|
|
THE RED HOT LEAF FLORAFerns
Order SCHIZAEALES
Frank, 1877
Distinguishing features and description. Width mostly uniform. Numerous, closely spaced secondary veins strongly decurrent on primary vein, diverging at low angles and dichotomizing multiple times. A lobe fragment with preserved length 3.8 cm, preserved width 1.6 cm. Comments. The specimen clearly matches the distinctive venation of Lygodium (Manchester and Zavada, 1987). Lygodium leaves are typically digitate-lobed, and this specimen is most likely a fragment of a single lobe. Berry (1914, 1916, 1924, 1930) reported several Lygodium species from various localities of the Gulf Coast Eocene, including L. kaulfussi which was widespread in the Eocene of North America, Europe, and Asia (Manchester and Zavada 1987). Manchester and Zavada (1987) concluded that all of Berry's species are indistinguishable from L. kaulfussi, which they considered the sole recognizable species of Lygodium in the Northern Hemisphere Eocene. The Lygodium considered here is also not distinguishable from L. kaulfussi and is accordingly placed in that species. Harrington (2003a) reported two schizaeaceous spore types from the Red Hot Truck Stop locality that could have been produced by Lygodium. Angiosperms
Order LAURALES
Perleb, 1826 Specimen. USNM 535048 (Figure 2.3; site 2). Morphotype RH12. Distinguishing features. Entire margin, acute basal pair of secondaries, and opposite percurrent tertiary veins that are approximately perpendicular to the primary vein. Description. Lamina elliptic, symmetrical, preserved length 8.3 cm, preserved width 4 cm; estimated l:w ratio 2.25:1; base convex, base angle acute, apex straight, apex angle acute; margin entire. Venation pinnate, primary vein straight. Secondary veins eucamptodromous, straight to slightly curved, angle from midvein 28°–65°, basal pair strongly acute, spacing irregular. Tertiary veins opposite percurrent, approximately perpendicular to the primary vein, or random reticulate. Fourth order veins random reticulate (photograph poorly). Comments. The combination of entire margin, acute basal secondaries, and tertiary veins oriented perpendicular to the midvein is widely prevalent among Lauraceae, to which we tentatively assign this single, poorly preserved fossil. MONOCOTYLEDONS
de Candolle, 1819
Distinguishing features and description. Two orders of parallel venation (A and B veins) with straight, curved, or sigmoidal oblique cross veins intersecting the parallel veins, both A and B. There are approximately 2–3 B veins between each pair of A veins. The cross veins run obliquely to the A and B veins and can be straight, curved, or sigmoidal. Comments. The parallel venation pattern and sigmoidal, oblique cross veins are characteristic of several monocot groups. Due to the lack of preservation details in these fragmented compression fossils, lower order taxonomic assignments cannot be made. Order MYRTALES
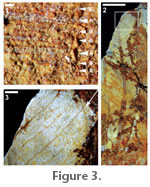
Reichenbach, 1828 Specimens. USNM 535051 (Figure 3.2, 3.3; site 1). Additional deposited specimens: four from site 1 and one from site 2. Morphotype RH02. Distinguishing features. Numerous thin, decurrent secondaries and intersecondaries that join a prominent intramarginal vein, and an entire margin. This distinctive architecture can be found in many genera of Myrtaceae (Carr et al. 1986). Description. Lamina oblong, symmetrical, narrow; preserved l:w ratio (l:w) ~5:1; precise l:w unknown; preserved length 4.7–7.0 cm, estimated to 8 cm; width 1.3–2.0 cm; base angle acute, apex not preserved; margin entire, fimbrial vein present. Venation pinnate; primary vein straight, thick basally. Secondary veins numerous (6–9 per cm), irregularly spaced, arising decurrently from primary vein at consistent 65° angles curving near intramarginal vein. Intersecondary veins strong, ~1 intersecondary per secondary with most intersecondaries reaching the intramarginal vein. Tertiaries not visible. Comments. Berry (e.g., Berry 1930) assigned various Eocene Gulf Coast leaves to myrtaceous genera such as Calyptranthes Swartz (1788), Eugenia Linnaeus (1753), and Myrcia de Candolle (1827), although these assignments are doubtful at the generic level (Manchester et al. 1998) and have never been revised. More recently, Dilcher and Lott (2005) tentatively assigned myrtaceous leaves from the middle Eocene Powers Clay Pit to Myrcia. Order FABALES
Bromhead, 1838
