|
||||
RECONSTRUCTION OF THE STEP CYCLEThe Neutral PositionWhen a quadruped takes a step, the front limb is extended,
the manus is placed on the substrate, and through contraction of muscles
originating from the trunk and inserting on the forelimb, the body is pulled
over the manus, while the manus remains stationary. This first half of the step
cycle is here referred to as the propulsive phase. Only at the end of the
propulsive phase is the manus raised off the ground. During the second half of
the step cycle, referred to here as the protractive phase, the manus is swung
forward again.
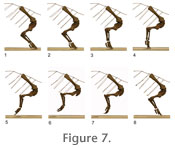
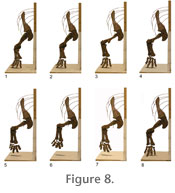
The analysis of the propulsive phase was begun by determining the most comfortable fit between the scapulocoracoid and the rib cage. In this position, the blade is 44 degrees to the horizontal, and the glenoid cavity faces primarily ventrally and posteriorly, and only slightly laterally. The limb was assembled with each element articulated at approximately the midpoint between the extremes of its joint excursions. This limb stance is hereafter referred to as the "neutral position" (Figure 6, Figure 7.2, Figure 8.2).
Viewed laterally, the long axis of the ulna is directed anteromedially at 50 degrees to the horizontal and viewed anteriorly, 2 degrees to the parasagittal. Viewed laterally, the long axis of the radius forms an angle of 55 degrees to the horizontal and, viewed anteriorly, 8 degrees to the parasagittal. The Propulsive PhaseThe beginning of the propulsive phase (Figure 7.1, Figure 8.1), here designated position 1, was established by moving the torso posteriorly from the "neutral position" while maintaining the manus in its print until at least one of the joints in the model could no longer be maintained in articulation (see "Materials and Methods"). The end of the propulsive phase (Figure 7.4, Figure 8.4), here designated position 4, was determined by moving the torso anteriorly over the stationary manus until at least one of the joints had been flexed or extended fully. Two additional positions, the neutral position (position 2 – Figure 7.2, Figure 8.2) and one midway between the neutral position and the end of the propulsive phase (position 3 – Figure 7.3, Figure 8.3), were also simulated. At each position, medial excursion was constrained by the medial tubercle and lateral excursion by glenoid shape. Each position was photographed in lateral (Figure 7) and anterior (Figure 8) views. The angles formed by the projections of the long axes of the humerus, ulna, and radius relative onto the horizontal (frontal) and parasagittal planes (Table 1) were measured directly from the photographs. Position 1 (Figure 7.1, Figure 8.1) represents the point at which the manus was first placed on the substrate at the beginning of the propulsive phase. The humerus was at its most protracted, with its long axis, viewed laterally, at 43 degrees to the horizontal and everted 29 degrees from the parasagittal (Table 1). Contact between the ventral surface of the humeral head and the ventral rim of the glenoid prevented further protraction. The elbow was flexed so that the ulna formed an angle of 89 degrees with the humerus. The sigmoid notch of the ulna articulated closely with the trochlea of the humerus, restricting rotation of the ulna about its long axis. The ulna and radius "crossed" proximally (Paul and Christiansen 2000), with the proximal head of the radius fitting into a depression on the extensor surface of the ulna immediately distal to the semilunar notch. The ulna formed angles of 43 and 3 degrees, and the radius, 50 and 17 degrees, with the horizontal and parasagittal, respectively. As the forelimb moved to position 2 (Figure 7.2, Figure 8.2), the torso of the model advanced 54 mm. The humerus retracted to a slightly more horizontal position, now forming an angle of 37 degrees to the horizontal plane and everted 27 degrees from the sagittal. To maintain the distance between the shoulder joint and the substrate, the elbow joint opened slightly (to 92 degrees) causing the epipodium to attain a more vertical orientation. To remain in articulation with the lateral condyle of the humerus, the ulna, and the manus, the proximal facet of the radius were required to migrate along the humeral condyle. The epipodium rotated closer to vertical. The humerus rotated slightly on its long axis to maintain the distal ulna and radial articulations with the manus. As the forelimb moved to position 3, the torso advanced an additional 120 mm (Figure 7.3, Figure 8.3). The humerus was more everted than at any other point in the propulsive phase, forming 32 degrees with the parasagittal. This orientation was necessary to maintain the proper relationship between the ulna, radius, and manus, otherwise, the radius moved too