

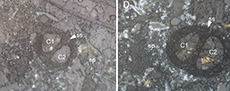
 Ovummuridae (calcareous microfossils) from the Much Wenlock Limestone Formation, Shropshire, UK
Ovummuridae (calcareous microfossils) from the Much Wenlock Limestone Formation, Shropshire, UK
Article number: 25.2.a27
https://doi.org/10.26879/1222
Copyright Palaeontological Association, August 2022
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 5 April 2022. Acceptance: 25 July 2022.
ABSTRACT
Ovummuridae are calcareous, egg-shaped microfossils with an unknown taxonomic affinity. Their limited observation is due to their occurrence only within exceptionally preserved carbonate rocks that have undergone little to no diagenesis or aggrading neomorphism. The Much Wenlock Limestone Formation (Homerian) is famous for its exceptionally preserved and diverse fossil biota, but Ovummuridae have not been previously observed and reported from the formation. This paper introduces the population of Ovummuridae from the off-reef tract limestones of The Much Wenlock Limestone Formation, Shropshire, UK. The authors analysed 124 polished and etched thin sections using reflected light microscopy to detect the presence of Ovummuridae. A total of 6591 ovummurids were identified, including observations of several previously undescribed morphotypes of Ovummuridae. Three new genera, Munneckella, Natantesprifmata and Hartonella, and subsequently four new species, Minourella wenlockiensis, Munneckella tribuscamera, Natantesprifmata rogersi and Hartonella oblonga, are introduced. The implications of this study suggest that reflected light microscopy is an effective and efficient method for observing calcareous microfossils. Ovummuridae may be more abundant than previously reported, their stratigraphic and palaeogeographic range is further extended, and it is highlighted that the off-reef tract may have been the microfossils’ preferred environment.
Rowshi Hussain. Curtins, Bridge Street, Birmingham, UK. rowshi.hussain@curtins.com
Steven L. Rogers. School of Geography, Geology and the Environment, Keele University, Keele, Staffordshire, UK. Correspondence author. s.l.rogers@keele.ac.uk
Joel A. Blackburn. School of Geosciences, St Mary’s Building, University of Aberdeen, Aberdeen, UK. joel.blackburn@abdn.ac.uk
Keywords: Ovummuridae; Wenlock Edge; calcareous microfossils; Silurian of Gotland; reflected light microscopy
Final citation: Hussain, Rowshi, Rogers, Steven L., and Blackburn, Joel A. 2022. Ovummuridae (calcareous microfossils) from the Much Wenlock Limestone Formation, Shropshire, UK. Palaeontologia Electronica, 25(2):a27. https://doi.org/10.26879/1222
palaeo-electronica.org/content/2022/3675-ovummuridae-wenlock-uk
Copyright: August 2022 Palaeontological Association.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
http://zoobank.org/AA8D31D4-1A87-49BD-ADEA-BD368676402E
INTRODUCTION
Ovummuridae have only been reported from a dozen or so localities globally (Munnecke et al., 2001), and only recently reported (Rogers et al., 2017) from Wenlock Edge (Shropshire, UK), despite the Wenlock Edge area having been extensively researched for centuries (Murchinson, 1833; Crosfield and Johnston, 1914; Shergold and Bassett, 1970; Scoffin, 1971; Hurst, 1975; Bassett, 1976; Ratcliffe, 1988; Bassett, 1989; Ratcliffe, 1991; Ratcliffe and Thomas, 1999; Ray and Thomas, 2007; Ray et al., 2010). The unknown biological affinity of Ovummuridae and its minimal reported occurrence within the literature makes them problematic to understand (Minoura and Chitoku, 1979; Munnecke et al., 2000).
