
v. (1986) Spiratella tutelina Curr. - Merle, p. 43 (non Curry).
v. 1990b Altaspiratella bearnensis (Curry 1981) - Janssen, p. 68.
? 1992 Altaspiratella bearnensis (Curry) - Hodgkinson et al., p. 13, pl. 1, figures 1-2.
. (1996) Altaspiratella bearnensis (Curry, 1981) - Kunz, p. 164, pl. 30, figures 1-3.
v. 2010 Altaspiratella bearnensis (Curry, 1982) - Cahuzac and Janssen, p. 24, pl. 2, figures 1-4, pl. 3, figure 1.
v. 2013 Altaspiratella bearnensis - King et al., p. 192, 193.
Material examined. TDP 12.28.1, 66-76 cm, depth 90.91-91.01 m below surface, composite depth 148.91-149.01 m; RGM 777374 (1 specimen, Figure 2.1-2, H = 1.36, W = 0.80 mm).
Description. Only available specimen with high conical shell, 1.7 times higher than wide, and apical angle of c. 40°. Four and a half slightly convex and comparatively high whorls, regularly increasing in diameter and separated by incised, oblique suture. Whorls attach below periphery of preceding whorl. Specimen incomplete, last whorl missing, in shell preservation, but filled with pyrite.
Discussion. Two closely resembling Altaspiratella species are currently known. Altaspiratella elongatoidea (Aldrich, 1887) occurred during the earliest Eocene (Ypresian, nannoplankton Zones NP 9 and 10) of the USA (Hodgkinson et al., 1992; Janssen et al., 2016); A. bearnensis (Curry, 1982), introduced from the Ypresian (zone NP 12/13) of SW France, is also known from the USA (with some doubt) in rocks of middle Eocene, Lutetian age (Zone NP 15). These two species differ only very slightly in the proportions of their early whorls. Of A. elongatoidea no specimens preserving apertural structures are known, so there might be differences in that respect as well.
In the single available specimen from Tanzania apertural structures are missing, and its apical whorls take a position in between the two known species mentioned above. This could be an indication that these two taxa represent a single, long-ranging species. However, as long as apertural structures cannot be compared it seems better to keep them apart and the one available Tanzanian specimen is here considered, for the time being, to be the youngest representative of the A. elongatoidea - A. bearnensis lineage. The specimen extends the vertical range to the Priabonian (Zones P 16-17, NP 19-20; Nicholas et al., 2006, figure 16).
Genus HELICONOIDES d’Orbigny, 1835
Type species. Atlanta inflata d’Orbigny, 1834, by subsequent designation of Herrmannsen (1846, p. 514) = Heliconoides inflatus (d’Orbigny, 1834) (Recent).
Heliconoides nikkieae sp. nov.
Figure 3.1-5, Figure 4
zoobank.org/ACDEDDCE-7069-4BBB-B984-EB04C789D9DE
 Type material. Holotype RGM 777415a (Figure 3.1); paratypes1-2, RGM 777415b-c (Figure 3.3, 3.5) from the type locality. Kilwa Group, Pande Formation (upper Eocene, Priabonian), Zones P 18 and NP 21; paratype 3, RGM 777 381 (Figure 3, Figure 4), from TDP 17.21.1, 9-20 cm, Kilwa Group, Pande Formation (early Oligocene, Rupelian), biozones P18 and NP 21.
Type material. Holotype RGM 777415a (Figure 3.1); paratypes1-2, RGM 777415b-c (Figure 3.3, 3.5) from the type locality. Kilwa Group, Pande Formation (upper Eocene, Priabonian), Zones P 18 and NP 21; paratype 3, RGM 777 381 (Figure 3, Figure 4), from TDP 17.21.1, 9-20 cm, Kilwa Group, Pande Formation (early Oligocene, Rupelian), biozones P18 and NP 21.
Additional specimens. TDP 11 (2 specimens), 12 (1 specimen) and 17 (49 specimens, some of which with a query because of poor preservation) (Table 1, Table 2, Table 3).
Type locality. Stakishari (Tanzania, Kilwa region), cored borehole TDP 17, sample 17.36.1, 10-25 cm, 104.00-104.10 m below surface, composite depth 116.00-116.10 m.
Etymology. Named after Nikkie Elert, the author’s second granddaughter. At age six she is, in many respects, more up-grown than many grown-ups. Heliconoides gender masculine (ICZN 1992, art. 30.1.4.4).
Diagnosis. Very small limacinid of 2.5 whorls in low-conical spiral, about as high as wide, with large aperture, apertural margin externally thickened and internally doubled in some specimens (Figure 3.2), basal margin with denticle. Apertural margin preceded by about 10 fine, margin parallel riblets.
Description. Strikingly small limacinid of 2.5 whorls, height/width-ratio variable between c. 1.19 (holotype H = 0.62, W = 0.52 mm, Figure 3.1) and 0.89 (e.g., H = 0.50, W = 0.56 mm, Figure 3.2), with depressed, low conical spire and distinct, incised suture. Shell surface smooth and shiny, growth lines invisible. Last whorl large, occupying almost entire shell height. Aperture relatively large, slightly higher than wide to almost circular, occupying 4/5th of entire shell height. Apertural margin externally thickened by narrow ridge, running all around margin, flexuous at base of shell, reaching umbilicus, internally (Figure 3.2) in some specimens. Protruding denticle on basal part of margin (Figure 3.3-4). Marginal thickening preceded by about 10 fine, margin parallel orthocline riblets. Shell base regularly rounded, umbilicus very narrow, c. 1/20th of shell diameter.
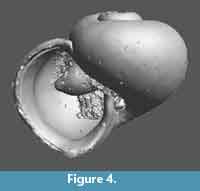 Discussion. In general shape the new species resembles somewhat the holotype of Limacina wechesensis Hodgkinson (1992, p. 21, pl. 5, figures 4-6) from the Lutetian of Texas, USA, but that species has one whorl more and reaches double the size of H. nikkieae, does not have the apertural structures of that species and its umbilicus is considerably wider.
Discussion. In general shape the new species resembles somewhat the holotype of Limacina wechesensis Hodgkinson (1992, p. 21, pl. 5, figures 4-6) from the Lutetian of Texas, USA, but that species has one whorl more and reaches double the size of H. nikkieae, does not have the apertural structures of that species and its umbilicus is considerably wider.
Size and apertural structures of the new species form a unique combination and cannot be compared to any limacinid currently known. The holotype was chosen from the sample with most specimens (13) of Priabonian age, but the species continues, in low numbers, well into the Rupelian part of the TDP 17 section.
