|
|
|
TAXONOMIC NOTES AND COMMENTS ON SPECIES ECOLOGY
The geographical and environmental distribution of many species (marked with **) in our data set is given in Figure 7, Figure 8, Figure 9 so the details are not repeated here. However, for the less common species the statement concluding with * refers to the ATA distribution recorded in this study. Relative abundance terms: rare = occasional occurrence; minor = commonly present in abundance <10%; common >10%; dominant = the most abundant species in the assemblage. All other referenced ecological comments are based on live (stained) occurrences in the study area (NE Atlantic) unless otherwise specified. Where the notes on species ecology are from Murray (2006) they include references cited therein. Mainly Brackish Marginal Marine Taxa
Ammobaculites balkwilli
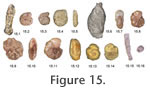
Haynes, 1973. Restricted to Europe; mainly low intertidal to subtidal; salinity 15-29 around the Skagerrak-Kattegat (Alve and Murray 1999). Ammoscalaria runiana (Heron-Allen and Earland) = Haplophragmium runianum Heron-Allen and Earland, 1916. Figure 15.2-5**. Intertidal-subtidal; common only in fine to medium sand with <20% mud and low TOC (0.2-0.7%); salinity 16-18 (Alve and Murray 1999). Ammotium cassis (Parker) = Lituola cassis Parker, 1870. Figure 15.6 (specimen metal coated)**. Infaunal detritivore lying horizontally in the top 5 cm of sediment; in brackish marginal marine environments, in the transitional water layer and possibly indicating the halocline; always subtidal; reproduces at <8°C (Olsson, 1976; Murray 2006). Ammotium salsum (Cushman and Brönnimann) = Ammobaculites salsus Cushman and Brönnimann, 1948**. Infaunal down to 10 cm, detritivore; marshes; withstands salinities of 60 (Murray 2006). Balticammina pseudomacrescens Brönnimann, Lutze and Whittaker, 1989**. Epifaunal; characteristic of low-salinity, brackish high marsh closest to land (Murray 2006). Haplophragmoides wilberti Anderson, 1953. Figure 15.7, 8**. Epifaunal and infaunal down to 30 cm; marshes (Murray 2006) Jadammina macrescens (Brady) = Trochammina inflata (Montagu) var. macrescens Brady, 1870. Figure 15.9-11**. Epifaunal, sometimes on decaying leaves, and infaunal down to 60 cm; herbivore or detritivore; widespread on high to mid marsh (Murray 2006). Miliammina fusca (Brady) = Quinqueloculina fusca Brady, 1870. Figure 15.12**. Epifaunal or infaunal down to 50 cm in marshes; intertidal to shallow subtidal in brackish waters (Murray 2006). The last survivor in marine environments that are cut off from the sea perhaps due to being infaunal (Lloyd and Evans 2002). Paratrochammina (Lepidoparatrochammina) haynesi (Atkinson) = Trochammina haynesi Atkinson, 1969. Figure 15.13, 14. Occasionally common in subtidal marginal marine*. Attached on gravel and shells, in the protection of crevices or ribs; current-swept areas such as channels (Murray and Alve 1993; Murray 2006). Reophax moniliformis Siddall, 1886. Figure 15.15, 16*. Detritivore on plant debris; husbands chloroplasts; intertidal to subtidal in brackish environments (Murray 2006). Tiphotrocha comprimata (Cushman and Brönnimann) = Trochammina comprimata Cushman and Brönnimann, 1948**. Epifaunal, free or clinging to algae, and infaunal down to 42 cm; herbivore or detritivore; marshes (Murray 2006). Trochammina inflata (Montagu) = Nautilus inflatus Montagu, 1808**. Epifaunal and infaunal down to 60 cm; herbivore or detritivores, mid marsh (Murray 2006). Shelf Sea, Shelf Deep and Fjord Taxa
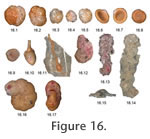
