|
|
|
DISCUSSIONImplications of the Experiment for Pneumatic Support of a Chain BeamAs can be seen from the experiment, already one ventral pneumatic body on a chain beam has a bracing and support effect even under low pressure. An additional bracing element at the base of the chain beam, represented in the experiment by a ventral compressive bracing element), is crucial to hold the entire beam. The higher the internal pressure of the balloon, the better is the support effect. A ventral pneumatic body increases the load capacity of a chain beam. Even a slight increase of pressure results in an over-proportional increase of load capacity, which is aligned with longitudinal torque problems. With two ventral pneumatic systems these torque problems vanish, however, the increase of the support effect and loading capacity is not doubled at the same time. Laterally placed pneumatic bodies have no support effect to a chain beam. The introduction of a dorsal pneumatic body additionally to the pair of ventral tubes increases the support effect further and decreases the sagging of the distal part of the chain beam. However, a single dorsal pneumatic system needs to be connected with the pivot segments. The support quality of a segmented beam with a dorsal pneumatic system depends on the quality of its fixation at the base: stabilizing the pivot point of the chain beam increases the load capacity of the dorsally pneumatically stabilized chain beam. In all arrangements of pneumatic systems tested here, pneumatic stabilization occurs already at low pressures and increases with increasing pressure. The mobility of the segments against each other is restricted, if the pneumatic system is firmly fixed to the segments, and the load capacity increases at the same time. Segmentation of the pneumatic tube decreases the support effect drastically even with proximal suspension. The intersegmental constriction of the balloons reduces the support effect to zero. There is some support effect, if the constrictions between the pneumatic subunits lie level with the middle of the chain beam segments. In general, the increase of pneumatic segmentation results in a decrease of the support effect. A unilateral increase of pressure results in curvature of the chain beam towards the low-pressure side. The pressure difference determines the degree of curvature. The support effect of the investigated pneumatic structures is also influenced by other bracing elements. Adding proximoventral intersegmental wedges in all experiments demonstrates that ventral compressive structures provide an essential bracing effect in a chain beam. One single dorsal pneumatic system only works together with a dorsal tensile bracing element attached to the pivot, which suspends the pneumatically braced chain beam as a whole. Ventral compressive and dorsal tensile structures both work independently, but a maximum support effect is achieved when both are present at the same time. The Reconstructed Distribution of Pneumatic Diverticula in the Neck of Sauropods and Their Influence on a Hypothesized Pneumatic SupportBased on osteological correlates of vertebral pneumaticity, the reconstruction of pneumatic diverticula in sauropods can only mirror their minimum hypothetical extension, determined from their bony boundaries on the vertebrae. The relationship between vertebral pneumaticity and its osteological correlates in extant birds shows that the diverticula can be much larger than suggested by osteology (Müller 1908; O'Connor 2006). For sauropods, it cannot be excluded that pneumatic diverticula formed similar large segmented air chambers extending all along the neck, and in between the extremes of a minimum and a maximum hypothetical extension of the pneumatic diverticula, everything is possible. However, the reconstructed extension of pneumatic diverticula determines the possibility of the diverticula system to contribute to neck support in sauropods. Laminae-bounded deep fossae and/or foramina on the surface of the cervical vertebrae of sauropods are similar to the characteristic traces that pneumatic diverticula leave on the cervical vertebrae of extant avians (Britt 1993; Wedel et al. 2000; O'Connor 2001; Wedel 2005; O'Connor 2006). Although it has been proposed that deep fossae in the presacral vertebrae of sauropods would represent muscle attachment pits (e.g., Bonaparte 1999), there are no muscles in extant Sauria that produce such deep insertion pits along the vertebral column (Britt 1993; O'Connor 2006). The general absence of Sharpey's fibres in these fossae, which would be indicative for muscle or tendon insertions, also argues for a pneumatic origin of these structures. The presacral vertebrae of many sauropods are also hollowed out by a cavity system like in extant birds (Britt 1993; Wedel et al. 2000; Wedel 2003a; Upchurch et al. 2004; O'Connor 2006). In birds, these cavities are the result of bone resorption caused by bone-penetrating pneumatic epithelium (Bremer 1940; Witmer 1997). The presacral intravertebral cavity system of sauropods is most probably the result of a similar process (Janensch 1947; Britt 1993; Wedel et al. 2000; Wedel 2003a). The assumption that the intravertebral cavities could have been filled with blood or lymph is highly unlikely, because pneumatic epithelium is the only material known among vertebrates that can actively produce such cavities. Furthermore, a liquid infill would reasonably increase the weight of the vertebrae and thus the load of the neck. Vertebral pneumaticity therefore is the most plausible explanation for the intravertebral system of caverns in sauropods. The direct comparison with extant birds suggests that the combination of fossae and foramina connecting to an intravertebral cavity system is indicative for vertebral pneumaticity in Eu- and Neosauropoda.
