| |
MATERIALS AND METHODS
Study specimens - 25 specimens of 23 species were included (of which 8 species were extinct) (Table 1) from the University Museum of Zoology Cambridge (UMZC), the American Museum of Natural History, New York (AMNH), the collections of the University of Bristol School of Biological Sciences (UBBS), and from the Digimorph collection of the University of Texas at Austin (http://digimorph.org) (see Appendix for details). Lateral aspect photographs comprised the majority of raw data, with the exception of a CT scan of a skull of Tapiris terrestris (lowland tapir), from which a lateral aspect image was created. This specimen was originally obtained from collections of the Texas Memorial Museum, University of Texas, United States of America (TMM).
In general, grazers tend to have jaws that appear to be more robustly built than those of browsers (Janis 1990a,
1990b,
1995;
Clauss et al. 2007). To reduce the likely influence of diet, on craniodental morphological features, and also possibly confounding effects of body size (allometric scaling, etc.), ruminant and hindgut-fermenting species were paired according to feeding strategy (i.e., browser, mixed feeder or grazer) and body mass (Table 1). Comparisons were made between each matched pair, and then compared overall.
There are substantially fewer extant hindgut fermenters than ruminants in all
dietary types and size groups. This imbalance is problematic as it means we do not have a full representative range of feeding strategies and body masses for the two groups. To partially redress this balance and explore the hypothesis in a broader range of taxa (i.e., removing the influence of phylogenetic affiliation), we included a species of hyrax (Dendrohyrax dorsalis), and we also examined some extinct equids to increase the range of size and dietary comparisons. (Other hyraxes [such as species of the mixed-feeding Heterohyrax or the grazing species of Procavia] were not included because there are no extant [or extinct] ruminants of this small of a body size [< 5 kg] that have this type of more fibrous diet [all are browsers or frugivores].) The extinct equids were assumed to be hindgut fermenters, like all other perissodactyls, and their diets were estimated from their dental morphology, from the degree of hypsodonty (see
Janis 1995) and also from microwear studies (see
Solounias and Semprebon 2002). Ontogenetic variation in all taxa was accounted for by using only specimens of adult animals (as determined by a fully erupted third molar).
Modelling Teeth
To test the effect of tooth row inclusion, 2D jaw FE-models of 16 species were created with and without the tooth row (simplified to a quadrilateral block consisting of all premolars and molars). Scaling was 1:1 with 10 mm model surfaces used for all species below ~100 kg and 20 mm for those above. Constraints were added at the dentary condyle above the mandibular notch and the back edge of the coronoid process and downward force of 100 N applied to a distal node of the tooth row and results recorded at five point intervals along a vertical transect originating from the mid-point of the tooth row. Although mean Von Mises stress across the transect differed, similar patterns of stress distribution were recovered.
It was clear from this study, that the unique dental morphology of individual taxa would heavily influence perceived robustness. It appeared also that the treatment of the tooth row as an immobile strip was wholly inaccurate, and each tooth unit would require individual modelling if they were to be included. Besides time constraints, this creates a problem generally in that it would also require the inclusion of periodontal ligament (a fibrous soft tissue attached to the cementum) around the base of each tooth. Unlike bone and dentine (where stress and strain increase in proportion under normal loading conditions) soft tissue in general is known to act with nonlinear elasticity with some studies suggesting transmission of load from teeth to the mandible is affected as a result (Kober et al 2008). Finally, it is important to consider that the direction of force transmitted through the tooth acts principally in the vertical plane; having little impact on resulting mandibular stress patterns. These additional intrinsic variables and the time involved in modelling realistic tooth units was therefore not justified for the remit of this study.
2D FE-Models
To create 2D FEmodels, basic line outlines of the jaw were generated from lateral aspect photographs of all 25 mandibles. Finite element analysis (FEA) is an engineering analysis tool that calculates stress and strain in a digital structure after the application of user-defined loads. It is used increasingly to determine functional mechanical behaviour of zoological and palaeontological specimens (see
Rayfield 2007;
Richmond et al. 2005 for reviews). The digital structure of interest is divided into a finite number of element blocks of regular geometry, linked at apices by nodal points. The stress-strain behaviour of each discrete region is computed, dependent upon user-defined loads and material properties, to provide a composite picture of the mechanical behaviour of the structure. Two-dimensional FE models have a standard thickness (20 mm), and so the models used in this study capture the outline geometry of the mandible but do not account for any differences in mandibular thickness. Jaw images were digitised using a polyhedral line tool of the java-based imaging software ImageJ (http://rsb.info.nih.gov/ij/).