Prior to 1987 adult tests with three or four chambers in the final whorl were included in this species but in that year Brönnimann and Whittaker erected a new species (A. wrighti) for those with three chambers. Infaunal - epifaunal; a negative correlation with TOC but tolerates a wide range of temperature 1.8-16.6°C in Gulf of Guinea (Timm 1992). However, in the Arctic Ocean it favours temperatures of 1.8-3.0°C (Williamson et al. 1984). Inhabits the phytodetritus layer and sediment (Gooday 1988). It moves up and down in sediment seasonally: phytodetritus feeder in spring, shallow infaunal for the remainder of the year (Kuhnt et al. 2000). Adercotryma wrighti Brönnimann and Whittaker, 1987. Figure 16.3-5**. Ammodiscus catinus Höglund, 1947. Figure 16.6-8. Rare in shallow fjord, shelf, shelf deeps*. Ammodiscus gullmarensis Höglund, 1948. Figure 16.9. Occasionally common but mainly rare in shallow fjord, shelf, shelf deeps*. Brackish, salinity 20-32; temperature 4-14°C, on organic-rich muddy sediment, dominant in deeper water close to the permanently anoxic layer, in Norwegian fjords (Alve 1995). Ammolagena clavata (Jones and Parker) = Trochammina irregularis (d'Orbigny) var. clavata Jones and Parker, 1860. Figure 16.10, 11. Rare in Shelf deeps, deep sea*. Sea floor organic flux 0.8-60 g m2 yr-1 (Altenbach et al. 1999). Ammoscalaria pseudospiralis (Williamson) = Proteonina pseudospiralis Williamson, 1858. Figure 16.12. More mature specimens have a longer uniserial part. Rare in shallow fjord, shelf, and common in shelf deeps*. Infaunal in top 1 cm (Barmawidjaja et al. 1992). Sea floor organic flux 7->100 g m2 yr-1 (Altenbach et al. 1999). Ammoscalaria tenuimargo (Brady) = Haplophragmium tenuimargo Brady, 1882. Figure 16.13-15. Rare in shallow fjord and deep sea, occasionally common in shelf deeps*. Cribrostomoides crassimargo (Norman) = Haplophagmium crassimargo Norman, 1826. Figure 16.16, 17**.
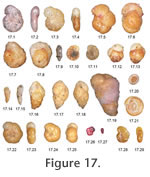
In #2070 there are smaller tests with a shiny surface and finer texture than the typical C. jeffreysii yet in other respects the morphology is the same. Tests with more mica are grey, and brown with no mica. We had considered using the name C. kosterensis but it was impossible to consistently separate the two. In Murray and Alve (2000a) we separated off Cribrostomoides sp. A but now that we have seen a larger data set we realise that it is not possible to consistently separate sp. A from C. jeffreysii. Infaunal to epifaunal, attached mobile (Murray 2006). Cribrostomoides nitidus (Goës) = Haplophragmium nitidum Goës, 1896. Figure 17.5, 6. Rare in shelf deeps, deep fjord, deep sea*. Cribrostomoides subglobosus (Sars) = Lituola subglobosa Sars, 1868. Figure 17.7, 8**. Epifaunal – infaunal, detritivore (Murray 2006). Sea floor organic flux 0.2-90 g m2 yr-1 (Altenbach et al. 1999). Linke (1989) suggested that the species entered a resting phase when food was in short supply and responded quickly to phytodetritus input. Cuneata arctica (Brady) = Reophax arctica Brady, 1881. Figure 17.9**. The range of morphology of Clavulina obscura Chaster seems to overlap that of this species. See Alve and Goldstein (2010) for further comments on synonymy. Deuterammina (Deuterammina) balkwilli Brönnimann and Whittaker, 1983. Minor in deep fjord*. Deuterammina (Deuterammina) rotaliformis (Heron-Allen and Earland) = Trochammina rotaliformis Heron-Allen and Earland, 1911. Rare in shelf, shelf deeps*. Deuterammina plymouthensis Brönnimann and Whittaker, 1990. Minor in shelf deep*. Deuterammina (Lepidodeuterammina) ochracea (Williamson) = Rotalina ochracea Williamson, 1858. Figure 17.10, 11**. Epifaunal, attached mobile (Murray 