1.
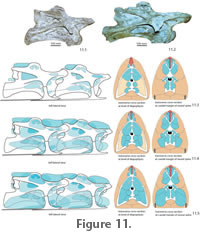
2. The Intermediate
Expansion Model. The pneumatic
diverticula in this reconstruction extended along the cervical vertebrae
according to their osteological correlates. However, the diverticula were larger
than their osteological correlates, overlapping each other distally and bridging
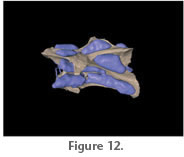
the interarticular gaps (Figure 11.4,
Figure 12). The segmented pneumatic diverticula would have been fixed to the vertebral segments by connective
tissue, the lateral bony arc formed by the tuberculum and diapophysis of the
cervical rib, the laterally adjacent axial musculature, as well as the
interspinal septum and supraspinal ligaments (Figure 3,
Figure 11).
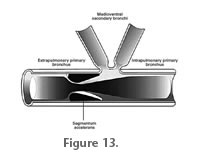
3. The Maximum Expansion Model. For this model, the pneumatic diverticula in the neck of sauropods are reconstructed to be bundled in large, densely packed diverticula systems, forming large hose-like canales intertransversarii and supraspinal diverticula units (Figure 11.5). Osteological structures on the cervical vertebrae of sauropods indicate the contact area of the diverticula with the vertebral surface, as well as the presence of interspinal septa and ligaments and muscles, which could, together with connective tissue, have wrapped expanded diverticula systems at least partially (see above). Both the presence of large hose-like canales intertransversarii and supraspinal diverticula units can only be reconstructed indirectly by comparison with extant birds. Thus, the reconstruction of the pneumatic diverticula to the maximum extent must remain partly speculative, hence not more likely or unlikely than the previous two options. The maximum extension reconstruction would result in a significant support effect of the pneumatic diverticula system as demonstrated in the experiment. This pneumatic support would have allowed a partial reduction of the tendinomuscular bracing system, and together with weight reduction by the pneumatic diverticula, a minimum of muscle mass would be needed to move the neck. The experiment also showed that the dorsal pneumatic diverticula would have needed to extend into the trunk to provide stable abutment for this diverticula group. Respiration, Regulation Mechanisms and Pressures in the Cervical Pneumatic System of SauropodsIf the pneumatic diverticula in the neck of sauropods are reconstructed according to an intermediate or maximum expansion model, they could have contributed to neck support. Such a supporting pneumatic system would require a minimum amount of pressure in the air sacs and diverticula, which supposedly would have been already necessary to warrant the open lumen. Pressure regulating mechanisms could have been present, but mechanically would not be necessary for support and bracing. None of these mechanisms leave osteological traces, and their presence can only be discussed in comparison with birds as the only extant vertebrates possessing pulmonary pneumaticity. The lung air sac system of modern birds differs from the respiratory system of other amniotes by its small incompressible lungs connected to nine air sacs. The avian lung is positioned ventrally adjacent to the vertebral column of the trunk and the arcades of the vertebral ribs. The pulmonary air sacs, positioned cranially and caudally to the lungs, change in volume and ventilate the lungs back-to-front in a manner of bellows. The gas exchange is conducted in the unidirectionally ventilated lung pipes, but not in the pulmonary air sacs (Duncker 1971; Schmidt-Nielsen 1971; Maina 2005). In modern birds, controlled pressure and volume changes of the cranial (interclavicular, cervical and cranial thoracic) and caudal (caudal thoracic and abdominal) pulmonary air sacs and between them are crucial for the unidirectional ventilation of the lungs (Brackenbury 1971; Duncker 1971; Schmidt-Nielsen 1971; Duncker 1974; Brown et al. 1995; Boggs et al. 1998; Fedde 1998). Starting from a normal pressure of zero, pressure differences comprise a positive and negative spectrum around 0.05 kPa (=0.0005 bar), and the pressure drop from the caudal thoracic air sac to the cranial intrapulmonary bronchus has been determined to be about 4.9 kPa (= 0.049bar) (Banzett et al. 1991; Brown et al. 1995). Pressure and volume of the different pulmonary air sacs are also influenced by locomotion (Schorger 1947; Kooyman 1971; Tucker 1972; Banzett et al. 1992; Boggs et al. 1997; Boggs et al. 1998; Boggs et al. 2001). Pressure differences between cranial and caudal air sacs during locomotion in different birds reach values between 0.12 kPa and 0.5 kPa (=0.0012 to 0.005 bar), pressure changes within the air sacs are around 0.35 kPa (= 0.0035 bar) (Boggs et al. 2001). Other muscle activities, singing or defecation lead to further pressure changes in the air sacs (Gaunt et al. 1973; Gaunt et al. 1976; Smith 1977; Brackenbury 1978; Suthers et al. 2002; Beckers et al. 2003; Franz and Goller 2003; Mindlin et al. 2003).