Distinguishing features. Conspicuously pulvinulate, marginally inserted petiolule; entire margin. Secondary veins diverge at low angle, extend distally without branching. Description. Lamina ovate to elliptical, symmetrical, usually narrow (preserved l:w 2:1 to 4:1; estimated l:w ~3:1; preserved length 1.0–4.5 cm, estimated 1.4–6 cm; width 0.7 cm to estimated 2.0 cm), base convex, base angle acute; apex generally straight, apex angle acute; margin entire, fimbrial vein present. Petiolule pulvinulate. Venation pinnate, primary vein straight. Secondary veins weak brochidodromous to eucamptodromous, extending distally, straight to slightly curved; ~3-4 secondaries/cm, arising from the midvein ~25°, spacing irregular, vein angles gradually decreasing and becoming more uniformly spaced proximally. Secondary veins closely spaced near the margin, becoming nearly parallel with margin, then joining superjacent secondary or diminishing. Intersecondaries weak to strong, not consistently present between each pair of secondaries. Tertiary veins opposite percurrent, perpendicular to the midvein between the closely spaced secondary veins near margin. Fourth order veins random reticulate; fifth order veins random reticulate to regular polygonal reticulate; freely ending veinlets (FEVs) unbranched. Comments. Legumes have been recognized in the Gulf Coast floras since Berry's time, and middle Eocene representatives have been extensively revised (Herendeen and Dilcher, 1990a, 1990b, 1990c; Herendeen, 1992). This morphotype is similar to Ormosia sp. reported by Herendeen (1992). Harrington (2003a) reported two pollen types from the Red Hot Truck Stop locality assignable to Fabaceae or Sapotaceae Durande (1782). Order FAGALES
Engler 1898
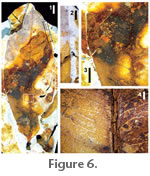
Distinguishing features. Base acute; margin irregularly serrate, tooth apices rounded, tooth apices weakly glandular. Secondary divergence angle ~65°; secondaries curve steeply distally near margin; intersecondaries weakly developed. Tertiary veins percurrent, sinuous, junctions with subjacent secondaries perpendicular to obtuse. Description. Lamina elliptic to ovate, symmetrical, narrow (preserved and estimated l:w ~3:1; preserved length 9.3 cm, estimated 12 cm; preserved width 3.6 cm, estimated 4 cm). Base angle acute, base shape unknown; apex angle and shape unknown; margin serrate, fimbrial vein present. Venation pinnate, primary vein straight. Secondary veins craspedodromous, straight near midvein, then steeply curving near the margin, 10–12 pairs, angle 60–75°, spacing irregular, angles smoothly decreasing proximally. Intersecondary veins weak; agrophic veins absent. Tertiary veins opposite percurrent, straight to sinuous; angle to primary obtuse, decreasing exmedially, junctions with subjacent secondaries perpendicular to obtuse. Fourth order veins random reticulate. Teeth small and in one order, weakly glandular, sinuses rounded, spacing irregular, about three teeth per centimeter. Comments. The venation of the single specimen is preserved only as faint impressions in the matrix and photographs poorly (Figure 5); nevertheless, it is surprisingly complete. In particular, the opposite percurrent, straight to sinuous tertiaries and their perpendicular to obtuse junctions with subjacent secondaries are clearly but faintly visible (Figure 5.2, 5.3). Despite the preservation, this fossil demonstrates the defining characteristics of Platycarya foliage (Wing and Hickey 1984; extant Platycarya may have simple or compound leaves depending on species). Pollen of this genus is also found in the same sediments (Frederiksen 1998; Harrington, 2003a). The specimen corresponds best to the Eocene Western Interior taxa P. americana (Hickey 1977) and P. castaneopsis (Lesquereux) Wing and Hickey (1984), whose leaves have many differences with living Platycarya but whose fruits and seeds are generically diagnostic (Wing and Hickey 1984). Due to preservation, one character found in Platycarya leaves, the ascending vein from sinus to superjacent tooth (Wing and Hickey 1984), was not observed. Order MALPIGHIALES
von Martius, 1835 Description. This morphogenus is erected for leaves having certain characteristics found in the extant genus Rhabdophyllum van Tieghem (1902a). The diagnostic characters of Rhabdophyllites, in combination, are: a toothed margin; closely spaced, thin, and numerous secondary veins of the "Clusia" type discussed by Gentry (1993) and Keller (2004); and distinct intersecondary veins. Discussion. This morphogenus is assigned to Ochnaceae, which is the sole extant family having the distinctive combination of toothed margin and "Clusia" venation. The morphogenus designation and –ites suffix (Greek, "having the nature of": Brown 1956) conservatively allow placement in the family and highlight the general similarity to extant Rhabdophyllum (tropical Africa) without categorization to any extant genus. Type species. Rhabdophyllites diapyros Rhabdophyllites diapyros sp. nov.