2006). Earlandammina bullata (Høglund) = Trochamminella bullata Höglund, 1947. Figure 17.12, 13. Brönnimann and Whittaker (1988) treat this as a synonym of E. inconspicua (Earland). However, that species is tiny and has a thin fragile wall – quite unlike our material or that of Höglund. Occasionally common in shelf deeps, minor in deep fjord and deep sea*. Eggerella europea (Christiansen) = Verneuilina europeum Christiansen, 1958, new name for Verneuilina advena Cushman of Höglund, 1947. Figure 17.14, 15**. #3234 has a few specimens that have a terminal biserial section. Eggerelloides medius (Höglund) = Verneuilina media Höglund, 1947. Figure 17.16**. Infaunal, positive correlation with % TOC and to a lesser extent with % <63 ?m sediment; a strong negative correlation with grain size >1000 ?m (Murray 2006). Eggerelloides scaber (Williamson) = Bulimina scabra Williamson, 1858. Figure 17.17, 18**. Infaunal, detritivore, subtidal, mainly shelf but tolerates salinity >24 for most of the year and temperatures 1-20°C (Murray et al. 2003; Murray 2006). Gaudryina pseudoturris (Cushman) = Textularia pseudoturris Cushman, 1922. Figure 17.19. Rare in shelf basin and deep sea*. Glomospira glomerata Höglund, 1947. Minor in fjord*. Glomospira gordialis (Jones and Parker) = Trochammina squamata var. gordialisJones and Parker, 1860. Figure 17.20, 21**. Shallow infaunal (Kuhnt et al. 2000). Sea floor organic flux 11-25 g m2 yr-1 (Altenbach et al. 1999). Haplophragmoides bradyi (Robertson) = Trochammina bradyi Robertson, 1891. Figure 17.22-25**. Shallow infaunal (Gooday 1990; Kuhnt et al. 2000). Haplophragmoides membranaceum Höglund, 1947. Figure 17.26, 27. Minor in shelf deeps, deep fjord, deep sea*. Haplophragmoides sp. 1. Figure 17.28, 29. Light brown yellowish, shiny, 4-5 chambers, flat. Rare in shelf, deep fjord*.
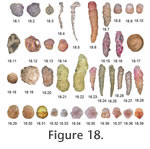
We have not found a species name for this. Rare in fjords, shelf and deep sea*. Lagenammina tubulata (Rhumbler) = Saccammina tubulata Rhumbler, 1931. Figure 18.3. Minor in shelf basin, deep fjord, deep sea*. Leptohalysis catella (Höglund) = Reophax catella Höglund, 1947. Figure 18.4**. Leptohalysis catenata (Höglund) = Reophax catenata Höglund, 1947. Minor in fjord and shelf*. Leptohalysis gracilis (Kiaer) = Reophax gracilis Kiaer, 1900. Minor in fjord, shelf basin, deep fjord*. Leptohalysis scottii (Chaster) = Reophax scottii Chaster, 1892. Figure 18.5. Fjord, shelf, shelf basin, deep fjord*. Under experimental conditions, lives in the top cm of sediment, responds quickly to disturbance and is very active, thus avoiding hostile environmental conditions, and is considered to be a good competitor (Ernst et al. 2000). Liebusella goesi Höglund, 1947. Figure 18.6**. Sea floor organic flux 7-90 g m2 yr-1 (Altenbach et al.1999). Morulaeplecta bulbosa Höglund, 1947. Figure 18.7. Occasionally common in shelf, rare in shelf basin*. Infaunal in top cm (Barmawidjaja et al. 1992). Paratrochammina (Lepidoparatrochammina) harti Brönnimann and Whittaker, 1990. Minor in shelf deeps*. Paratrochammina (Paratrochammina) wrighti Brönnimann and Whittaker, 1983. Minor in shelf*. Portatrochammina murrayi Brönnimann and Zaninetti, 1984. Figure 18.8-10**. The same as Trochammina globigeriniformis var. pygmaea of authors – including papers by Murray. Psammosphaera bowmani Heron-Allen and Earland, 1912. Figure 18.11. Minor in fjord and shelf*. Common just below the surface brackish layer in inner Drammensfjord, Norway (Alve 1990). Psammosphaera fusca Schultze, 1875. Figure 18.12**. Common in lower canyon at depths of 4810-4976 m off Portugal (Koho et al. 2007). Recurvoides