far medially, and would have disarticulated from the carpus. In order to maintain the distal end of the ulna in close enough proximity to the metacarpus, the humerus was rotated about its long axis, resulting in a more ventrally directed deltopectoral crest and more laterally facing external surface of its proximal expansion (Figure 7.3). The distal end of the humerus continued to swing dorsally, with the long axis of the humerus now forming an angle of 25 degrees with the horizontal. The elbow continued to open (105 degrees to the humerus), swinging the epipodium into a more vertical orientation. However, this movement was not sufficient to maintain a constant distance between the epipodium and metacarpus. To prevent the shoulder from moving ventrally, the metacarpals and proximal phalanges were rotated into a more vertical position. This is associated with increased dorsiflexion of the manus, but the relative positions of the distal ends of the digits did not change. As the forelimb moved from position 3 to position 4 (Figure 7.4, Figure 8.4), the torso advanced an additional 83 mm. Throughout this transition the distal end of the humerus rotated only one additional degree dorsally. Almost all of the forward propulsion during this phase of the cycle was derived from extension of the elbow joint. As the humero-ulnar articulation opened up completely, the distal end of the ulna swung laterally. In order to prevent the manus from being dragged laterally across the substrate, the distal end of the humerus was required to rotate medially. When position 4 was attained, the long axis of the humerus formed an angle of only 25 degrees with the parasagittal plane and 25 degrees from the parasagittal. The ulna became progressively more elevated, raising the lateral side of the manus off the substrate and shifting the weight toward the radial side of the manus. Position 4 represents the orientation of the forelimb just before the foot was lifted off the ground. The manus was at its point of greatest dorsiflexion to maintain proper relations between the epipodium and metacarpus without causing the glenoid to move ventrally. The elbow joint was maximally extended (114 degrees, and the epipodium is directed ventrally and slightly posteriorly. The lateral digits (four and five) had to be moved well off the substrate in order to maintain articulation between the metacarpus and ulna, shifting the weight to the preaxial side of the manus. As the manus pushed off the substrate, the first digit lost contact first. The second and third digits then appear to have lifted off the substrate simultaneously. In this simulated propulsive phase, the torso advanced a total of 257 mm. The Protractive PhaseThere is no way to determine the orientation of the manus during the protractive phase of the step cycle. However, as in the propulsive phase, the geometry of the joint surfaces sets some limits: the medial tubercle still must not interfere with the rib cage, and since known trackways show no evidence of knuckle drag, there must be an appropriate combination of distal humeral elevation and elbow flexion to allow the digits to remain clear of the substrate throughout the protractive phase. Within these admittedly incomplete parameters, we have attempted to reconstruct a plausible protractive phase (Figure 7.5-7.8, Figure 8.5-8.8). Once the digits have cleared the ground at the end of the propulsive phase, they are free to plantarflex. This, in combination with the posterior flexion of the wrist, is sufficient to keep the digits free of the substrate without requiring significant elevation of the distal end of the humerus during the initial stage of the protractive phase (Figure 7.5, Figure 8.5). However, in order to "set up" the manus for the remainder of the protractive phase, it was necessary to simultaneously elevate the distal end of the humerus to its maximum allowable travel in the glenoid, while simultaneously strongly flexing the elbow joint (Figure 7.7, Figure 8.7). This flexion begins early in the protractive phase (Figure 7.7). If the elbow remains flexed throughout most of the protractive phase, the distal end of the humerus can drop without creating knuckle drag, swinging the manus anteriorly (Figure 7.7-7.8, Figure 8.7-8.8). As this occurs, the manus swung through a broad lateral arc (Figure 8.7-8.8). Since the manus was not constrained by any contact with the substrate, there were no factors preventing such a movement; however, there is no evidence, other than that the model moved most naturally in this manner, that this would have actually occurred in the living animal. In the last portion of the protractive phase, elbow extension combines with continuing humeral depression to place the manus onto the substrate to begin the next step cycle (Figure 7.8, Figure 8.8). |
||||
|
|