Etymology. Greek, "red-hot, fiery" (Brown 1956). Named in memory of the Red Hot Truck Stop, a popular local landmark, 1955–2000. Distinguishing features. Elliptic leaf with single primary vein. Secondary veins numerous, thin, closely spaced (3-8 mm apart along primary), eucamptodromous; intersecondary veins well developed, closely spaced (2-3 mm apart along primary), but both secondaries and intersecondaries vary and intergrade in distance covered from midvein to margin before branching or deflection; thus, individual intersecondaries are difficult to distinguish from secondaries as well as tertiaries. Tertiary veins random reticulate; teeth cryptic to conspicuous, best developed towards the apex.
Comments. The distinctive, closely spaced, parallel, thin secondaries and intersecondaries mark the "Clusia" venation type (Gentry 1993; Keller 2004) found within many Clusiaceae Lindley (1836b) and certain genera of Sapotaceae, such as Manilkara Adanson (1763), Micropholis (Grisebach) Pierre (1891), and many others; Vochysiaceae Saint-Hilaire (1820), including Qualea Aublet (1775) and Ruizterania Marcano-Berti (1969); and Ochnaceae de Candolle (1811a) such as Blastemanthus Planchon (1846), Elvasia de Candolle (1811b), Euthemis Jack (1820), Philacra Dwyer (1944), Rhabdophyllum van Tieghem (1902a), Schuurmansiella Hallier (1913), and Tyleria Gleason (1931). All four groups are characteristically tropical in distribution (e.g., Heywood 1993). Leaves in Ochnaceae are often toothed, whereas the three other families listed uniformly have untoothed leaves (Gentry 1993). An extremely rare exception is the Sapotaceae species Chrysophyllum imperiale (Linden) Bentham and Hooker (1876), which has vein architecture completely unlike this fossil. Thus, the combination of "Clusia" venation and a toothed margin diagnoses the fossils to Ochnaceae, the only other possibility being an extinct toothed genus from one of the families mentioned above.
