|
|
Introduction
The prosauropod Plateosauru engelhardti
Meyer, 1837 from the Late
Triassic of Central Europe is well known from many specimens from a number of
locations, several of them in nearly perfect articulation. Among early
dinosaurs, it is one of the best-known genera. The finds have caused heated
debate about the locomotory adaptations of the animal.
Huene (1907-08,
1926,
1928) argued for digitigrade bipedality and attributed a grasping function to
the manus. He was convinced that Plateosaurus was an obligate biped, much
as the similar Anchisaurus that had been described as exclusively bipedal
and digitigrade by Marsh (1893a,
1893b).
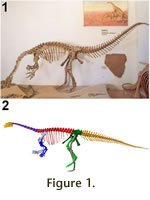 A good indicator for this is the highly
divergent length of fore- and hindlimbs. Huene had the mounts of GPIT1 and GPIT2
(Figure 1.1) set up in Tübingen in bipedal postures and adamantly stuck to this
interpretation of the material, despite criticism from many sources. A good indicator for this is the highly
divergent length of fore- and hindlimbs. Huene had the mounts of GPIT1 and GPIT2
(Figure 1.1) set up in Tübingen in bipedal postures and adamantly stuck to this
interpretation of the material, despite criticism from many sources.
Other researchers have suggested practically any possible stance: obligate
quadrupedality and plantigrady 'like lizards' was proposed by
Jaekel (1910), who
later changed his mind and concluded a clumsy, kangaroo-like hopping as the only
possible mode of locomotion (Jaekel 1911,
1913-14).
Fraas (1912,
1913) referred
to the (dorsoventrally compressed by sediment compaction) position of the
skeletal finds in the field, arguing for a sprawling obligatorily quadrupedal
gait. He had the skeleton SMNS 13200 mounted in this position in the Stuttgart
museum.
Later, researchers began to agree with Huene on the issue of digitigrady,
although plantigrady made a comeback in
Sullivan et al. (2003).
Weishampel and
Westphal (1986) depicted Plateosaurus running digitigrade and bipedally,
but they argued for facultative quadrupedality. Interestingly, the metacarpals
were shown widely spread, in marked contrast to the interpretation by
Huene
(1926, but contra the reconstruction drawing in that publication), a position
that does not seem to fit an active role of the manus in locomotion.
Paul (1997)
also argued for bipedality, but his outlined skeletal drawing seemed to imply
permanent quadrupedality, not bipedality. Facultative bipedality was also
proposed by Van Heerden (1997).
Among others, Galton (1971a,
1976,
1990,
2000) advocated facultative bipedality
in prosauropods. He based his opinion on the hindlimb to trunk ratio, which was
also invoked by Bonaparte (1971).
Wellnhofer (1994) also depicted
Plateosaurus in a quadrupedal stance, based on characteristics of the tail
of material from Ellingen now in the BSP, which he figured with a strong
downward curve making a bipedal stance impossible.
In the first functional morphology approach on Plateosaurus locomotion,
Christian et al. (1996) studied the vertebral column's resistance to bending in
various vertebrates in order to determine their locomotory modes. Since
Plateosaurus shows an intermediate pattern between obligate bipeds and
obligate quadrupeds, exhibiting a medium peak of resistance to bending over the
shoulders instead of either the small peak of bipeds or the large peak of
quadrupeds,
Christian et al. (1996) argued that the animal was probably facultatively bipedal at high speeds only.
Christian and Preuschoft (1996)
investigated the shape of the acetabulum and agreed with
Huene 1926 on a
near-vertical position of the femur in anterior view instead of a more sprawled
configuration.