Genus LIMACINA Bosc, 1817
Type species. Clio helicina Phipps, 1774 by monotypy = Limacina helicina (Phipps, 1774) (Recent).
Limacina robusta (Eames, 1952)
Figure 5, Figure 6
*1952 Aplexa robusta Eames, p. 152, pl. 6, figure 149.
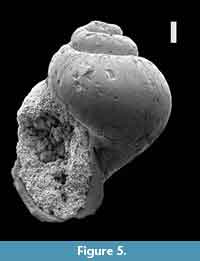 Type material. Holotype (Figure 6) in the Natural History Museum, London, NHMUK BM 68457, presented by the Burma Oil Co. Ltd., March 1950; Eames (1952) furthermore recorded 19 paratypes from the type locality.
Type material. Holotype (Figure 6) in the Natural History Museum, London, NHMUK BM 68457, presented by the Burma Oil Co. Ltd., March 1950; Eames (1952) furthermore recorded 19 paratypes from the type locality.
Type locality. Rahki Nala (Pakistan, western Punjab), 255’ above base, local zone 9. Lower Chocolate Clays; according to Afzal et al. (2009, p. 20) nowadays indicated as Kirthar Formation of late Lutetian - Priabonian age.
Material examined. TDP 11 (1 specimen), TDP 12 (1 specimen), TDP 17 (16 specimens, all from the Priabonian), see Table 1, Table 2, Table 3 for specification.
Description. Most available specimens are juveniles and most in pyritic internal mould preservation. Largest and best preserved specimen (Figure 5) higher than wide (H = 0.94, W = 0.72 mm) with four convex whorls rapidly increasing in diameter. Aperture large, oval, occupying more than half shell height. Base of shell regularly rounded with narrow umbilicus.
Discussion. Most of the available specimens are poorly preserved and juvenile, but the illustrated Tanzanian specimen (Figure 5) has a shell height of 0.94 mm, whereas the holotype of Limacina robusta has a shell height of 0.75 mm. Some of the smaller specimens have a somewhat wider apical angle than the illustrated specimen. In spite of these small differences the Tanzanian specimens are thought to represent the same species as the Pakistanian L. robusta, which has a comparable age and was described from a pre-eastern-Paratethys locality under influence of the Indian Ocean.
 Initially the Tanzanian specimens were thought to represent Limacina conica (von Koenen, 1892, p. 994, pl. 62, figures 5-6), a species described from the “early Oligocene" of two localities (Atzendorf, Unseburg) in the eastern part of Germany. However, specimens from Atzendorf (NP 21 interval), made available by Arnold Müller (Leipzig, Germany) differ in shape and reach far larger dimensions. Their apical angle is smaller, the whorls are more convex and the aperture remains smaller than half shell height.
Initially the Tanzanian specimens were thought to represent Limacina conica (von Koenen, 1892, p. 994, pl. 62, figures 5-6), a species described from the “early Oligocene" of two localities (Atzendorf, Unseburg) in the eastern part of Germany. However, specimens from Atzendorf (NP 21 interval), made available by Arnold Müller (Leipzig, Germany) differ in shape and reach far larger dimensions. Their apical angle is smaller, the whorls are more convex and the aperture remains smaller than half shell height.
Several species described by Hodgkinson et al. (1992), from the Paleogene of the United States also resemble the Tanzanian shells. Especially similar is Limacina smithvillensis Hodgkinson (in Hodgkinson et al., 1992, p. 19, pl. 3, figure 16) from the Lutetian of Texas. However, that species reaches a shell height of 1.5 mm, has less convex whorls, a wider apical angle, and its aperture occupies just half the shell height. Finally, that species is considerably older (Lutetian, NP 15) than the Priabonian (NP 21) specimens from Tanzania. At first glance also L. stenzeli Garvie (in Hodgkinson et al., 1992, p. 19, pl. 4, figure 1) from the NP13-14 interval (Ypresian) of Texas is similar. However, that species should be included in the genus Heliconoides, because of its reinforced apertural margin.
Limacina tanzaniaensis sp. nov.
Figure 7.1-4, Figure 8
zoobank.org/8C1AB54C-4643-4D30-9455-97C37B793992
Type material. Holotype (Figure 7.1) RGM 777428b; paratype 1 (Figure 7.2), RGM 777438; paratypes 2 and 3 (Figure 7.3-4) RGM 777416b-c. Kilwa Group, Pande Formation (Eocene, Priabonian, biozones P 18 and NP 21).
 Additional specimens. Fourteen specimens from TDP 17 (Table 3), all from the Priabonian part of the section. Most specimens poorly preserved as pyritic internal moulds.
Additional specimens. Fourteen specimens from TDP 17 (Table 3), all from the Priabonian part of the section. Most specimens poorly preserved as pyritic internal moulds.
Type locality. Stakishari (Tanzania, Kilwa region), cored borehole TDP 17, sample 38.2, 22-29 cm, 111.12-111.19 m below surface = 123.12-123.19 m composite depth.
 Diagnosis. Spherical limacinid of almost four whorls, apex flattened. Last whorl inflated, 95% of total shell height, aperture c. 75% of total shell height, margin simple, base of shell umbilicate.
Diagnosis. Spherical limacinid of almost four whorls, apex flattened. Last whorl inflated, 95% of total shell height, aperture c. 75% of total shell height, margin simple, base of shell umbilicate.
Description. Limacinid of spherical shape, measurements of holotype H = 0.92, W = 1.00 mm, consisting of 3.75 moderately convex whorls with convex tangent. First two whorls flattened, last whorl very large, inflated, occupying 95% of total shell height. Aperture large, about 75% of total shell height, attaching on (holotype, Figure 7.1) or slightly below (paratype 1, Figure 7.2) periphery of penultimate whorl. Apertural margin simple, semicircular, inner margin and columella invisible as all specimens are in pyritic internal mould preservation. Base of shell regularly rounded, umbilicus present, 20-25% of shell diameter.
Discussion. The available specimens demonstrate variability in height of the apex, in some the initial flattening continues to the third whorl, resulting in an only slightly raised apical shell part. There is some resemblance to the Ypresian species Limacina heatherae Hodgkinson in Hodgkinson et al. (1992, p. 17, pl. 2, figures 15-18). In that species, however, the whorls attach higher on the foregoing whorl, the aperture is narrower and its umbilicus smaller.
Limacina timi sp. nov.