The fossil record of other Malpighiales groups, as well as molecular clock divergence estimates, suggests a Cretaceous origin for all Malpighiales families (Davis et al. 2005). Fossil fruits that resemble the extant genus Ochna Linnaeus (1753) have recently been found in the Late Paleocene of North Dakota and are currently being described (Pigg and DeVore 2005). However, to our knowledge, the distinctive specimens reported here are the most reliable fossil leaves of Ochnaceae, a family with approximately 30 extant genera and 500 species, of mostly trees and shrubs, having tropical South America as its major center of diversity within a pantropical distribution (Heywood 1993). Berry described middle Eocene leaves from Tennessee (Berry 1930) and Río Negro, Argentina (Berry 1938; dated in Wilf et al. 2005) as species of Ouratea Aublet (1775), but neither bears close resemblance to the extant genus. Although there are scattered reports of fossil Ochnaceae pollen in the literature (Gruas-Cavagnetto 1976; Barbin 1992), these must be considered unreliable, until more detailed analyses are done, because the extant pollen morphology is unspecialized and thus very difficult to distinguish from similar groups (e.g., Muller 1969). There appear to be no reliable pollen records of Ochnaceae older than Holocene (e.g., Muller 1981). Order SAPINDALES
Dumortier, 1829
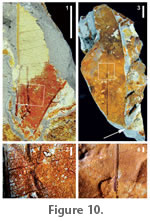
Distinguishing features. Large, compound, irregular teeth; craspedodromous, nearly cladodromous secondary veins branch conspicuously and terminate into both tooth orders and into sinuses; thin but prominent intersecondary veins; secondaries and intersecondaries irregularly angled. Comments. Conspicuously forking to cladodromous secondary veins, irregularly angled secondaries and intersecondaries, large and irregular to compound teeth, and termination of secondary veins or branches thereof in pointed tooth apices and sinuses comprise the characteristic architecture for toothed Rhus leaflets (e.g., Wolfe and Wehr 1987; Martínez-Millán and Cevallos-Ferriz 2005). A few other genera in Anacardiaceae show some components of this venation syndrome, including Loxopterygium Bentham and Hooker (1862) and Schmaltzia Desvaux ex Small (1841), but it is best expressed and most widely prevalent in Rhus. Harrington (2003a, 2003b) reported possible Anacardiaceae pollen from the Red Hot Truck Stop section. Description. Lamina narrow (size data not available due to fragmentation), shape and angles of apex and base unknown; margin serrate, fimbrial vein present. Venation pinnate, primary vein straight. Secondary veins craspedodromous to cladodromous, 6–8 pairs preserved, thickness and angle irregular. Intersecondaries thin, ~parallel to secondaries, angle irregular. Tertiary veins random reticulate. Teeth in two orders, ~four teeth/cm; teeth large, glandular; sinuses angular; tooth apices pointed; secondary vein branches terminate in sinuses and compose principal veins. Incertae sedis, morphotype RH15 Specimens. USNM 535064 (Figure 9.3; site 1). Distinguishing features. Strongly asymmetrical base; irregularly spaced secondary veins that diverge off primary vein with a different divergence angle on either side of the primary vein. Description. Lamina (probable leaflet fragment), oblong, wide (preserved l:w ratio 1.6:1, estimated l:w ratio ~2:1), base asymmetrical, decurrent, angle acute, petiole attachment marginal. Margin entire, fimbrial vein present. Venation pinnate; primary vein thick, curved basally; basal veins three. Secondary veins craspedodromous, 8–10 pairs spaced irregularly, arising from midvein decurrently at ~55°, curving distally near margin. Intersecondaries weak. Tertiary veins random reticulate; fourth order veins regular polygonal reticulate; fifth order veins random reticulate, FEVs unbranched where preserved. Incertae sedis, morphotype RH16 Specimen. USNM 535065 (Figure 9.4; site 1). Distinguishing features. Thick primary vein with strongly decurrent secondary veins and prominent intersecondaries. Description. Leaf fragment, base and apex characteristics unknown, preserved lamina width ~2.5 cm; observed margin entire. Venation pinnate, primary vein stout, tapering distally; secondary veins eucamptodromous or craspedodromous, curved, thin, numerous (4 pairs in small exposed area), decurrent on midvein with divergence 45–60°. Intersecondaries present, conspicuous. Tertiary veins random reticulate; higher order venation not preserved. Incertae sedis, morphotype RH04
