|
|
|
SYSTEMATIC PALEONTOLOGY
DIAPSIDA Osborn 1903
sensu
Laurin 1991 Type Species. Vancleavea campi Long and Murry, 1995, by monotypy.
Vancleavea
campi
Long and Murry, 1995
2002
Vancleavea campi;
Hunt, Heckert, Lucas and Downs, p. 269, fig. 2. Holotype. PEFO 2427, fragmentary postcranial skeleton. Referred Specimens from Petrified Forest National Park. PEFO 31202, left femur, from locality PFV 215 (Zuni Well Mound), Petrified Forest Member. PEFO 33978, partial skeleton, from locality PFV 231 (The Giving Site), Petrified Forest Member. PEFO 34035, partial skeleton, from locality PFV 215 (Zuni Well Mound), Petrified Forest Member. PEFO 34224, distal end of femur, and PEFO 34231, sacral centrum, both from an unknown locality in the Blue Mesa Member. UCMP 178050, caudal vertebra, from UCMP locality V82251 (PFV 97 - Saurian Valley), Sonsela Member. Locality information for these specimens is available at Petrified Forest National Park for qualified researchers. See Hunt et al. (2002) for further referred specimens outside Petrified Forest National Park. Revised Diagnosis. Non-archosaurian archosauriform distinguished by the following autapomorphies: 1) cervical centra with a length/width ratio of greater than 1.5; 2) mid-cervical centra with a convex posterior articular face; 3) dorsal vertebral centra with weakly developed double ventral keels; 4) caudal centra that are subrectangular in ventral view with almost no waisting and possessing extremely well-developed sharp ventral keels; 5) a carapace of numerous subrounded osteoderms, each possessing a pronounced median keel and anterior projection; and 6) ilium with a tall anteriorly directed blade that does not extend anterior to the pubic peduncle, which superficially resembles the ilium of drepanosaurs. Distinguishing synapomorphies include 1) loss of the "terminal" femoral head and ventral ridge system ( sensu Carroll 1988) as in basal archosauromorphs and basal archosauriforms; 2) cervical vertebral diapophyses and dorsal vertebral parapophyses divided by the neurocentral suture as in Euparkeria and Erythrosuchus; and 3) presence of a prominent, medially directed femoral head as in Euparkeria, Turfanosuchus, Doswellia, Chanaresuchus, and archosaurs. Distribution. Blue Mesa, Sonsela, and Petrified Forest Members, Chinle Formation, Arizona; Siltstone Member, Chinle Formation, New Mexico; Los Esteros Member, Santa Rosa Formation, Dockum Group, New Mexico; Bull Canyon Formation, Dockum Group, New Mexico; Redonda Formation, Dockum Group, New Mexico; and Tecovas Formation, Dockum Group, Texas (Long and Murry 1995; Hunt et al. 2002). Age. Late Triassic, Norian (Muttoni et al. 2004; Furin et al. 2006; Irmis and Mundil 2008). Description
Vertebrae
An isolated, well-preserved cervical centrum from PEFO 34035 (Figures 4.4-4.7) differs from the vertebrae described above in possessing well-developed parapophyses (Figures 4.4-4.5). The anteroventral surface of the centrum is slightly beveled (Figure 4.7) and the dorsal surface is broad, forming a shelf (the ventral portion of the diapophysis) where it meets the ventral surface of the neural arch (Figure 4.6). The lateral surfaces of the centrum are concave, and the ventral surface possesses a sharp anteroposteriorly directed keel (Figure 4.5).