Figure 9.1-4, Figure 10
zoobank.org/E5DC43EC-4F67-4173-890D-0145DEBEC0B6
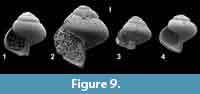 Type material. Holotype (Figure 9.1), RGM 1007748b; paratype 1 (Figure 9.2), RGM777408a, from TDP 17.34.1, 0-7 cm, 98.90-98.97 m below surface and composite depth; paratype 2 (Figure 9.3), RGM 777408b, TDP 17.34.1, 0-7 cm; paratype 3 (Figure 9.4), RGM 777414m TDP 17.36.1, 0-5 cm. Kilwa Group, Pande Formation (Eocene, Priabonian); planktic foraminefera zone P 16-17, calcareous nannoplankton zone NP 19-20.
Type material. Holotype (Figure 9.1), RGM 1007748b; paratype 1 (Figure 9.2), RGM777408a, from TDP 17.34.1, 0-7 cm, 98.90-98.97 m below surface and composite depth; paratype 2 (Figure 9.3), RGM 777408b, TDP 17.34.1, 0-7 cm; paratype 3 (Figure 9.4), RGM 777414m TDP 17.36.1, 0-5 cm. Kilwa Group, Pande Formation (Eocene, Priabonian); planktic foraminefera zone P 16-17, calcareous nannoplankton zone NP 19-20.
Additional specimens. Boreholes TDP 11, 12 and 17: 15 specimens (see Table 1 and Table 3).
Type locality. South of Stakishari, Tanzania, Kilwa Region, cored borehole TDP 12, coordinates UTM 37L 560222-8981309, sample TDP 12, sample 27.1, 35-45 cm = 89.60-89.70 m below surface = 147.60-147.70 m composite depth.
 Etymology. Named after Tim Janssen, the author’s six year old second grandson. For him life is nonstop fun. Keep it that way, my friend!
Etymology. Named after Tim Janssen, the author’s six year old second grandson. For him life is nonstop fun. Keep it that way, my friend!
Diagnosis. Small limacinid of 3.75 whorls, slightly higher than wide. Whorls rounded angular in juveniles, more regularly rounded in adults. Aperture lunate, somewhat higher than half shell height. Base imperforate.
Description. Holotype (H = 1.04, W = 1.00 mm) a small, regularly coiled, conical limacinid of 3.75 slightly convex whorls, separated by incised suture. Last whorl occupying 80% of shell height, slightly angular at periphery. Shell surface smooth, growth lines invisible. Aperture large, lunate, 65% of shell height, no apertural reinforcements present. Columellar side of aperture smooth, slightly concave internally, straight externally. Base of shell regularly convex, umbilicus absent in holotype, extremely small or absent in juveniles. Angularity of whorls more clearly developed in juvenile specimens, resembling a rounded keel, but covered by following whorls attaching at the place of angularity, almost disappeared in adults. Juvenile specimens wider than high, in some of these the keel is rather strong and the apical angle wider (RGM 777425).
Discussion. The angularity of especially juvenile specimens is not very clearly visible in the SEM images given here in Figure 9.3-4, but under light microscope, with illumination from left above it cannot be overlooked.
Limacina timi resembles somewhat the Ypresian species L. gormani (Curry, 1982) described from the Marnes de Gan Formation, of Gan, SW France (Curry, 1982; Cahuzac and Janssen, 2010). The same species or a closely related form was also described from the Stone City and Cook Mountain formations (Lutetian/Bartonian) of Texas, USA (Hodgkinson et al., 1992, p. 19, pl. 3, figures 14-15) as Limacina pygmaea (non Lamarck, 1805), in which also the juvenile whorls are angular and the base imperforate. The adult shell, however, reaches one and a half times the size of L. timi, has a different, more spherical shape and a distinctly wider apical angle. Subsutural crests (Cahuzac and Janssen, 2010, pl. 11, figure 2) as seen in L. gormani are not present in L. timi.
Adult specimens of Limacina timi also resemble Heliconoides nemoris (Curry, 1965), described from the Bartonian of the UK and also recorded from the Priabonian of Biarritz, SW France by Curry (1982) and Cahuzac and Janssen (2010). That species belongs to the genus Heliconoides because of its apertural reinforcements that are apparently absent in L. timi, but also its apex is flattened and juvenile specimens have no angular periphery. Limacina timi straddles the Eocene/Oligocene boundary.
Limacina sp. 1
Figure 11.1-2
Material examined. Five specimens, all from the same sample TDP 17.37.1, 0-13 cm; 106.90-107.03 m below surface, 118.90-119.03 composite depth; RGM 777440a-c (Table 3).
 Description. Low conical limacinid of 3.75 moderately convex whorls attaching on periphery of foregoing whorls. Aperture semicircular, slightly more than half shell height, Base regularly rounded, umbilicus c. 1/6th of shell diameter.
Description. Low conical limacinid of 3.75 moderately convex whorls attaching on periphery of foregoing whorls. Aperture semicircular, slightly more than half shell height, Base regularly rounded, umbilicus c. 1/6th of shell diameter.
Discussion. Three of the available specimens are juveniles in poor preservation. In specimen RGM 777440b (Figure 11.2) the first whorl is missing. The two more adult specimens show a striking resemblance with a limacinid illustrated by Lokno and Kumar (2008, figure 3-2) from the Upper Disang Formation (Bartonian - Priabonian) of the Phek District, south-central Nagaland (Assam - Arakan Basin), northeastern India, indicated by these authors as “Limacinidae type A." As these specimens were recorded from Indian Ocean Basin rocks of more or less similar age they could very well represent the same species as the Tanzanian ones. However, the material is insufficient for a reliable identification. Lokno and Kumar compared their specimens with “Limacina pygmaea," as illustrated by Hodgkinson et al. (1992, pl. 3, figures 14-15), correctly stating that that name “most certainly is incorrect," as Hodgkinson et al.’s species seems to be closely related to L. gormani (Curry, 1982) (see above) and not to Limacina pygmaea.
Limacina sp. 2
Figure 12.1-4
Material examined. One specimen from TDP 12 (Table 2); 4 poorly preserved and presumably juvenile specimens in pyritic internal mould preservation, from TDP 17 (Table 3).
Description. Limacinid of very low conical shape with almost flat, slightly raised, or slightly concave apical plane. Width of illustrated specimen 1.20 mm, height 0.86 mm. Whorls 3.75, regularly increasing in diameter. Aperture semicircular, occupying c. 80% or more of total shell height, reaching to far beyond base of preceding whorl. Apertural structures absent or not preserved. The apparently present groove along the apertural margin, as vsible in Figure 13.2-3, is considered to be caused by damage of the mould. Base perforated by umbilicus of 1/5th to 1/7th of shell diameter.