The recent observation of Ovummuridae from the Wenlock Edge area (Rogers et al., 2017) included no detailed systematic study. This study provides: 1) systematic reporting of 6591 specimens of Ovumurridae from the Wenlock Edge Limestone Formation; 2) three new genera, Munneckella, Natantesprifmata and Hartonella and subsequently four new species, Minourella wenlockica, Munneckella tribuscamera, Natantesprifmata rogersi and Hartonella oblonga; 3) the increased palaeogeographical extent of the Ovummuridae; and 4) the extension of the known stratigraphic range of M. cameroni to Homerian (Wenlock), Silurian to the Frasnian-Famennian, Upper Devonian.
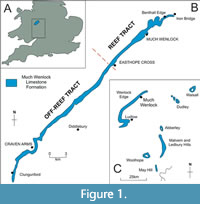 The Much Wenlock Limestone Formation (MWL Fm.) is a well-known “classical” formation in geological science. The focus of this study is the exposed outcrops of the MWL Fm. around the Much Wenlock area, Shropshire, UK (Figure 1). Wenlock Edge exhibits exceptionally preserved outcrops of the MWL Fm., making it an ideal area of study. Material from several localities, including Lea Quarry, Harton Hollow, Moorwood Quarry, Longville Stanway, Strefford Quarry, Burrington and the Ludlow Anticline, were studied.
The Much Wenlock Limestone Formation (MWL Fm.) is a well-known “classical” formation in geological science. The focus of this study is the exposed outcrops of the MWL Fm. around the Much Wenlock area, Shropshire, UK (Figure 1). Wenlock Edge exhibits exceptionally preserved outcrops of the MWL Fm., making it an ideal area of study. Material from several localities, including Lea Quarry, Harton Hollow, Moorwood Quarry, Longville Stanway, Strefford Quarry, Burrington and the Ludlow Anticline, were studied.
The deposition of the Silurian aged MWL Fm. occurred on a gently subsiding shelf margin forming a large carbonate platform, which is today exposed in two key areas: Wenlock Edge (Shropshire) and as series of small inliers between Dudley and Walsall (West Midlands) (Ratcliffe and Thomas, 1999; Ray et al., 2010). The formation is exceptionally well preserved in outcrops due to little or no evidence of diagenetic overprint, alteration, dolomitisation or neomorphism in the region (Ratcliffe and Thomas, 1999; Munnecke et al., 2000).
The MWL Fm. exposed at Wenlock Edge visibly extends as a lateral belt from southwest to northeast from Craven Arms to Much Wenlock, Shropshire (Murchinson, 1883). The reef tract present within the northeast of Wenlock Edge exhibits reefal limestones with various biological assemblages (Scoffin, 1971; Shergold and Bassett, 1970). The off-reef track is found to the southwest of Much Wenlock and is characterised by the absence of coral-stromatoporoid reefs (Scoffin, 1971; Shergold and Bassett, 1970). There are obvious variations along the outcrop belt that are observable laterally from both the sedimentology and palaeontology and which relate to the water depths at the time of deposition, as well as other environmental controls such as salinity and temperature (Holland et al., 1963; Shergold and Bassett, 1970; Scoffin, 1971; Ray et al., 2010). The reef tract section of the MWL Fm. represents a shallower depositional environment relative to the southwestern off-reef tract section towards the Ludlow Anticline (Fry et al., 2017).
The first discovery of Ovummuridae was reported from the Upper Pennsylvanian and Lower Permian limestones of Eastern Kansas, USA, via SEM analysis by Minoura and Chitoku (1979). Initially mistaking the microfossil for pollen spores, Minoura and Chitoku (1979) identified and described calcareous egg-shaped microfossils comprising two equal sized internal chambers separated by a septum and the brick-like calcareous ultrastructure of the wall. The characteristic wall-like or tablet nature of the ovummurid walls resembled that of a bryozoan but may have affinities to calcareous algae (Minoura and Chitoku, 1979). Minoura and Chitoku (1979) conclude that this was a newly undiscovered calcareous microfossil of unknown taxonomic affinity and established the genus: Ovummurus and the type species Ovummurus duoportius.