Even if the respiratory system of sauropods did incorporate unidirectional ventilation of the lungs by pulmonary air sacs, the rigidity of the sauropod trunk dictates completely different pumping mechanisms of these air sacs from that of birds. Together with the sheer dimensions of the respiratory apparatus, this would necessarily have required active regulation mechanisms, pressure and air flow control combined with a much slower breathing frequency. The presence of air sacs both in sauropods and in birds, the hypothesized similarities in the respiration mechanism between both groups, and the close phylogenetic relationship between sauropods and birds make bird-like regulation mechanisms for air sac volume and pressure possible for sauropods. The large variety in regulation mechanisms of birds shows that such a bird-like respiration system bears a great potential for the development of structures regulating air flow and pressure properties of the air sacs. However, it remains unknown, which of these is crucial for the maintenance of extended air sac systems in general. What can be reconstructed is that in sauropods at least the contraction of some muscles inserting ventrally on the scapulocoracoid and running to the neck, e.g., m. levator scapulae and m. capitisternalis, and of muscles running from the sternum to the skull, like m. episternobranchiotendineus and m. episternobranchialis would have resulted in a pressure increase of the cranial pulmonary air sacs. It is also possible that sauropods had developed their own, unique regulation and ventilation mechanisms, such as dynamic valves or constrictor muscles. Sauropod lungs were most likely as in birds positioned ventrally adjacent to the vertebral column of the trunk and the vertebral ribs and possessed large, sac-like dilatations (pulmonary air sacs) in the cranial and probably also caudal trunk region (Britt 1993, 1997; Perry and Reuter 1999; Wedel et al. 2000; Wedel 2003b). The pneumatic diverticula in the cervical vertebral column of sauropods can be most plausibly explained as derivates of pulmonary air sacs in the cranial trunk region (Wedel et al. 2000; Wedel 2003b, 2005). Assuming similar histology and properties of the pneumatic epithelium of sauropods as in birds, the air sacs and their continuation of cervical pneumatic diverticula were non-respiratory and connected to the respiratory apparatus as dead space. The presence of extraordinarily long necks in sauropods is combined with exceedingly long tracheas. If a bidirectional breathing mechanism would be assumed, much of the tracheal air pillar would just be moved forth and back without any respiration effect. This dead air pumping could have been avoided with a unidirectional ventilation of the lungs similar to birds. The incorporation of pulmonary air sacs into the respiration process for ventilation of the lungs would have made a cross-current gas exchange possible (Daniels and Pratt 1992; Perry and Sander 2004). Physiological considerations demonstrate that a bird-like model of a cross-current gas exchange, with a unidirectional ventilation of the lungs, would have yielded an optimum breathing efficiency in sauropods compared to other breathing mechanisms (Daniels and Pratt 1992; Perry and Sander 2004). No data are available about pressures in the cervical pneumatic system of extant birds. The pressure in the air sacs of birds is very low, and varies only slightly during breathing (e.g., Boggs et al. 1997; Boggs et al. 1998). Akersen and Trost (2004) described experimental inflation of air sacs and pneumatic diverticula of the neck of extant birds, which revealed a support effect already at 0.6 +/-0.2 psi (= 4.137 +/-1.379 kPa). These values are similar to the pressure differences between parts of the lung air sac system in extant birds (see above), making it possible that similar pressures can be reached in the pneumatic diverticula system. Even if stabilizing pressures of around 4 kPa would have occurred in their cervical diverticula system, regulation of the pressure or the possibility to disconnect the cervical system from the pulmonary air sacs were necessary preconditions for pneumatic stabilization in sauropods. Because