Discussion. Several limacinids with an almost planorboid shell shape are known from the Eocene-Oligocene interval in Europe, Asia and the USA. Some of these are characterised by having a slightly concave apical plane, or, in other cases, by an irregular development of early whorls. In the present specimens, however, the whorls are in a regular spiral and the apical plane is a bit raised, with the first 1.5 whorls flattened (Figure 12.3 ).
).
Very similar is a species from the early Oligocene of Japan, described as Limacina karasawai Ando (2011, p. 248, figures 3.1-2. This species was said to have three quarters of a whorl more than the Tanzanian specimen illustrated here, but we fail to see that from Ando’s photographs. Also closely similar is Limacina canadaensis Hodgkinson (in Hodgkinson et al., 1992, p. 16, pl. 2, figures 4-6), but its last whorl seems to be relatively lower. This species was collected from downhole contaminated cutting samples and could be anything between early Eocene and early Oligocene. An occurrence of similar age (earliest Oligocene) was described from the North Sea Basin and is also known from contemporaneous rocks (base of Viborg Formation) in Jylland, Denmark, as Limacina mariae Janssen (1989, p. 111, pl. 4, figures 2-5), but that species always has a concave apical spiral, has a somewhat wider umbilicus and reaches to over 2 mm shell width. Finally, as yet unpublished similar material is available from the Eocene - Oligocene interval in the NE United States and from the eastern part of Germany.
A reliable interpretation of all these forms depends on a larger material for comparisons and therefore the present specimens, apart from the illustrated specimen in poor condition, are left in open nomenclature.
Limacinidae indet.
Material examined. One specimen from TDP 12.24.3, 0-10 cm; 10 specimens from TDP 17 (see Table 3).
Description. These specimens are in internal pyritic mould preservation, more or less strongly deteriorated and cannot be identified any further.
Superfamily CAVOLINIOIDEA Gray, 1850 (1815) [= Hyalinea Rafinesque, 1815]
Family CRESEIDAE Rampal, 1973
Genus BOVICORNU Meyer, 1886
Type species. Bovicornu eocenense Meyer, 1886, by monotypy (early Oligocene, USA).
Discussion. Shortly after its introduction by Meyer (1886) the validity of the genus Bovicornu was denied by Dall (1892, p. 302), who considered both species introduced in that genus by Meyer (1886, 1887) to belong to the caecid (benthic) genus Meioceras Carpenter, 1858. Dall’s point of view was followed, with doubts, by Cossmann (1912, p. 154-155), but rejected by later authors (Collins, 1934, p. 212; van Winkle Palmer, 1947, p. 464; Hodgkinson et al., 1992, p. 24). Indeed, in Meioceras the shell wall is considerably thicker and its protoconch is utterly different, whereas in Bovicornu the shell is thin-walled and the larval parts agree with species of the pteropod genus Creseis. Zilch (1959, p. 49) followed Collins and included Bovicornu with a query in the cavoliniid pteropods, considering it a possible synonym of Euchilotheca Fischer, 1882, which is also a creseid genus, but its type species, E. succincta (Defrance, 1828), shows only very faint traces of spiralisation. We agree with Hodgkinson et al. (1992) that Bovicornu should be considered an independent genus in Creseidae.
Bovicornu species demonstrate a certain resemblance with Hameconia edmundi Janssen, 2008, described from the late Oligocene (Chattian) of SW France. In that species the bilaterally symmetrical shell has a curvature of c. 180°, but it is curved in one plain, not in a spatial spiral. The curvature is dorso-ventral, as is clear by the presence of lateral carinae. Its larval stage differs from Bovicornu in having separate protoconchs 1 and 2. Janssen and Maxwell (in Janssen, 1995, p. 164), Janssen (2008, p. 160) and Cahuzac and Janssen (2010, p. 111) included the genus Hameconia in the Sphaerocinidae family.
Bovicornu aff. eocenense Meyer, 1886
Figure 13.1-2
cf 1886 Bovicornu eocenense Meyer, p. 79, pl. 3, figure 12 (not figure 2).
c 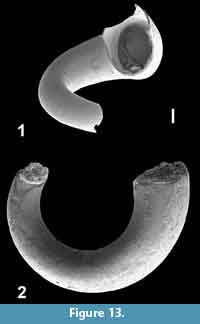 f 1892 Meioceras eocenense (Meyer) - Dall, p. 302.
f 1892 Meioceras eocenense (Meyer) - Dall, p. 302.
cf 1912 Bovicornu eocænense [sic] Meyer - Cossmann, p. 155.
cf 1934 Bovicornu eocenense Meyer - Collins, p. 212, pl.9, figure 3, pl. 13, figure 5.
cf 1959 Bovicornu eocenense O. Meyer - Zilch, p. 49, figure 164.
cf 1992 Bovicornu eocenense Meyer - Hodgkinson et al., p. 24, pl. 7, figures 9-10 (with additional synonymy).
Type material. Holotype (H = 2,8, W = 0,7 mm) United States Natural Museum (Smithsonian Institution) nr. 644596.
Type locality. Red Bluff, USGS locality 5264, Mississippi, USA (Red Bluff Clay; Oligocene, Rupelian, NP 21).
Material examined. Only fragments were found, 2 from TDP 11, 9 from TDP 12, and 11 from TDP 17 (see Table 1, Table 2, Table 3 for details).
Description. Spatially spiralised tube with free volutions. Transverse section of tube circular, no surface ornamentation or growth lines visible. Only smaller fragments are available among which no complete aperture or protoconch. Diameter of tube doubles in about half a volution.