Munnecke et al. (2000) reported observations of Ovummuridae under SEM within the limestone-marl alternations from the Silurian of Gotland. They established the family Ovummuridae (Munnecke et al., 2000) and three new genera based on variation in morphology, Arouxina, Minourella and Samtlebenella, as well as three new species: Minourella gotlandica, Arouxina pluricellata and Samtlebenella circumcamerata (Munnecke et al., 2000). The work established the stratigraphical extent of certain Ovummuridae species by investigating several etched and polished thin sections from several locations. Munnecke et al. (2001) provided a more detailed and extended stratigraphical extent for Ovummuridae, ranging from the Llandovery (mid-Silurian) to the Guadalupian (Upper Permian), with no observations from Devonian sediments.
MacNeil and Jones (2006) followed up with observations of ovummurids under SEM within the late Devonian aged carbonate ramp of the Upper Escarpment and Alexander Formation from 10 cores. They showed that SEM was not required to detect the microfossils and provided evidence of ovummurids from observations using a petrological microscope using transmitted light (Leica DMLP microscope). In addition to the genera of ovummurids described by Minoura and Chitoku (1979) and Munnecke et al. (2000) they also reported and established a new species of Ovummuridae: Minourella cameroni from 30 recorded specimens.
METHODS AND MATERIAL
The materials used for this study are part of the thin section collection of Keele University (UK). Samples of the MWL Fm., including various lithofacies (the crinoidal grainstone, nodular limestone and tabular mudstone and limestone) were collected from the MWL Fm. from six localities around Wenlock Edge: B4371 road cutting near Longville, above Eaton Track, Harton Hollow, Lower Moorwood Quarry, Moorwood Quarry, Strefford Quarry, Craven Arms, Pitch Coppice, Ludlow Anticline, Burrington, Lower Dinchope to Strefford Lane and Upper Burrington Quarry (Figure 1 and Table 1).
The thin sections are 4.75 cm x 2.75 cm polished thin sections, mounted at a thickness of 60 μm, which were then etched during this study. Following a similar method to that given by Munnecke and Servais (1996), the sections were cleaned and then submerged into a solution of diluted 0.1M hydrochloric acid of a 1:10 ratio of distilled water for approximately 7 seconds. Once the sections were observed to be effervescing they were plunged into distilled water then placed on a tray to dry. Following etching, the thin sections were observed under a polarising petrographic microscope (model: Nikon Eclipse LV100N POL with digital sight DS-Fi-2 camera) using both transmitted and reflected light to produce several photomicrographs of every specimen of Ovummuridae observed. The reflected light method used here is outlined in Hussain et al. (2019), it is a quick and efficient method to detect these calcareous walled microfossils, which are otherwise difficult to see in transmitted light, or require time consuming and much more expensive SEM analysis. The drawback of using reflected light is that the images often include some glare (where reflective material is prominent on the surface compared to the calcareous microfossils due to the etching process, therefore it is often out of focus), and are not always as well-defined as SEM images.
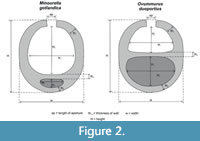 Each thin section was studied for Ovummuridae, and identified specimens were measured, imaged and recorded. Measurements were taken in the same manner as those made by Munnecke et al. (2000) (Figure 2). Each specimen was measured/described for their morphological and anatomical features, including: chamber shape, size and number; test shape; wall thickness; and septal structure. From this their taxonomical position (to at least the genus level) was established. The distribution of Ovummuridae across the MWL Fm., was established and compared to previous reports of the Ovummuridae. SEM analysis was later used to observe the details of the ultra-structure of the wall (and to measure wall thickness, in particular).
Each thin section was studied for Ovummuridae, and identified specimens were measured, imaged and recorded. Measurements were taken in the same manner as those made by Munnecke et al. (2000) (Figure 2). Each specimen was measured/described for their morphological and anatomical features, including: chamber shape, size and number; test shape; wall thickness; and septal structure. From this their taxonomical position (to at least the genus level) was established. The distribution of Ovummuridae across the MWL Fm., was established and compared to previous reports of the Ovummuridae. SEM analysis was later used to observe the details of the ultra-structure of the wall (and to measure wall thickness, in particular).