Distinguishing features. Two primary veins (probably three or more originally) diverging from each other and connected by interior secondary veins. Leaf possibly palmately lobed. High order venation and areolation unusually well-preserved (Figure 9.2) for this flora, suggesting well-lignified veins. Description. Fragment; lamina size, base, apex, and margin features unknown. Primary venation apparently actinodromous or palinactinodromous; preserved secondary venation interior, spacing decreasing basally, perpendicular to primaries. Tertiary veins random reticulate, spacing close (1.5–3 mm). Fourth and fifth order veins random reticulate; areolation moderately developed, five or more sided; FEVs unbranched or one-branched. Incertae sedis, morphotype RH05 Specimen. USNM 535059 (Figure 10.3, 10.4; site 1). Distinguishing features. Rounded sinus (Figure 9.3, arrow), which indicates this is a lobed leaf, and thick tertiary veins arising perpendicular to the primary vein that join the subjacent secondary vein. The marked difference in secondary vein divergence angle on either side of the lobe primary (Figure 9.4) indicates that this is a left lateral lobe. From same locality as RH04 but lacking the distinctive impressed areolation (Figure 9.2). Description. Preserved lobe narrow, lobe sinus rounded; margin entire, fimbrial vein present. Venation presumably actinodromous or palinactinodromous. Secondary veins eucamptodromous. Tertiaries arising from midvein thick, divergence from primary perpendicular, then deflecting to subjacent secondary; tertiary veins arising from secondary veins. Higher-order venation present but indeterminate. Incertae sedis, morphotype RH08
Distinguishing features. Ovate blade, entire margin, and eucamptodromous or weak brochidodromous, irregularly spaced secondary veins, diverging from the midvein at ~40°, and then curving towards the margin with few, weak intersecondary veins. Description. Lamina elliptic to ovate, symmetrical, narrow (preserved and estimated l:w ratio ~3:1; preserved length 1.2–6.5 cm, estimated 3.5–7 cm; preserved width 1.0–3.0 cm, estimated 1.3–3.2 cm); margin entire, fimbrial vein present. Venation pinnate; primary vein straight. Secondary veins eucamptodromous to weakly brochidodromous, ~2–3 secondaries per cm, divergence angle ~40°, gently to sharply curving off primary vein and curving apically near margin; spacing irregular. Intersecondary veins few to absent. Tertiary veins random reticulate, oriented nearly perpendicular to primary vein near margin where secondaries are closely spaced; fourth order veins random reticulate. Incertae sedis, morphotype RH14 Specimen. USNM 535063 (Figure 11.2, 11.3; site 2). Additional deposited specimens: five from site 2. Distinguishing features: Poorly preserved morphotype with eucamptodromous or weakly brochidodromous secondary veins, which diverge from the midvein at 45°. Although very generalized in appearance, RH14 is distinct from other morphotypes presented here in the combination of eucamptodromous or weakly brochidodromous secondary venation, entire margin, and a high length:width ratio. Description. Lamina elliptic, probably symmetrical prior to preservation, narrow (preserved l:w ratio ~5:1, estimated l:w ratio ~3.5:1; preserved length 5.7–10 cm; preserved width 2.5–4.5 cm); base angle acute, apex rounded, apex angle obtuse; margin entire, fimbrial vein present. Venation pinnate; primary vein straight, width decreases distally. Secondary veins eucamptodromous or weakly brochidodromous, straight-slightly curved, divergence angle ~45°. Higher order venation not preserved. Incertae sedis, morphotype RH10