the air sac system of sauropods was most likely non-respiratory, the cervical system could be easily disconnected from the rest of the respiratory apparatus without a negative influence to the respiration process. The small size of the connections between the cervical pneumatic diverticula of sauropods must have made gas exchange between these diverticula slow, and certainly decreased the influence of inspiration- or expiration-induced pressure and volume changes of the pulmonary air sacs. It is therefore likely, that even without a possibility to shut the cervical diverticula system off, the pressure within the cervical pneumatic system remained more or less constant. During movements of the sauropod neck (i.e. flexion, extension and lateral movements), pneumatic diverticula on the flexed part of the neck would be compressed. This could result in a partial deflation of these diverticula, with air exhausting into the opposite diverticula systems through pneumatic commissures, and/or into the cranial pulmonary air sacs. Alternatively, the small size of the pneumatic ducts could hinder such partial air flow, in which case air in these pneumatic diverticula would be compressed. In any case, the presumably small pressures in the air sac system of the neck would not lead to a failure of the system or the membrane. If regulation mechanisms were present in the diverticula system, this would make the system adjustable, leveling the compression effect during neck movement. The Bracing System of Sauropod Necks and Pneumatic SupportWith a few exceptions (i.e., Brachytrachelopan, Rauhut et al. 2005), sauropod dinosaurs possessed enormously long necks, which could exceed 9 m in length (e.g., Brachiosaurus, Janensch 1950; or Mamenchisaurus, Young et al. 1972). With the enormous body sizes of sauropods, such long necks needed to be sustained by a combination of different bracing elements. Simplified to the mechanical model of a segmented cantilever (see above), the model sauropod neck would have to accommodate tensile forces dorsally and compression forces ventrally by dorsal and ventral bracing structures. Sauropods show deep insertion scars for supraspinal and elastic ligaments (Tsuihiji 2004). The supraspinal ligaments connected the apices of each neural spine with each other, and were separated into two sheets in the case of bifurcate neural spines evidenced by cranial and caudal ridges on the neural spines. The elastic ligament system of sauropods extended craniocaudally connecting the bases of successive neural spines (Schwarz et al. 2007). The large dorsal ligament systems of sauropods most probably transmitted dorsal tensile forces into the vertebral chain, stabilizing the neck from the dorsal side. The ligament system was most probably supported by the tendinous system of the strongly segmented epaxial musculature (Wedel and Sanders 2002; Schwarz et al. 2007). The cervical ribs of most sauropods overlap ventral to the caudal following intervertebral gap and were connected with each other by short, intercostal ligament fibers (Schwarz et al. 2007). The cervical ribs can form overlapping bundles of up to five costal bodies like in Mamenchisaurus. Less overlapping is seen in Brachiosaurus or Camarasaurus. In other cases, the cervical ribs are approximately 1.5 times as long as the vertebrae with segment-to-segment overlap and have a stout, dorsomedially slightly concave costal body (e.g., in Malawisaurus, Jobaria or Shunosaurus). Along the cervical ribs, the compressive forces acting in longitudinal direction on the costal bodies probably transformed into a tensile load of the intercostal ligaments (Martin et al. 1998). Therefore, the elastic fixation of the overlapping costal bodies by ligaments allowed the rib apparatus along the neck to accommodate a part of the loading of the neck beam. Among extant animals, such a ventral costal bracing of the neck occurs in crocodilians only (Frey 1988b; Salisbury and Frey 2001). Based on the mechanisms described above, the neck of sauropods was suspended by dorsal ligaments in combination with strongly segmented epaxial muscles. Ventrally, it was supported by the paired parallel line of cervical ribs. This dorsoventral combination of bracing elements corresponds mechanically to a segmented truss, where the dorsal, tensile element is conducted along the hard parts of the beam and thereby separated from the compression member. As in other vertebrates, the cervical vertebrae of sauropods were also stabilized by the zygapophyseal articulations especially against torsion (see above). The presence of these bracing