SYSTEMATIC PALAEONTOLOGY
The thin sections used in this study are stored at the School of Geography, Geology and the Environment at Keele University, United Kingdom. Each thin section is identified by their locality and sample number.
Phylum Incertae Sedis
Categorised as having an unknown taxonomic affinity, most fossil species are categorised as Incertae sedis if they have no known phylogenetic links.
Family OVUMMURIDAE
Munnecke, Servais and Vachard, 2000
Derivation of name. After Ovummurus, first genus described.
Genera included. Ovummurus Minoura and Chitoku, 1979; Minourella Munnecke, Servais, and Vachard, 2000; Arouxina Munnecke, Servais, and Vachard, 2000; Samtlebenella Munnecke, Servais, and Vachard, 2000; Natantesprifmata gen. nov; Hartonella gen. nov. and Munneckella gen. nov.
Original diagnosis (Summarised from Munnecke et al. 2000). Calcareous microfossils with an unknown biological affinity characterised by their small size and consisting of two or more chambers, an aperture at the apex of the chamber and a calcareous wall comprising of small slab or tablet like calcite crystals regularly arranged in concentric or pseudo-concentric layers.
Original description (Summarised from Munnecke et al. 2000). Spherical to ovoid or dolioform in shape with a maximum diameter of <100 µm. The walls are composed of 1 µm thick calcite tablets in roughly lamellar series and appear similar to those described in bryozoans and tentaculitoids (Larsson, 1979; Minoura and Chitoku, 1979; Munnecke and Samtleben, 1996). The wall thickness varies within different parts of the test. The partitioning of the internal cavity by a subequatorial or equatorial pseudo-septum is a common characteristic of the family. The apertures are variable, and, where present, determine the axial plane of the test.
Remarks. The Ovummuridae observed within the Much Wenlock Limestone Formation showed similar size ranges to those observed within the Silurian of Gotland with the majority of specimens under 100 µm. A few specimens observed did, however, exceed the original dimensions of 120 µm maximum diameter for Ovummurus as originally described by Minoura and Chitoku (1979), and some exceeded the 100 µm described by Munnecke et al. (2000). Here, we observe diameters ranging from 20 µm-298 µm.
The characteristics of the Ovummuridae are described as more or less spherical by Munnecke et al. (2000) and Minoura and Chitoku (1979), however, we note morphotypes of prismatic and elongated nature and some with triple chambers as first observed by Munnecke et al. (2000) followed by MacNeil and Jones (2006). Hence, we propose the addition of three new genera to the family. The composition of ovummurid walls in many specimens observed comprise pyrite crystals as well as the calcite tablets noted in the original diagnosis by Munnecke et al. (2000). Their walls are almost identical to the walls of bryozoans. Several specimens contained clastic grains within their chambers similarly described by Minoura and Chitoku (1979), Munnecke et al. (2000) and MacNeil and Jones (2006). MacNeil and Jones (2006) were unconvinced of reports of Samtlebenella circumcamerata sp. within the Upper Carboniferous samples of Munnecke et al. (2001) and believed they were mistaken for bryozoans. Here, we observe a small number of Samtlebenella circumcamerata with one or two chambers within their walls.
Stratigraphical range. Lower Silurian to Upper Permian (Munnecke et al., 2001).
Genus OVUMMURUS
Minoura and Chitoku, 1979
Type species. Ovummurus duoportio Minoura and Chitoku, 1979. Upper Pennsylvanian, Kansas, USA, by original designation.
Original diagnosis (Summarised from Minoura and Chitoku, 1979). A free calcareous microorganism characteristically ovoid to ellipsoidal shaped walls. The internal chambers are oval to elliptical and are diagonally divided into two equal cavities by a septum-like structure. The test is distinctive of two narrow slit-like apertures extending symmetrically to the centre and diagonally towards both the short and long axes of the test. These apertures equate to the length of the longer axis. Along the aperture the test is elevated to give shape to slender lips surrounding the apertures.