Discussion. Two species of the genus Bovicornu are currently known and both are exclusively recorded from the USA. The older one of these, B. gracile Meyer, 1887 (p. 9, pl. 2, figure 17), of the Moodys Branch Formation, Texas, has an age of Bartonian (NP 17). The other species, B. eocenense Meyer, 1886 (p. 79, pl. 3, figure 12), from the Red Bluff Formation of Mississippi, USA occurred during the Priabonian and early Rupelian (NP19-21) (Hodgkinson et al., 1992, figure 3). In both species the shell is creseid, but instead of 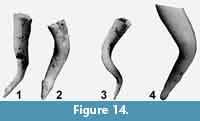 being straight or slightly curved, as in Creseis species, the tube demonstrates clear twisting in a wide spatial spiral that was said to be stronger in B. gracile. Hodgkinson et al. (1992), however, collected numerous specimens at both type localities and noted that in many specimens of both species the twisting is stronger and that the species cannot be distinguished on the degree of twisting. There is, however, a clear difference in protoconch morphology: an inflated bulb in B. eocenense and a more cylindrical shape in B. gracile. The holotypes of both species were re-illustrated in Hodgkinson et al. (1992, pl. 7, figures 9-10 and 11-12), reproduced herein as Figure 14.1-4.
being straight or slightly curved, as in Creseis species, the tube demonstrates clear twisting in a wide spatial spiral that was said to be stronger in B. gracile. Hodgkinson et al. (1992), however, collected numerous specimens at both type localities and noted that in many specimens of both species the twisting is stronger and that the species cannot be distinguished on the degree of twisting. There is, however, a clear difference in protoconch morphology: an inflated bulb in B. eocenense and a more cylindrical shape in B. gracile. The holotypes of both species were re-illustrated in Hodgkinson et al. (1992, pl. 7, figures 9-10 and 11-12), reproduced herein as Figure 14.1-4.
Although the few larger fragments from Tanzania seem to indicate a considerably stronger spirally twisted shell than in either of the holotypes it is preferred to indicate the Tanzanian species as related to the younger of the two American species and is indicated here as Bovicornu aff. eocenense. Once specimens preserving their protoconch become available this position might be revised.
RESULTS
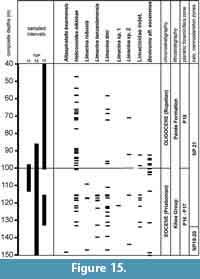
The number of specimens per species/per sample are specified in Table 1, Table 2, Table 3. Vertical ranges of the species in the three Tanzanian sections are shown together in Figure 15.
 Eight pteropod species are recognised, seven of them belonging to the Limacinidae and one (Bovicornu) to the Creseidae. Three of the limacinids are represented by a single or very few specimens only. Three species could only be identified in open nomenclature. Three of the limacinid species are described as new.
Eight pteropod species are recognised, seven of them belonging to the Limacinidae and one (Bovicornu) to the Creseidae. Three of the limacinids are represented by a single or very few specimens only. Three species could only be identified in open nomenclature. Three of the limacinid species are described as new.
The stratigraphic ranges are remarkable. The two most commonly occurring species, Heliconoides nikkieae and Limacina timi (both introduced herein), and the less frequently represented Limacina sp. 2 occur in comparable numbers both below and above the EOB and do not seem to be influenced by changing environmental conditions. Two other species, Limacina robusta and L. tanzaniaensis, on the contrary, disappear some 10 m below the EOB, at a level close to the extinction of the Turborotalia cerroazulensis- group of planktic foraminifera and the first oxygen isotope step (Step 1 of Pearson et al., 2008; Lear et al., 2008) although improved sampling could of course extend the ranges up to the EOB or beyond. One species, Altaspiratella bearnensis, was found as a single specimen only in one of the lowermost samples. Finally, a single species (Bovicornu aff. eocenense), is irregularly distributed in the Eocene part of the sections, but disappears in the basal 10 m of the Oligocene. No pteropods are found exclusively in the Oligocene part of the record.
DISCUSSION
The newly described taxa, as well as species recorded in open nomenclature, do not present clues that can be applied in long distance correlations. The present material from Tanzania furthermore originates from a very large area from which Paleogene pteropods have never been recorded previously. From the African continent only a late Eocene occurrence is known from Nigeria (Bende Ameki; Curry, 1965 and Naturalis collection) and some Eocene (Ypresian, Bartonian) material was collected from Egypt (Valley of the Kings) by the late Chris King (Naturalis Biodiversity Center collections, unpublished).
The presence of Altaspiratella bearnensis is quite unexpected. That species is only known from the Aquitaine Basin in SW France (type locality, Ypresian) and from the southern USA (Ypresian and Lutetian). Its presence in the Tanzanian Priabonian is surprising. The record, however, is based on a single incomplete specimen and better preserved material might prove that another species is involved.
The occurrence of Limacina robusta is interesting. That species was originally described from Pakistan, a locality under the influence of the Indian Ocean, which is equally the case for the Tanzanian material. If Limacina sp. 1 is indeed closely related to or even identical with “Limacinidae type A" as described by Lokho and Kumar (2008) from northern India it similarly represents an interesting palaeogeographical occurrence on the Indian Ocean.
The creseid species Bovicornu eocenense is to date exclusively known from the United States and its vertical distribution includes the Priabonian and the Rupelian transition, which is consistent with its range in the Tanzanian material. However, the Tanzanian material is poorly preserved and better specimens might lead to another specific interpretation.
When compared with previous isotope and microfossil data from the Tanzanian material it may be concluded that three pteropod species (Heliconoides nikkieae, Limacina timi and Limacina sp. 2) do not show clear response following the drastic climatic cooling at the EOB, but two species (Limacina robusta and L. tanzaniaensis) seem to disappear close to the cooling related step of the EOT, perhaps indicating a temperature sensitivity if a true disappearance. The last occurrence of these two taxa precedes that of the planktic foraminiferal family Hantkeninidae and the extinction level of the larger benthic foraminifera (Pearson et al., 2008; Wade and Pearson, 2008; Cotton and Pearson, 2011). However, it is similar to the last occurrence of the planktic foraminifera Turborotalia cerroazulensis Cole, 1928, and related species (Pearson et al., 2008; Wade and Pearson, 2008). In addition, the nannofossil assemblage has shown an increase in nutrient loving taxa close to the onset of the EOT (Dunkley Jones et al., 2008), suggesting nutrient increase in the water column as a potential contributing factor.
There are currently plans to re-core the EOB of Tanzania with wide diameter boreholes (Pearson and Hudson, 2014). Much larger samples and denser sampling may shed further light on the pteropod record across the EOB in the region.
CONCLUSION
Here we have shown a small but important insight into pteropod fauna from both an under-represented geological time and geographic region. Eight species were recognised, three of which were new. Two of the taxa show an apparent, at least, local extinction close to the first major cooling step of the EOT whilst the others seem unaffected, or have too few occurrences to tell. The occurrences of several taxa are surprising with Altaspiratella bearnensis, and Bovicornu only previously known from Europe and the USA, and from the USA, respectively. The occurrence of Limacina robusta, which is only previously known from Pakistan, suggests an Indo-Pacific connection. This therefore underlines the need for increased studies of Paleogene pteropod occurrences, particularly in regions that may be under-sampled but have good preservation potential. Furthermore it shows the importance of carrying out studies of less conventional micro-fossils on cores generally used for foraminiferal or nannoplankton studies.