The latest extensive publication on the osteology of Plateosaurus,
Moser
(2003), claimed that Plateosaurus would only have been capable of tiny
shuffling steps when walking bipedally. Moser also described the remounting of
the skeletal mount previously exhibited in the BSP for the Naturhistorische
Gesellschaft in Nuremberg. The animal was forced into what Moser called the
track of a quadrupedal prosauropod (Moser did not specify which track exactly;
probably he referred to the track depicted in
Moser (2003: fig. 28,
Tetrasauropus unguiferus, from
Ellenberger [1972]), despite the fact that
the track exhibits medially curving and rotated short toes and fingers, while
almost all articulated finds of Plateosaurus and the morphology of the
phalangal articular surfaces indicate no longitudinal rotation of the toes or
fingers. Also, the toes of Plateosaurus are long and slender.
Moser
(2003) suggested that this discrepancy may indicate an early sauropod instead of
a prosauropod as the trackmaker.
Galton (1971a,
1971b) already suggested medial
curving of the fingers in Plateosaurus, on the basis of GPIT1, and
proposed a walking configuration of the manus that kept the laterally diverging
first digit's claw off the ground.
Baird (1980) concluded that the track of
Navahopus falcipollex from the Navaho Sandstone of Arizona was made by a
quadrupedally walking plateosaurid dinosaur. The ichnofossil Otozoum,
originally described by Hitchcock (1847) and redescribed by
Rainforth (2003), is
another candidate for a prosauropod track: a bipedal track with two manus
imprints on which the fingers point outward at a right angle to the direction of
movement. The imprints fit the shape of the Plateosaurus manus and pes in
a semi-plantigrade position. The pes of Plateosaurus has been claimed
also to fit the pes print of, suggested as another possible quadrupedal
prosauropod track by
Lockley and Meyer (2000).
Porchetti and Nicosia (2007)
concluded that a Plateosaurus-like prosauropod is a possible creator of
Pseudotetrasauropus, again requiring a more or less plantigrade position.
The latest development in the controversy about the locomotory abilities of
Plateosaurus is an assessment of the range of motion of the forelimb of
Plateosaurus and the close relative Massospondylus by
Bonnan and
Senter (2007), which indicated that manus pronation was impossible. Therefore,
Bonnan and Senter (2007) concluded that Plateosaurus was an obligate
biped.
Recent research (Fechner 2006;
Remes 2006,
2008) suggests that the paradigm of
obligatorily bipedal ancestral dinosaurs may be wrong, and that the first
dinosaurs were at most facultatively bipedal, holding their hind limbs in
strongly flexed poses during quadrupedal locomotion. Such a posture allows
subequal functional limb and thus stride lengths despite significantly different
total limb lengths, and is similar to the posture of many small mammals. If this
model of early dinosaurian locomotion is correct, then the obligate quadrupedal
posture of sauropods is a primitive character, most small ornithischians with
highly different limb lengths may have been facultatively quadrupedal as well,
and bipedal posture would have evolved de novo and potentially separately in
several dinosaurian lineages, such as theropods, some 'prosauropods', and some
ornithopods.
The best approach to assess the locomotion capabilities of an extinct animal is
to create an exact 3D digital mount of it, based on a well-preserved complete
individual, and test all potentially possible postures for osteological and for
kinematic probability. For the latter, it is necessary to create an accurate 3D
model to determine the position of the center of mass (COM). Impossible postures
can thus be eliminated. Here, I test the hypothesis that Plateosaurus was
a facultative bipedal, using four-legged gaits for slow speed, while running
bipedally. I used two possible basic postures of Plateosaurus engelhardti
in various variations and assessed the position of the center of mass (COM) for
a spread of mass distributions for each posture. If only the first posture, a
quadrupedal stance, results in a stable and probable posture, it is safe to
assume that Plateosaurus was an obligate quadruped. The second posture,
bipedal, would indicate an obligate biped if it is the sole feasible posture. If
both postures are possible, at similar or different walking speeds, it is
probable that Plateosaurus may indeed have been capable of both
locomotory modes.
NOTE: Different researchers have articulated the bones of Plateosaurus
differently, both in museum mounts and in reconstruction drawings. Those
reconstructions that have a bearing on the investigations described here will be
discussed below with the details of the articulation of the digital skeleton of
Plateosaurus.
|