structures in sauropods shows that the pneumatic diverticula system in the neck of sauropods could only have contributed to neck support as a mechanically coherent part of the bracing system. If so, the diverticula system must have been compatible with the other bracing structures. Like in the experiment, the dorsal spinal ligaments and cervical ribs could have functioned as basal pivot supports, buffered by pneumatic diverticula as compression transmitting structures. There is a correlation between the development of the cervical pneumatic system and the other bracing elements in specific sauropod neck constructions, which will be discussed here. Dual Pneumatic Systems. Sauropods for which a dual pneumatic system in the neck is reconstructed (a pair of intertransversal diverticula units and a segmented chain of dorsal diverticula) have generally cervical ribs, which are longer than the adjacent vertebral segment. In Mamenchisaurus and Omeisaurus, the extremely low neural spines indicate a drastic reduction of the dorsal tensile elements due to the low spinal lever, which is combined with an extreme elongation of the cervical ribs, which span as much as seven vertebral segments (Young et al. 1972; Young and Zhao 1972; Russell and Zheng 1993; Tang et al. 2001). Probably, the twin chain of cervical ribs could in these cases support the neck beam nearly alone. If pneumatic diverticula in sauropod necks have a support function, the well-developed paired ventral pneumatic diverticula unit could have contributed to the ventral cervical support, increasing the efficiency of the intercostal bracing system due to pneumatic stabilizing effects. In contrast, the dorsal pneumatic structures in these sauropods were diminutive and mechanically could not contribute to the bracing of the neck. Regardless of the role of pneumatic diverticula, the focusing on ventral bracing elements in these sauropods is on the cost of overall mobility of the neck, because the long cervical ribs could slide along each other only to a small amount. In other taxa with a dual pneumatic system (such as in Brachiosaurus, Camarasaurus, Malawisaurus, Jobaria or Shunosaurus), the presence of well-developed cervical neural spines and large insertion areas for dorsal ligaments indicate the existence of a dual ligamentocostal bracing. If pneumatic diverticula supported the neck of these sauropods, then the paired ventral pneumatic diverticula could have contributed to their cervical bracing system. Additionally, the bifurcation of the neural spines in the neck base and the cranial part of the thorax of these sauropods indicates the existence of pneumatic diverticula, which were transient from neck to trunk. Thus, these diverticula systems could have additionally reinforced the neck from dorsally, but only in context with the strong ligament system. The extension of the dorsal diverticula units into the trunk is consistent with the experimental data showing a need for an abutment of the dorsal diverticula system at the pivot plate. Tripartite Pneumatic System. Sauropods with a tripartite pneumatic system (a paired ventral intertransversal diverticula unit and a dorsal unit of pneumatic diverticula, which continues onto the trunk), such as dicraeosaurids, Apatosaurus, and Diplodocus, possess high neural spines indicating large dorsal ligaments (Tsuihiji 2004; Schwarz et al. 2007) and high epaxial muscles acting on long spinal levers. The diameter of the ligaments, however, was equal or less than in those taxa with a dual system of pneumatic diverticula like Camarasaurus. The cervical ribs do not reach more than the length of the adjacent vertebral segment and thus do not overlap. As a consequence, bracing of the neck must have been achieved mainly by the dorsal ligaments and the epaxial tendinomuscular systems. If it had a support function, the tripartite system of pneumatic diverticula would have reinforced the dorsal bracing structures. The pneumatic diverticula lying between the metapophyses of the bifurcate neural spines in the trunk were very likely connected to the vertebrae of neck and trunk. Due to this and its transient nature from neck to body, the cranial half of the dorsal pneumatic system of the trunk might have been part of the cervical bracing system, providing stable abutment for the dorsal unit of cervical diverticula at the trunk. A reduction of the cervical rib apparatus in these sauropods increased the overall mobility of the neck beam in comparison to other sauropods. An additionally pneumatic support might have triggered the reduction of the cervical costal chain. |
|