ACKNOWLEDGEMENTS
The authors thank Y. Ando (Graduate School of Environmental Studies, Nagoya University, Furo-cho, Chikusa-ku, Nagoya, Japan), for discussion on Paleogene pteropod occurrences, and A. Müller (Institut für Geophysik und Geologie, Geologisch-Palaeontologische Sammlung, Leipzig, Germany) for providing pteropod material for comparisons. R. Helwerda of Naturalis Biodiversity Center, Leiden, NL, was kind enough to make the SEM images. D. van der Marel of the same institute made the CT-scannings. The Tanzanian Petroleum Development Corporation and COSTECH are acknowledged for their support of the TDP. Two anonymous reviewers are thanked for critically reading the manuscript and for their helpful comments.
REFERENCES
Afzal, J., Williams, M., and Aldridge, R.J. 2009. Revised stratigraphy of the lower Cenozoic succession of the Greater Indus Basin in Pakistan. Journal of Micropalaeontology 28:7-23.
Aldrich, T.H. 1887. Notes on Tertiary fossils, with descriptions of new species. The Journal of the Cincinnati Society of Natural History, 10(2):78-83.
Ando, Y. 2011. Oligocene pteropods (Gastropoda: Thecosomata) from the Kishima Formation, Saga Prefecture, southwest Japan. Revista Mexicana de Ciencias Geológicas 28(2):245-253.
Bednaršek, N. and Ohman, M.D. 2015. Changes in pteropod distributions and shell dissolution across a frontal system in the California Current System. Marine Ecology Progress Series, 523:93-103. doi: 10.3354/meps11199
Bodiselitsch, B., Montanari, A., Koeberl, C., and Coccioni, R. 2004, Delayed climate change in the late Eocene caused by multiple impacts: High-resolution geochemical studies at Massignano, Italy. Earth and Planetary Science Letters 223:283-302. doi: 10.1016/j.epsl.2004.04.028.
Bosc, [L.A.G.]. 1817. Limacine. Nouveau Dictionnaire d’Histoire Naturelle, 18:42.
Bown, P.R., Dunkley Jones, T., Lees, J.A., Randell, R.D., Mizzi, J.A., Pearson, P.N., Coxall, H.K., Young, J.R., Nicholas, C.J., Karega, A., Singano, J., and Wade, B.S. 2008. A Paleogene calcareous microfosil Konservat-Lagerstätte from the Kilwa Group of coastal Tanzania. Geological Society of America Bulletin 120(1/2):3-12. doi: 10.1130/B26261.1
Burmeister, H. 1837. Handbuch der Naturgeschichte zum Gebrauch bei Vorlesungen; zweite Abtheilung., Zoologie. T.C.F. Enslin, Berlin.
Cahuzac, B. and Janssen, A.W. 2010. Eocene to Miocene holoplanktonic Mollusca (Gastropoda) of the Aquitaine Basin, southwest France. Scripta Geologica 141:1-193.
Cande, S.C. and Kent, D.V. 1995. Revised calibration of the geomagnetic polarity timescale for the late Cretaceous and Cenozoic: Journal of Geophysical Research 100(B4):6093-6095.
Carpenter, P.P. 1858. First steps towards a monograph of the Cæcidæ, a family of rostriferous Gasteropoda. Proceedings of the Zoological Society of London 26:413-443.
Cole, W.S. 1928. A foraminiferal fauna from the Chapapote Formation in Mexico. Bulletins of American Paleontology 14:3-32.
Collins, R.L. 1934. A monograph of the American Tertiary pteropod mollusks. Johns Hopkins University Studes in Geology 11:137-234.
Cossmann, M. 1912. Essays de Paléoconchologie Comparée, 9. Paris (Cossmann and Lamarre).
Cotton, L.J. and Pearson, P.N. 2011. Extinction of larger benthic foraminifera at the Eocene/Oligocene boundary. Palaeogeography, Palaeoclimatology, Palaeoecology 311:281-296.
Coxall, H.K. and Pearson, P.N. 2007. The Eocene-Oligocene transition. In Williams, M., Haywood, A.M., Gregory, F.J., and Schmidt, D.N. (eds.), Deep-time perspectives on climate change: marrying the signal from computer models and biological proxies: The Micropalaeontological Society, London, Special publication: 351-387.
Curry, D. 1965. The English Palaeogene pteropods. Proceedings of the Malacological Society of London 36:357-371.
Curry, D. 1982. Ptéropodes éocènes de la tuilerie de Gan (Pyrénées-Atlantiques) et de quelques autres localités du SW de la France. Cahiers de Micropaléontologie 4 (1981):35-44.
Cuvier, G. 1795. Second mémoire sur l'organisation et les rapports des animaux à sang blanc, dans lequel on traite de la structure des mollusques et de leur division en ordre, lu à la Société d'Histoire Naturelle de Paris, le 11 prairial an troisième [30 May 1795]. Magazin Encyclopédique, ou Journal des Sciences, des Lettres et des Arts, 1:433-449.
Cuvier, G. 1804. Mémoire concernant l’animal de l’Hyale, un nouveau genre de mollusques nus, intermédiaire entre l’Hyale et le Clio, et l‘établissement d'un nouvel ordre dans la classe des mollusques. Annales du Muséum national d’Histoire Naturelle 4:223-234.
Dall, W.H. 1892. Contributions to the Tertiary fauna of Florida, with especial reference to the Miocene silex-beds of Tampa and the Pliocene beds of the Caloosehatchie River, 2. Transactions of the Wagner Free Institute of Science of Philadelphia 3:201-473.
de Blainville, H.M.D. 1824. Mollusques, Mollusca (Malacoz.). Dictionnaire des Sciences naturelles, 32:1-392.
Defrance, J.L.M. 1828. Vaginella. (Foss.). Dictionnaire des Sciences naturelles 56: 427.
Diester-Haass, L and Zahn, R. 2001. Paleoproductivity increase at the Eocene-Oligocene climatic transition: ODP/DSDP Sites 763 and 592. Palaeogeography, Palaeoclimatology, Palaeoecology. 172:153-170.
