|
||||
DISCUSSION AND CONCLUSIONS
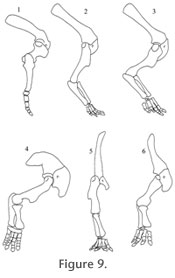
At maximum protraction, the long axis of the humerus was 43 degrees to the horizontal, at maximum retraction, about 24 degrees to the horizontal, with the capacity to elevate the humerus an additional 14 degrees during the protractive phase. Thus, throughout the step cycle, the humerus remained closer to the horizontal than vertical, and its range of motion (33 degrees if the protractive phase is included) was much more restricted than the 70 degrees predicted by Paul and Christiansen (2000, figure 4e). As pointed out by Johnson and Ostrom (1995), the elbow is strongly flexed throughout the entire step cycle, ranging from 87 to 114 degrees to the humerus. A sequence of images of the limb assembled to simulate the step cycle is provided in Figure 7.9. This simulation makes it clear why many traditional reconstructions of neoceratopsid forelimb stance include a strongly everted humerus. In these analyses, both the distal end of the epipodium and the metacarpus (and therefore manus) has been reconstructed as broad and flat (e.g., Johnson and Ostrom 1995, figure 12.7). Our specimen shows clearly that both were strongly arched, so that even when the humerus was strongly retracted, the distal extensor surface of the radius faced almost directly anteriorly. As noted by Paul and Christiansen (2000), footprint data (Lockley and Hunt 1995) indicates that the manus is directed anterolaterally (or to be more precise, the digits splay so that the first digit actually is directed somewhat medially, the second more or less directly anterior, and digits three through five are directed progressively more laterally) during the propulsive phase. If the humerus of our model is forced into the classic sprawling position, the manus is rotated so that all but the most lateral digits are directed medially at an improbable angle, and certainly into a position inconsistent with the trackway data. This locomotor pattern is highly derived relative to that of basal tetrapods, but shows some interesting parallels with the primitive sprawling stance. The humerus is maintained relatively close to the horizontal throughout the step cycle. The elbow is relatively inflexible and remains strongly flexed throughout the step cycle. Although Chasmosaurus can hardly be described as a "sprawler," the elbows are everted to a greater degree than in quadrupedal mammals. As a result, as the propulsive phase progresses, the weight gradually shifts to the radial side of the carpus as the ulnar side is lifted off the ground near the end of the phase. This is functionally analogous to the "metatarsal rotation" (Brinkman 1980) observed in the rear limb of extant lizards. This posture, which has no precise analogue in other tetrapods, and the resulting forces generated in the metacarpus and manus during propulsion, may explain the common occurrence of bone pathologies in the front foot of ceratopsids. The metacarpal and first phalanx of the medial (first) digit of the specimen described here are highly pathological (Figure 5). Significantly, in a specimen of Chasmosaurus belli (ROM 843), both left and right medial metacarpals and associated phalanges are similarly pathological. Such bilateral deformities are highly unlikely to be the result of acute trauma, but are consistent with chronic injury resulting from repetitive stress associated with compression of the medial side of the manus during walking (Rega and Holmes 2006), particularly in large, older individuals. Since it appears that primitive marginocephalians such as psittacosaurs, from which neoceratopsids presumably evolved, were bipedal (You and Dodson 2004), it is improbable that these features represent remnants of the primitive sprawling posture. Instead, it is likely that they evolved, de novo, within the group. Although it is now clear that Chasmosaurus, and probably all neoceratopsids, did not have sprawling front limbs, this simulation shows clearly that their elbows were distinctly everted, causing the front feet to track wider than the hind (Lockley and Hunt 1995). Extension of the elbow during the propulsive phase probably generated some transverse movement of the trunk, resulting in a slightly waddling gait. Whether these were adaptations to stabilize the huge heads of these dinosaurs, or simply reflect a secondary consequence of the evolution of the ability to assume a "secondary, wider forelimb gauge" (Paul and Christiansen 2000) is a matter for speculation. Although both lateral bending of the trunk and transverse movement created by extension of the everted elbows during the final stages of the propulsive phase would almost certainly have occurred, it is difficult to assess in a quantitative manner its effects on the step cycle of Chasmosaurus. The presence of ossified tendons would have stiffened the vertebral column, limiting lateral flexure and its effect on the length of the stride. Transverse movement of the trunk, produced by elbow extension, would have been modest given the restricted range of motion at this joint. As a result, we would predict that these excursions would be limited and would not affect fundamentally the model presented here. Inferred Stride Length – Does It Agree with Trackway Data?Although the position and orientation of the manus with respect to the sagittal plane in this simulation was based directly on the trackway data, the resulting estimate of stride was not. Rather, this was determined by the limitations placed on the range of motion by the morphology of the joints as preserved in the specimen. The stride of the trackway (Lockley and Hunt 1995) scaled to our half-scale model of C. irvinensis is approximately 600 mm. During the propulsive phase, the glenoid of our model moved forward a total of 257 mm (positions 1- 4). It is reasonable to assume that the propulsive phase would make up approximately half the distance between consecutive manus impressions, with the other half made by the opposite front limb while the manus swung forward to begin the next propulsive phase. This would give a stride of 514 mm in our model, only slightly less than the stride independently estimated from the scaled trackways derived from Lockley and Hunt (1995). The remainder of the stride could easily have been made up by modest excursion of the scapulocoracoid on the rib cage, a movement that almost certainly occurred during the step cycle (Paul and Christiensen 2000) and/or slight lateral undulation of the trunk. This provides independent corroboration for the step cycle hypothesized here. |
||||
|
|