Remarks. The shape of the aperture in almost all specimens is difficult to determine, Munnecke et al. (2000) suggest the aperture is cylindrical rather than slit-like as described by Minoura and Chitoku (1979). Areal apertures are observed in both dorsal and ventral chambers.
Stratigraphical range. Lower Silurian to Upper Permian (Munnecke et al., 2001).
Species Ovummurus duoportius
Minoura and Chitoku, 1979
Figure 3A-D
Material. 194 specimens.
 Original diagnosis. Species diagnosis the same for genus (Minoura and Chitoku, 1979).
Original diagnosis. Species diagnosis the same for genus (Minoura and Chitoku, 1979).
Description (Modified from MacNeil and Jones, 2006). Ovoid to elliptical shaped test with varying sizes based on the plane of intersection depending on a sagittal or transverse cross section. The aperture to each chamber is only visible in longitudinal cross sections, and sometimes the wall around the aperture is thickened. Sometimes flange-like features on the outer walls of an aperture are visible.
Dimensions. MacNeil and Jones (2006) report measurements of: test, 160-230 µm long, 70-100 µm wide and 65-70 µm high, the wall and septum-like structure is approximately 3-12 µm thick. Our specimen values are smaller with test ranges between 14-142 µm in height and a test width of 14-86 µm, a wall thickness of 1-14 µm and septum thickness of 1-18 µm.
Remarks. It is very difficult to obtain three-dimensional measurements, as noted by MacNeil and Jones (2006). Only the test height and length have been noted as it is near impossible with two-dimensional cross-sectional cuts to determine the three-dimensional measurements. The population of Ovummurus observed was smaller than previously noted by MacNeil and Jones (2006); none of our specimens conform to the 160-230 µm test range given by them, with specimens reaching a maximum test length of 142 µm. The average test height of Ovummurus was recorded as 53 µm and width of 39 µm. However, we observed the largest overall test range between 14-142 µm in height and an overall test width of 14-86 µm. We note the largest wall thickness with a range from 1-14 µm and a septum thickness of 1-18 µm. The largest aperture observed was 3.8 µm wide.
Stratigraphic range. Lower Silurian to Upper Permian (Munnecke et al., 2001).
Genus MINOURELLA
Munnecke, Servais, and Vachard, 2000
Type species. Minourella gotlandica.
Diagnosis (Modified from Munnecke et al., 2000 and MacNeil and Jones, 2006). The morphology of Minourella is similar to the Tubertinidae, a Foraminifera. It is characterised by a bilocular, globular or conical test with a dorsal section that is gently vaulted with a small apical aperture in the central part. The ventral chamber corresponds to the second chamber and is with or without an aperture. The thickness of the wall is usually constant but can also be non-uniform throughout even at the aperture with the pseudo-septum being relatively thinner than the wall (Munnecke et al., 2000). The external and internal surfaces are smooth.
Remarks. Diagnosis amended to include a non-uniform wall thickness as observed within various specimens of Minourella gotlandica. The apertures are present within both the dorsal and ventral chambers in some of the specimens as similarly noted by MacNeil and Jones (2006). We position M. wenlockensis in the Genus Minourella because: (1) it resembles the same morphology as M. gotlandica, however, contains a conical ventral chamber that has a sharp apex rather than a smooth apex, (2) has the characteristic wall structure of Ovummuridae, (3) and meets the same ranges of measurements of Minourella.
Stratigraphical range. Silurian to Upper Devonian.
Minourella gotlandica
Munnecke, Servais and Vachard, 2000
Figure 4A-D
Material. 4665 specimens.