trochamminiforme Höglund, 1947. Figure 18.13, 14**. Recurvoides turbinatus (Brady) = Haplophragmium turbinatum Brady, 1881. Minor in fjord*. Remaneica anglica Brönnimann and Whittaker, 1990. Minor in shelf*. Remaneica helgolandica Rhumbler, 1938. Minor in fjord and shelf*. Remaneica plicata (Terquem) = Patellina plicata Terquem, 1876. Minor in shelf*. Remaneicella gonzalezi (Seiglie) = Remaneica gonzalezi Seiglie, 1964. Minor in shelf and deep sea*. Reophax fusiformis (Williamson) = Proteonina fusiformis Williamson, 1858. Figure 18.15**. Normal marine salinity and temperature -1 to 13°C on sand and muddy sand down to >3000 m (Murray et al. 2003). Reophax micaceus Earland, 1934. Figure 18.16**. Reophax rostrata Höglund, 1947. Rare fjord, shelf*. Reophax scorpiurus Montfort, 1808. Minor in fjord, shelf and shelf basin*. Shallow infaunal (Kuhnt et al. 2000). Epifaunal; sea floor organic flux 0.8-60 g m2 yr-1 (Altenbach et al. 1999). Repmanina charoides (Jones and Parker) = Trochammina squamata Jones and Parker var. charoides Jones and Parker, 1860. Figure 18.17**. Shallow infaunal (Gooday 1990; Kuhnt et al. 2000). Saccammina socialis Brady, 1884. Figure 18.18**. Common in Laptev Sea (Wollenburg and Kuhnt 2000). Saccammina sphaerica Sars, 1872. Figure 18.19. Rare in shelf, deep fjord, deep sea*. Common in Laptev Sea (Wollenburg and Kuhnt 2000). Spiroplectammina biformis (Parker and Jones) = Textularia agglutinans d'Orbigny var. biformis Parker and Jones, 1865. Figure 18.20-22**. Estuarine, fjord, shelf in northern regions; tolerates salinity down to 20; low oxygen <2 ml/l (Williamson et al. 1994; Alve 1990; Alve and Nagy 1990). Technitella legumen Norman, 1878. Rare in shelf deep, deep sea*. Epifaunal , attached (Castignetti and Hart 2000). The genus is considered to thrive in organic-rich sediment and an opportunistic coloniser of barren sediments following sediment disturbance (Anschutz et al. 2002; Koho et al. 2007). Textularia contorta Höglund, 1947. Minor in shelf and deep fjord*. Textularia earlandi Parker = new name for Textularia tenuissima Earland, 1933. Figure 18.23-25**. Shallow infaunal, tolerates dysoxia (Bernhard et al. 1997). Seems to be an omnivorous opportunist (Alve 2010). Textularia kattegatensis (Höglund) = Textularia gracillima Höglund, 1947. Figure 18.26, 27. Minor in fjord, shelf, shelf deep*. Respiration rate 5.3 ± 1.9 nmol O2 d-1 ind-1 for individuals 563-611 ?m in length (Nomaki et al. 2007) Textularia skagerakensis Höglund, 1947. Figure 18.28. Minor in shelf, shelf deep, deep fjord, deep sea*. Tritaxis britannica Brönnimann and Whittaker, 1990. Figure 18.29, 30. Høglund (1947) called this Trochammina cf. rotaliformis. Rare in fjord, shelf and deep sea*. Tritaxis conica (Parker and Jones) = Valvulina triangularis d'Orbigny var. conica Parker and Jones, 1865. Figure 18.31. Minor in shelf deep*. Tritaxis fusca (Williamson) = Rotalina fusca Williamson, 1858. Minor in shelf and deep fjord*. Trochammina sp. skrumpa (shrunken chambers). Figure 18.32-35. Distinguished by its thin test wall, almost transparent, possibly made of mica. A low trochospiral. A characteristic feature is that that some chambers may collapse through shrinkage when dried. Subtriangular with 4 chambers in the final whorl, the final chamber forming at least one third of the test. Rare in shallow fjord, deep fjord, shelf basin and deep sea*. Some chambers may collapse through shrinkage when dried. Trochamminopsis quadriloba Höglund, 1948 = Trochammina pusilla Höglund, 1947. Figure 18.36-39**. Deep Sea TaxaBuzasina ringens (Brady) = Trochammina ringens Brady, 1879. Rare in deep sea*.