Planar (x,y) co-ordinates of the image outline were then plotted in the Geostar component of CosmosM Finite Element Analysis (FEA) package (v. 2.8, SRAC, Ca. USA and Cenit Ltd, UK). Spline curves were generated to connect imported co-ordinates to recreate 2D FE model geometry with appropriately scaled surfaces (Pierce et al. 2008,
2009;
Rayfield 2005).
A user error study was conducted by digitising the outline of the jaw of a plains zebra (Equus burchelli) 20 times. Error involved while capturing geometry was negligible with 0.98% difference in area between attempts, with areas of complex curvature appearing to produce the greatest variation. It is worth noting that these and more subtle deviations from true geometry are largely negated by recreation of object margins with the spline curve tool of the FE processing software.
The model was meshed using triangular 3-noded finite elements, which were then attributed the material properties of bovine Haversian bone (Young's modulus = 10 GPa; Poisson ratio = 0.4:
Reilly and Burstein 1975).
The 2D FE models were linearly transformed to the scale of Equus burchelli),
a mid-sized species (~250 kg) from the study set (Table 1). The scaling allowed
analysis of pure geometric properties, reducing the influence that skull size may have had on the results. Average element size and model thickness was 20 mm, with number of elements ranging from 807 to 1561 depending largely on shape.
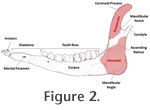 Loading and Constraint. Areas of attachment for both the temporalis and masseter muscles were standardized (Figure 2,
Figure 3) and the ratio for force allocation calculated. The masseter attachment was limited to an area below the mandibular notch between the condyle and coronoid process, and behind a vertical line drawn at the posterior border of the lower molar row (i.e., behind the third molar). The temporalis attachment consisted of the coronoid process area immediately above the mandibular notch. The models were constrained at both the distal anterior tooth of the tooth row in the Y direction to simulate a bite at this point, and an area extending from the posterior edge of the coronoid process and the condyle above the level of the mandibular notch: in all comprising six degrees of freedom to represent immobilisation of the mandible at the temporomandibular
joint. Loading and Constraint. Areas of attachment for both the temporalis and masseter muscles were standardized (Figure 2,
Figure 3) and the ratio for force allocation calculated. The masseter attachment was limited to an area below the mandibular notch between the condyle and coronoid process, and behind a vertical line drawn at the posterior border of the lower molar row (i.e., behind the third molar). The temporalis attachment consisted of the coronoid process area immediately above the mandibular notch. The models were constrained at both the distal anterior tooth of the tooth row in the Y direction to simulate a bite at this point, and an area extending from the posterior edge of the coronoid process and the condyle above the level of the mandibular notch: in all comprising six degrees of freedom to represent immobilisation of the mandible at the temporomandibular
joint.
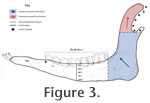 An arbitrary muscle force value of 100 N was used for comparative
purposes, but was distributed in proportion to surface area across the finite
element nodes of the masseter and temporalis in directions appropriate for the relative direction of force during mastication (Figure 3). An arbitrary muscle force value of 100 N was used for comparative
purposes, but was distributed in proportion to surface area across the finite
element nodes of the masseter and temporalis in directions appropriate for the relative direction of force during mastication (Figure 3).
Measurement. Stress measurements were taken from five evenly distributed nodes along a vertical transect of the model at the mid-line of the tooth row (Figure 3). These nodes were measured at 0%, 25%, 50%, 75% and 100% from the uppermost to lowermost point, respectively. Von Mises stress values were recorded for each of these points, and whole jaw colour plots were created to visualize the distribution of stress. Differences in stress patterns and magnitudes between ruminants and hindgut fermenters were analysed for all jaw models pooled together; pairs of extant ruminants and hindgut fermenters matched for dietary habit (grazer, browser, mixed feeder) and body size; pairs of extant ruminants and extinct hindgut fermenters also matched for dietary habit and body size.
|