| |
DISCUSSION
Extant hindgut fermenters have more robust jaws than ruminants, when size is removed from comparison, and regardless of whether dietary habit (grazer versus browser) is considered. "Robustness" is defined by our criteria of a jaw possessing lower mandibular stress than other jaws subject to the same loading regime. A similar trend is seen in the pairings that include extinct equids, which can be assumed on phylogenetic grounds to be hindgut fermenters, with extant ruminants. A prediction from this result is that this methodology could be used to identify the probable digestive physiology of extinct ungulates with no extant relatives, such as the endemic South American litopterns and notoungulates.
Feeding Strategy and Body Size
As previously discussed, feeding strategy has a significant effect on jaw morphology. For example, grazers have larger masseter muscles than browsers (Clauss et al. 2009b), which will produce larger forces during mastication, and the jaws of grazers appear to be more robustly built in terms of general morphology. However, no clear pattern of differences between stresses in browsers and grazers within digestive physiology groups emerges from this study. Perhaps surprisingly, some of the lowest stress levels (and hence higher robustness) were recorded in the smaller browsing forms, in the tree hyrax (Dendrohyrax) among the hindgut fermenters, and the mouse deer (Tragulus) among the ruminants and also in the larger browsing tapirs (hindgut fermenters). It appears that depth of the jaw at the point in front of the mandibular angle generally has the most effect on robustness here, but further work would be required to elucidate the true nature of the biomechanics involved. It might be the case that smaller animals can "afford" to have jaws that are more robust than larger ones, but that with increasing body size the absolute weight of the jaw becomes an increasing consideration in craniodental design, given that it is manipulated with muscles placed only at the posterior end, with a relatively weak mechanical advantage. Of course, this remains to be tested.
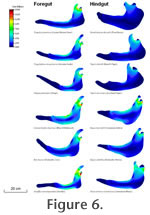 These observations on extant browsers can also be seen in some extinct ones. Hyracotherium is the earliest known (early Eocene) and most primitive fossil equid, usually considered to have a folivorous/frugivorous type of diet (like that of a modern mouse deer), due to its brachydont and bunolophodont cheek teeth (MacFadden 2005). However, despite the apparent gracile nature of the jaw morphology (see
Figure 6.1), the FEA analysis revealed the jaw of this species shows low levels of stress (0.93 MPa), and thus possess a high level of robusticity, much more so than in other, slightly younger brachydont equids (Mesohippus), or even the more closely related, contemporaneous Eurohippus. One morphological feature that may be related to this is the unusual (for an equid) deepening of the mandibular corpus beneath the premolars. This morphology could suggest feeding adaptations such as cracking seeds or nuts, a notion possibly supported by dental microwear (Solounias and Semprebon 2002 p. 30). Alternatively, at least one species of Hyracotherium (H. tapirinum) has been shown to be sexually dimorphic in canine size, with (presumed) males having larger canines (Gingerich 1981). If this mandibular robusticity was sexually dimorphic it might relate to male fighting behaviour. Further study would be needed to elucidate this idea. These observations on extant browsers can also be seen in some extinct ones. Hyracotherium is the earliest known (early Eocene) and most primitive fossil equid, usually considered to have a folivorous/frugivorous type of diet (like that of a modern mouse deer), due to its brachydont and bunolophodont cheek teeth (MacFadden 2005). However, despite the apparent gracile nature of the jaw morphology (see
Figure 6.1), the FEA analysis revealed the jaw of this species shows low levels of stress (0.93 MPa), and thus possess a high level of robusticity, much more so than in other, slightly younger brachydont equids (Mesohippus), or even the more closely related, contemporaneous Eurohippus. One morphological feature that may be related to this is the unusual (for an equid) deepening of the mandibular corpus beneath the premolars. This morphology could suggest feeding adaptations such as cracking seeds or nuts, a notion possibly supported by dental microwear (Solounias and Semprebon 2002 p. 30). Alternatively, at least one species of Hyracotherium (H. tapirinum) has been shown to be sexually dimorphic in canine size, with (presumed) males having larger canines (Gingerich 1981). If this mandibular robusticity was sexually dimorphic it might relate to male fighting behaviour. Further study would be needed to elucidate this idea.
Model Limitations
The finite element models presented here are two-dimensional and can only measure in-plane stress and strain generated by behaviour such as parasaggital bending. In vivo strain gauge, data and kinematic analysis of feeding in two ruminating artiodactyls (goat and alpaca) (Lieberman and Crompton, 2000;
Williams et al. 2007,
2009) demonstrate that the working side mandible also undergoes torsion about its longitudinal axis, and transverse and/or parasagittal bending in the balancing side mandible. Feeding loads are therefore experienced in three dimensions in these artiodactyls, and probably also in other ungulates, due to the laterally directed adductor muscle resultant seen in these taxa. Our 2D FE models restrict the extent to which the full effect of ungulate feeding loads on jaw morphology can be assessed. Further work using three dimensional FE-models is desirable, but there are practical and time-constraint issues to this process. Computed tomography (CT) scanning would be required to capture the internal and external 3D geometry of specimens, and for some taxa
that are poorly preserved or of a large size, scanning would be required to
capture the internal and external 3D geometry of specimens, which would be
difficult for taxa that are poorly preserved or of a large size. This paper offers a first step towards these future research directions.
|