Distinguishing features. Acute base and apex angles, and secondary veins diverging off the midvein at ~65°. Description. Lamina elliptic, narrow (preserved l:w ratio ~3.3:1, estimated l:w ratio ~3.5:1; preserved length 12.5–13.5 cm; width 3.5 cm); base apparently asymmetrical, base angle acute; apex straight and acute; margin entire, fimbrial vein present. Venation pinnate; primary vein width decreasing distally. Secondary veins eucamptodromous to weakly brochidodromous, slightly curved, divergence angle 65°; agrophic veins absent; intersecondary veins weak if present. Tertiary veins random reticulate. Incertae sedis, morphotype RH11 Specimen. USNM 535062 (Figure 12.2, 12.3; site 2). Distinguishing features. Large, blunt, widely spaced, semitriangular teeth, concave on both flanks; craspedodromous secondary veins lose gauge as they enter teeth. Description. Lamina apparently elliptic; margin serrate, fimbrial vein absent. Preserved length 5.6 cm (estimated 9 cm); preserved width 2.8 cm (estimated 3 cm). Venation pinnate; secondary veins craspedodromous, spacing irregular, angles uniform. Intersecondary veins absent. Tertiary veins opposite percurrent. Teeth of one order, spacing ~1 tooth/cm, large, concave on both flanks, sinuses angular, apices glandular; secondaries lose gauge, or decrease in width, entering teeth. Incertae sedis, morphotype RH18 Specimen. USNM 535067 (Figure 12.4; site 1). Distinguishing features. Basally eucamptodromous and apically brochidodromous secondary veins that turn upwards sharply along the margin towards the apex of the leaf, and well-developed opposite percurrent tertiary veins that change course with the secondary veins. Description. Lamina elliptic, symmetrical; narrow (preserved l:w ratio 2.4:1; estimated l:w ratio ~2.1:1; preserved length 6 cm, estimated 7 cm; preserved width 2.2, estimated 3 cm), base and apex angle acute but shape indeterminate; margin entire. Venation pinnate, primary vein slightly sinuous. Secondary veins brochidodromous apically, eucamptodromous basally, extend straight towards the margin, turning steeply near margin, 9 pairs, divergence angle ~50°, spacing regular. Tertiary veins opposite percurrent; angle to midvein obtuse admedially, perpendicular to midvein near margin. Higher order venation present but poorly preserved. Incertae sedis, morphotype RH17 Specimen. USNM 535066 (Figure 12.5, 12.6; site 2). Distinguishing features. Entire margin, acute base. Weakly brochidodromous, thin secondary veins diverging from the midvein at ~55°; random reticulate tertiary veins. Description. Lamina apparently elliptic, narrow (preserved l:w ratio 2.5:1, estimated l:w ratio 2.25:1; preserved length 2.5 cm, estimated 4.5 cm; preserved width 1.5, estimated 2 cm); base cuneate, base angle acute; margin entire, fimbrial vein present; petiole attachment marginal. Venation pinnate; secondary veins weakly brochidodromous, looping close to margin, 5 pairs observed, divergence angle ~55°, spacing increasing basally. Intersecondary veins well developed; tertiary veins random reticulate. Incertae sedis, cuticle morphotype 1
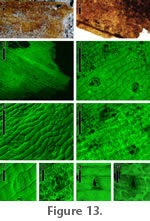
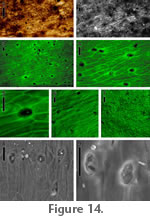
Distinguishing features. The strap-shaped cuticle fragments are often preserved yellow to orange. Cells are visible under stereoscope (Figure 13.1), with both epidermis and hypodermis preserved. The epidermal surface is striated, and stomata are present. Description. Strap-shaped cuticle, ~ 5.5 mm width. Epidermal cells elongate (width 14-26 m), with surface striations, cells more elongate along leaf margin. Hypodermis thick-walled, cells less elongate than those of epidermis. Stomatal areas sunken in relation to surrounding epidermis, guard cell pairs (24-28 m in polar length) oriented obliquely to long axis of surrounding epidermal cells; subsidiary cell arrangement paracytic. Venation absent. Comments. Paracytic stomata and strap-shaped leaves are suggestive of Monocots. This cuticle morphotype may have been produced by the same source species as monocot compression morphotype RH03 described above, but without preserved venation, this is impossible to establish. Incertae sedis, cuticle morphotype 2
Distinguishing features. The cuticle is preserved in fragments of variable shape that are dark brown. Cells are difficult to distinguish under stereoscope, and only epidermis is preserved. The epidermis lacks surface striations and bears densely distributed pore-like structures with raised rims. Description. Cuticle fragments, epidermal cells elongate, surface striations lacking, surface rough to smooth under ESEM. Hypodermis and stomata not present. Epidermis bears densely distributed pore-like structures with raised rims, pores filled with debris (visible under ESEM). Comments. The pores consistently lack remnants of guard cells or trichome bases. Thus, the pores appear to be a true feature of the cuticle, rather than sites of detached or degraded stomatal areas. Open pores of varying morphology are produced by thalloid liverworts (Ligrone et al. 2007). |
|