Di Martino, E., Taylor, P., Cotton, L.J., and Pearson, P.N. 2017. First bryozoan fauna from the Eocene/Oligocene transition in Tanzania. Journal of Systematic Palaeontology. http://dx.doi.org/10.1080/14772019.2017.1284163
Dunkley Jones, T., Bown, P.R., Pearson, P.N., Wade, B.S., Coxall, H.K., and Lear, C.H. 2008. Major shifts in calcareous phytoplankton assemblages through the Eocene‐Oligocene transition of Tanzania and their implications for low‐latitude primary production. Paleoceanography, 23. doi: 10.1029/2008PA001640
Eames, F.E. 1952. A contribution to the study of the rocks in western Pakistan and western India, C. The description of the Scaphopoda and Gastropoda from standard sections in the Rakhi Nala and Zinda Pir areas of the western Punjab and in the Kohat District. Philosophical Transactions of the Royal Society of London (B)631(236):1-168.
Fischer, P. 1880-1887. Manuel de conchyliologie et de paléontologie conchyliologique ou histoire naturelle des mollusques vivants et fossiles suivi d'un appendice sur les brachiopodes par D.P. Oehlert. Paris (Savy):1-12,i-xxiv,1-1369.
Gray, J.E. 1847. A list of the genera of recent Mollusca, their synonyms and types. Proceedings of the Zoological Society of London, 15(78):129-219.
Herrmannsen, A.N. 1846-1852. Indicis Generum Malacozoorum primordia. Nomina Subgenerum, Generum, Familiarum, Tribuum, Ordinum, Classium: adjectis Auctoribus, Temporibus, Locis Systematicis atque Literariis, Etymus, Synonymis. Praetermittuntur Cirripedia, Tunicatae et Rhizopoda, 1 (1846-1847), 2 (1847-1849), supplement (1852). T. Fischer, Cassel.
Hodgkinson, K.A., Garvie, C.L., and Bé, A.W.H. 1992. Eocene euthecosomatous Pteropoda (Gastropoda) of the Gulf and eastern Coasts of North America. Bulletins of American Paleontology, 103(341):5-62.
Janssen, A.W. 1989. Some new pteropod species from the North Sea Basin Cainozoic (Mollusca: Gastropoda, Euthecosomata). Mededelingrn vsn de Werkgrorpvoor Tertiaire en Kwartaire Geologie 26:91-133.
Janssen, A.W. 1990. Long distance correlation of Cainozoic deposits by means of planktonic gastropods (pteropods); some examples of future possibilities. Tertiary Research, 11:65-72.
Janssen, A.W. 1995. Systematic revision of holoplanktonic Mollusca in the collections of the Dipartimento di Scienze della Terra at Torino, Italy. Monografie Museo Regionale di Scienze Naturali Torino 17:1-233.
Janssen, A.W. 2008. Notes on the systematics, morphology and biostratigraphy of fossil holoplanktonic Mollusca, 20. A new pteropod genus and species, Hameconia edmundi gen. nov. spec. nov. (Mollusca, Gastropoda, Sphaerocinidae), from the Late Oligocene of SW France. Basteria 72:159-163.
Janssen, A.W., Sessa, J.A., and Thomas. 2016. E. Pteropoda (Mollusca, Gastropoda, Thecosomata) from the Paleocene-Eocene Thermal Maximum (United States Atlantic Coastal Plain). Palaeontologia Electronica, 19.3.47A:1-26.
palaeo-electronica.org/content/2016/1662-pteropoda-from-the-usa-petm
Jimènez Berrocoso, À., Huber, B.T., MacLeod, K.G., Petrizzo, M.R., Lees, J.A., Wendler, I., Coxall, H., Mweneinda, A.K., Falzoni, F., Birch, H., Singano, J.M., Haynes, S., Cotton, L., Wendler, J., Bown, P.R., Robinson, S.A., and Gould, J. 2012. Lithostratigraphy, biostratigraphy and chemostratigraphy of Upper Cretaceous and Paleogene sediments from southern Tanzania: Tanzania Drilling Project Sites 27-35. Journal of African Earth Sciences 70: 36-57.
Jiménez Berrocoso, Á., Huber, B.T., MacLeod, K.G., Petrizzo, M.R., Lees, J.A., Wendler, I., Coxall, H., Mweneinda, A.K., Falzoni, A., Birch, H., Haynes, S.J., Bown, P.R., Robinson. S.A., and Singano, J.M. 2015. The Lindi Formation (upper Albian-Coniacian) and Tanzania Drilling Project Sites 36-40 (Lower Cretaceous to Paleogene): Lithostratigraphy, biostratigraphy and chemostratigraphy. Journal of African Earth Sciences 101:282-308.
Jiménez Berrocoso, Á., MacLeod, K.G., Huber, B.T., Lees, J.A., Wendler, I., Bown, P.R., Mweneinda, A.K., Londoño, C.I., and Singano, J.M. 2010. Lithostratigraphy, biostratigraphy and chemostratigraphy of Upper Cretaceous sediments from southern Tanzania: Tanzania drilling project sites 21-26. Journal of African Earth Sciences 57 (2010):47-69.
Kent, P.E., Hunt, J.A.,and Johnstone, D.W. 1971. The Geology and Geophysics of coastal Tanzania. Institute of Geological Sciences Geophysical Paper 6, HMSO, London.
King, C., Iakovleva, A., Steurbaut, E., Heilmann-Clausen, C., and Ward, D. 2013. The Aktulagay section, west Kazakhstan: a key site for northern mid-latitude early Eocene stratigraphy. Stratigraphy, 10:171-209.
Korobkov, I.A. 1966. Krylonogie (Mollusca Pteropoda) paleogenovykh otlozhenij juga S.S.S.R. Voprosy Paleontologii 5:71-92. (In Russian)
Kunz, A. 1996. Schalenmorphologische Merkmale der Ontogenesestadien ausgewählter Pteropoda (Gastropoda: Heterostropha). Hamburg, Diplom-Arbeit im Studienfach Geologie-Paläontologie an der Universität Hamburg. In Germany.
Lamarck, [J.B.P.A. de] 1805-1806. Mémoire sur les fossiles des environs de Paris, comprenant la détermination des espèces qui appartiennent aux animaux marins sans vertèbres, et dont la plupart sont figurées dans la collection des vélins du Muséum, 5. Annales du Muséum d’Histoire Naturelle de Paris 5:28-36, 91-98,179-188,237-245, 349-357.
Lear, C.H., Bailey, T.R.,Pearson, P.N., Coxall, H.K. and Rosentha, Y., 2008. Cooling and ice growth across the Eocene-Oligocene transition. Geology 36: 51-254. doi: 10.1130/G24584A.1
Linnaeus, C. 1758. Systema Naturae per Regna tria Naturae, secundum Classes, Ordines, Genera,S species, cum Characteribus, Differentiis, Synonymis, Locis, 1 (editio decima, reformata). Salvii, Holmiae.