The neural arch of the dorsal vertebra (Figures 4.11-4.12) is tall (27 mm), roughly equal to the height of the centrum. The prezygopophyses project anterodorsally just past the anterior articular face of the centrum. The articular facets of the prezygopophyses are angled about 45 degrees medially, similar to the cervical series; however, the postzygopophyses do not project as far dorsally as those of the cervical series. The short (18 mm) transverse processes are anterolaterally expanded, project posterolaterally, and are situated posteriorly on the neural arch Figures 4.11-4.12). The articular surface (divided diapophysis) is broad. Distinct parapophyses and diapophyses in the cervical and dorsal vertebrae demonstrate that the cervical and dorsal ribs of Vancleavea were dolichocephalous. Sacral. According to Long and Murry (1995) two sacral centra are preserved in the holotype (PEFO 2427); sacral centra are also known from PEFO 34035 and PEFO 34231. These centra are longer than high (L/H ratio of 1.9:1 with lengths of 30 mm and 35 mm), platycoelus, and strongly keeled ventrally (Figures 4.13-4.16). In PEFO 2427 (Figure 4.16), there is a double keel (Long and Murry 1995), whereas in the larger specimens PEFO 34035 (35 mm length; Figure 4.13) and PEFO 34231 (36 mm length) there is only a single keel. The significance of this difference cannot be unambiguously determined given the material at hand, but either the two keels merge into a single larger keel through ontogeny of the individual or sacrals one and two differ in the number of keels that they possess. Other possibilities include individual or taxonomic differences. Raised facets for attachment of the sacral ribs cover much of the lateral side of the centrum (Figures 4.13-4.16). In one centrum (PEFO 2427) and the preserved sacral in PEFO 34035, the facet is subrounded in lateral view (Figure 4.15), whereas in PEFO 34231 and the other centrum in PEFO 2427 the facet is more anteroposteriorly elongate and particularly pronounced anteriorly. Long and Murry (1995) considered the centra with the subrounded facet from PEFO 2427 to represent the second sacral. All preserved sacral centra have open neurocentral sutures and no neural arches from this region are preserved. It is possible that the smaller centrum, figured by Long and Murry (1995, figure k) as the first sacral, represents an anterior caudal vertebra instead and that chevrons did not occur in the anteriormost caudals, similar to other archosauriforms. Evidence for this interpretation includes a shorter centrum (30 mm length) and the lack of a broad rib attachment; however, this interpretation cannot be confirmed with the evidence at hand. Hunt et al. (2005) also considered this centrum to belong to the caudal series. Long and Murry (1995) mention a centrum in PEFO 2427 that could represent an anterior caudal because of the lack of chevron facets but did not provide a more explicit identification of the element. Caudal. The caudal vertebrae of Vancleavea (Figures 4.17-4.18) are distinctive because of their box-like, rectangular shape in ventral view with almost no waisting and by the presence of two sharp ventral keels that originate at the posterior margins of the chevron facets and delineate the ventral surface of the centrum (Long and Murry 1995). All of the preserved caudal vertebrae are longer than high with oval articular faces with well- developed rims. In PEFO 2427 the presumably more anterior centra, which are anteroposteriorly shorter (average of 30 mm in length), have anteroposteriorly elongate, laterally projecting transverse processes. The more elongate (> 30 mm in length), and presumably more posteriorly situated caudal centra completely lack transverse processes. Pectoral Girdle
Pelvic Girdle
Ilium.
Both ilia are preserved in PEFO 34035. The left ilium is missing the posterior-most portion of the iliac blade and is diagenetically fused to the left femur; however, the right ilium is isolated and complete (Figure 7.1). The iliac blade is tall (40 mm) and separated from the acetabular by a well-defined neck. The blade apex is formed by the anterior portion of the blade. The dorsal margin slopes posteriorly toward a small posterior process.