Shallow infaunal (Kuhnt et al. 2000). Tolerates dysoxia (Schönfeld 1997). Sea floor organic flux 1-9 g m2 yr-1 (Altenbach et al. 1999). Eratidus foliaceus (Brady) = Lituola (Haplophragmium) foliaceum Brady, 1881. Figure 19.2-4**. Infaunal; correlates with higher TOC but tolerates temperature of 1.8-7.9°C in Gulf of Guinea (Timm 1992). Sea floor organic flux 0.2-20 g m2 yr-1 (Altenbach et al. 1999). Shallow infaunal (Kuhnt et al. 2000). Glaphyramina americana (Cushman) = Ammobaculites americanus Cushman, 1910. Figure 19.5. Rare in deep sea*. Haplophragmoides sphaeriloculus Cushman, 1910. Figure 19.6, 7**. Shallow infaunal (Gooday 1986; Kuhnt et al. 2000). Hormosinella distans (Brady) = Lituola (Reophax) distans Brady, 1881. Figure 19.8. Minor in deep sea*. Shallow infaunal (Kuhnt et al. 2000). Found in lower canyon at depths of 4810-4976 m in sediments low in phytodetritus and with refractory organic matter and subject to near-annual sediment gravity flows (off Portugal, Koho et al. 2007). Hormosinella guttifer (Brady) = Lituola (Reophax) guttifer Brady, 1881. Figure 19.9-11**. Epifaunal; may respond to sea floor organic fluxes/ phytodetritus inputs (Wollenburg and Kuhnt 2000; Fontanier et al. 2003); highest total occurrences at 4°C in the Barents Sea (Hald and Steinsund 1992). Hyperammina laevigata Wright = Hyperammina elongata Brady var. laevigata Wright, 1891. Minor in fjord, shelf deeps, deep fjord, deep sea*. Reophax pilulifera Brady, 1884. Figure 19.12. Rare deep sea*. Karrerulina apicularis (Cushman) = Gaudryina apicularis Cushman, 1911. Figure 19.13. The juveniles of this species are very difficult to distinguish from Eggerelloides medius. In samples with only the triserial tests we call them E. medius. Occasionally common in deep sea*. Deep infaunal (Kuhnt et al. 2000). Sea floor organic flux 0.8-10 g m2 yr-1 (Altenbach et al.1999). Labrospira scitula (Brady) = Lituola (Haplophragmium) scitulum Brady, 1881. Figure 19.14-16. Rare in deep sea*. Sea floor organic flux 0.9-50 g m2 yr-1 (Altenbach et al. 1999). Labrospira wiesneri Parr, 1950. Rare in deep sea*. Infaunal (Gooday 1990). Lagenammina arenulata (Skinner) = Reophax difflugiformis arenulata Skinner, 1961. Figure 19.17, 18**. Nodellum membranaceum (Brady) = Reophax membranaceum Brady, 1879. Minor deep sea*. Placopsilina confusa Cushman, 1920. Rare in deep sea*. Portatrochammina challengeri Brönnimann and Whittaker, 1988. Rare in deep sea*. Reophax bilocularis Flint, 1899**. Shallow infaunal (Kuhnt et al. 2000). Sea floor organic flux 0.1-90 g m2 yr-1 (Altenbach et al. 1999). Reophax dentaliniformis Brady, 1881. Rare in deep fjord and deep sea*. Shallow infaunal (Kuhnt et al. 2000). Sea floor organic flux 0.8-90 g m2 yr-1 (Altenbach et al. 1999). Trochammina subturbinatus Cushman, 1920. Figure 19.19, 20**. Other authors may have called this T. inflata in deep sea samples. The umbilical opening seems to get deeper as the individuals grow larger and the later chambers become more globular. Some have a kummerform final chamber. This form appears to be distinct from T. pseudoinflata Scott and Vilks, 1991 from the Arctic Ocean. Trochamminella siphonifera Cushman, 1943 Rare in deep sea*. |
|