Liu, Z., Tou, S., Zhao, Q., Huang, W. 2004. Deep-water Earliest Oligocene Glacial Maximum (EOGM) in the South Atlantic. Chinese Science Bulletin 49:2190-2197.
Lokho, K. and Kumar, K. 2008. Fossil pteropods (Thecosomata, holoplanktonic Mollusca) from the Eocene of Assam-Arakan Basin, northeastern India. Current Science 94:647-652.
Meisenheimer, J. 1905. Pteropoda. Wissenschaftliche Ergebnisse der Deutschen Tiefsee-Expedition auf dem Dampfer "Valdivia“1898-1899, 9(1):1-314.
Merle, D. 1986. Contribution à l'étude paléontologique du gisement cuisien de Gan (Pyrénées-Atlantiques): systématique, évolution et paléoécologie, 1-2. PhD thesis École Pratique des Hautes Études, Dijon, France.
Meyer, O. 1886. Contribution to the Eocene paleontology of Alabama and Mississippi, 2. In : Smith, E.A. (ed.). Geology of Alabama. Geological Survey of Alabama, Bulletin 1:61-85.
Meyer, O. 1887. Beitrag zur Kenntnis der Fauna des Alttertiärs von Mississippi und Alabama. Bericht über die Senckenbergische Naturforchende Gesellschaft (1886):3-22.
Nicholas, C.J., Pearson, P.N., Bown, P.N., Dunkley Jones, T., Huber, B.T., Karega, A., Lees, J.A., McMillan, I.K., O’Halloran, A., Singano, J.M., Wade, B.S. 2006. Stratigraphy and sedimentology of the Upper Cretaceous to Paleogene Kilwa Group, southern coastal Tanzania. Journal of African Earth Sciences 45:431-466.
Pearson, P.N., Ditchfield, P.W., Singano, J., Harcourt-Brown, K.G., Nicholas, C.J., Olsson, R.K., Shackleton, N.J. and Hall, M.A. 2001. Warm tropical sea surface temperatures in the Late Cretaceous and Eocene epochs. Nature 413:481-487.
Pearson, P.N., McMillan, I.K., Wade, B.S., Dunkley Jones, T., Coxall, H.K., Bown, P.R.and Lear, C.H. 2008. Extinction and environmental change across the Eocene-Oligocene. Geology 36:179-182. doi: 10.1130/G24308A.1., 279-317
Pearson, P.N., Nicholas, C.J., Singano, J.M., Bown, P.R., Coxall, H.K., van Dongen, B.E.,Huber, B.T., Karega, A., Lees, J.A., Msaky, E., Pancost, R.D., Pearson, M., Roberts,A.P. 2004. Paleogene and Cretaceous sediment cores from the Kilwa and Lindi areas of coastal Tanzania: Tanzania Drilling Project Sites 1-5. Journal of African Earth Sciences 39:25-62.
Pearson, P.N., Nicholas, C.J., Singano, J.M., Bown, P.R., Coxall, H.K., van Dongen, B.E., Huber, B.T., Karega, A., Lees, J.A., MacLeod, K., McMillan, I.K., Pancost, R.D., Pearson,M., Msaky, E. 2006. Further Paleogene and Cretaceous sediment cores from the Kilwa area of coastal Tanzania: Tanzania Drilling Project Sites 6-10. Journal of African Earth Sciences 45:279-317.
Pearson, P.N. and Wade, B. 2015. Systematic taxonomy of exceptionally well-preserved planktonic Foraminifera from the Eocene/Oligocene boundary of Tanzania. Cushman Foundation Special Publication 45:1-85.
Phipps, C.J. 1774. A Voyage towards the North Pole undertaken by his Majesty’s Command 1773. Bowyer, W. and Nichols, J., London.
Premoli Silva, I. and Jenkins, D.G. 1993. Decision on the Eocene-Oligocene boundary stratotype. Episodes 16:379-382.
Rampal, J. 1973. Phylogénie des ptéropodes thécosomes d'après la structure de la coquille et la morphologie du manteau. Comptes Rendus de l’Académie des Sciences de Paris 277(D):1345-1348.
Rafinesque, C.S. 1815. Analyse de la Nature ou Tableau de l'Univers et des Corps organisés. Rafinesque, Palerme.
Sherborn, C.D. and Griffin, F.J. 1934. On the dates of publication of the natural history portions of Alcide d’Orbigny’s “Voyage Amérique Méridionale”. Annals and Magazine of Natural History, (10)13(73):130-134.
Van Dongen, B.E., Talbot, H.M., Schouten, S., Pearson, P.N., and Pancost, R.D. 2006. Well preserved Palaeogene and Cretaceous biomarkers from the Kilwa area, Tanzania. Organic Geochemistry 37:39-557. doi:10.1016/j.orggeochem.2006.01.003
van Winkle Palmer, K. 1947. The Mollusca of the Jackson Eocene of the Mississippi Embayment (Sabine River to the Alabama River), 2. Univalves. Bulletins of American Paleontology 30:207-563.
von Koenen, A. 1892. Das norddeutsche Unter-Oligocän und seine Mollusken-Fauna, 4(1). Rissoidae-Littorinidae-Turbinidae-Haliotidae-Fissurellidae-Calyptraeidae-Patellidae; (2) Gastropoda Opisthobranchiata; (3)(1) Gastropoda Polyplacophora; (2) Scaphopoda; (3) Pteropoda; (4) Cephalopoda. Abhandlungen zur geologischen Specialkarte von Preussen und den Thüringischen Staaten 10:819-1004.
Wade, B.S. and Pearson, P.N. 2008. Planktonic foraminiferal turnover, diversity fluctuations and geochemical signals across the Eocene/Oligocene boundary in Tanzania. Marine Micropaleontology 68:244-255.
Wall-Palmer, D., Hart, M.B., Smart, C.W., Sparks, R.S., Friant, A.L., Boudon, G., Deplus, C., and Komorowski, J.C. 2012. Pteropods from the Caribbean Sea: variations in calcification as an indicator of past ocean carbonate saturation. Biogeosciences, 9:309-15.
Zilch, A. 1959. Gastropoda, 2. Euthyneura 1, xii + 200 pp. In Wenz, W. (ed.). Handbuch der Paläozoologie 6. Bornträger, Berlin-Nikolassee.