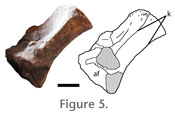
The ilium makes up almost the entire acetabulum. A distinct supra-acetabular rim is present, and the anterior margin of the ilium is almost twice the length of the posterior margin. The pubic and ischiadic peduncles meet at an obtuse angle; both are crescentic in distal view, curving slightly laterally (Figure 7.3; Hunt et al. 2005). The pubic peduncle is mediolaterally thickened compared to the ischiadic peduncle. Pubis. The proximal portions of both pubes are preserved in the holotype material (PEFO 2427). The pubis only contributes slightly to the acetabulum and possesses a distinct posterodorsal process (Figure 7.4). The articular surface for the ilium is mediolaterally thickened, rugose, and crescent shaped (Long and Murry 1995; Hunt et al. 2005). This surface continues posteriorly onto the posterior process and is confluent with a second rugose articular surface that is vertical and faces posteriorly to meet an anterior edge of the proximal portion of the ischium. The ventral surface of the posterior process of the pubis forms the dorsal rim of the obturator foramen. Because the ventral portion of the pubis is missing, it is not clear whether this foramen was fully enclosed by the pubis, or if it bordered the symphysis with the ischium (Long and Murry 1995). It is possible that the pubis and ischium met below this foramen, similar to basal archosauromorphs. The presence of a distinct thyroid fenestra, suggested for the Ghost Ranch specimen (Small and Downs 2002), cannot be determined from the PEFO material. Ischium. The ischium is known from proximal portions of both ischia preserved in PEFO 2427 and the proximal portion of the left ischium in PEFO 34035. Approximately a quarter of the acetabulum is formed by the ischium, a larger contribution than that of the pubis, but much less so than that of the ilium. The acetabular portion forms a distinct ventrally flaring semicircular rim (Figure 7.4; Long and Murry 1995). The articular surface for the ilium is rugose and curves posteromedially. There is an anterior projection with a subrounded anteriorly facing articular facet for the pubis. The ventral portion is not preserved. The groove mentioned by Long and Murry (1995) and questioned by Hunt et al. (2005) is interpreted here as a scallop-shaped fracture of the bone surface. Forelimb
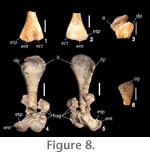
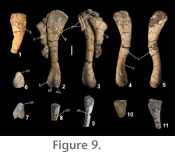
The distal end of the humerus is best preserved in PEFO 2427. The ento- and ectocondyles are distinct; the entocondyle is larger than the ectocondyle (Long and Murry 1995). There is a very slight concavity between the two condyles (contra Long and Murry 1995) and the distal surface is expanded medially forming a distinct entepicondyle (Figure 8.1). No entepicondylar or ectepicondylar grooves or foramina are present. The proximal and distal ends of the humerus are of equal width. The best preserved humerus (PEFO 34035) has a length of approximately 100 mm. Ulna. The proximal end of the left ulna is present in PEFO 34035 (Figure 8.6). The proximal articular surface is gently rounded convexly, and the head is inclined slightly posteriorly. The sigmoid notch is weakly expressed. An olecranon process is present but not strongly developed. Hind Limb
Tibia.
Proximal portions of tibiae are preserved in both PEFO 34035 and PEFO 2427 (Figure 9.6-9.11). In proximal view the element narrows anteriorly forming a distinct, but weak, cnemial crest (Figure 9.6-9.7).
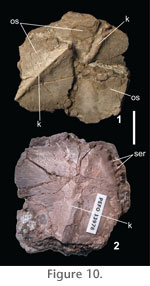
Osteoderms. Numerous osteoderms are preserved in the Petrified Forest Member material; however, most of these are poorly preserved. The osteoderms are sub-rounded with a median keel, prominent anterior prong, and serrated margins and are best preserved in PEFO 33978 (Figures 10.1-10.2). Unfortunately, the osteoderms are poorly preserved in the holotype; however, the fragmentary keeled osteoderms ("ankylosaur" type of Long and Murry 1995) are diagnostic of the genus (Hunt et al. 2002). Long and Murry (1995) also described two additional osteoderm morphotypes, tall, triangular osteoderms, and conical osteoderms, both described as " Stegosaurus -like." Neither of these morphotypes can be identified in PEFO 33978 or PEFO 34035. Hunt et al. (2005) considered the conical osteoderms to belong to the diminutive aetosaur Acaenasuchus geoffreyi , although there is no direct evidence for this interpretation. |
|