 Diagnosis (Modified from Munnecke et al., 2000 and MacNeil and Jones, 2006). A species of Minourella, consists of one large and one small chamber, separated by a flat septum-like structure. Rare specimens exhibit an apical aperture in the smaller ventral chamber and occasionally within both ventral and dorsal chambers.
Diagnosis (Modified from Munnecke et al., 2000 and MacNeil and Jones, 2006). A species of Minourella, consists of one large and one small chamber, separated by a flat septum-like structure. Rare specimens exhibit an apical aperture in the smaller ventral chamber and occasionally within both ventral and dorsal chambers.
Description (Modified from Munnecke et al., 2000 and MacNeil and Jones, 2006). A globular micro-organism characterised by two asymmetrical chambers separated by a septum-like structure. The larger chamber is globular and oval shaped with the long axis parallel to the septum. The small chamber is globular and oval/elongated-oval shaped, similar to the dorsal larger chamber. The aperture within the dorsal chamber is not always present. The aperture in the ventral chamber is rare but can occur. Three specimens with an aperture at the apex of both the dorsal and ventral chamber were observed from the Wenlock Edge material.
Dimensions. Overall test height/length of 21-233 µm, overall test width of 14-284 µm, large chamber diameter height of 2-184µm and width of 20-245 µm, the wall around the large dorsal chamber is 0.47-35.5 µm thick. The smaller ventral chamber height of 1-113µm and width of 1-103 µm the wall around the small ventral is 0.5-16 µm thick. A septum thickness of 0.4-36 µm and an aperture of 1-14 µm in diameter.
Remarks. Diagnosis amended to include the possibility of an aperture present both within the dorsal and ventral chambers. It is noted that very rare specimens exhibit apertures within both their dorsal and ventral chambers unlike in the Gotland material in which a single aperture was reported to be visible in the dorsal chamber and conversely only from the ventral chamber within the Hay River material (MacNeil and Jones, 2006). The specimens observed are comparable in morphotype and size to those described by Munnecke et al. (2000), however, the MWL Fm. specimens are in general: (1) larger, (2) have thicker walls with quartz overgrowths, (3) some specimens contain pyrite within their walls and/or within their chambers as a result of diagenesis, (4) larger apical apertures and exhibit apertures within the ventral chamber. The largest Minourella gotlandica specimen is noted as having a test height of 233 µm and a test length of 284 µm. A maximum wall thickness of 36 µm and a septum thickness of 19 µm. The largest aperture width noted was 14 µm.
Stratigraphical range. Lower Silurian (Homerian) to Devonian (Famennian)
Minourella cameroni
MacNeil and Jones, 2006
Figure 4H
Material. 7 (27, if triangular cross-sectional cuts are considered)
Original diagnosis (MacNeil and Jones, 2006). Conical test, one large dorsal chamber and one small ventral chamber with apertures at anterior end of test and a dorsal secondary aperture.
Description (Modified from MacNeil and Jones, 2006). A conical test dominated by a large, anteriorly expanding dorsal chamber. The septum partly divides the dorsal chamber into a large, posterior sub-chamber and small, anterior sub-chamber. The large, posterior dorsal sub-chamber can contain a secondary aperture, possibly slit-like, on the dorsal side of the test. The small sub-chamber can have a large simple aperture at the anterior dorsal end of the test, with or without lateral flanges. The small ventral chamber narrows around and over the area of the dorsal chamber that contains the septum before opening into an aperture at the anterior end.
Dimensions. MacNeil and Jones (2006) introduced measurements of overall test length of 195-275 μm long, 70-115 μm high, dorsal chamber height of 55-105 μm high, ventral chamber 0.5-13 μm high, and walls 3-6 μm thick. Our specimen measurements were: overall test height/length of 42-298 µm, overall test width of 7-132 µm, large dorsal chamber diameter height of 20-260 µm and width of 5-71 µm and the wall around large dorsal chamber is 2-14 µm thick. The smaller ventral chamber has a height of 2-115 µm and width of 1-32 µm and the wall around the small dorsal ventral is 2-5 µm thick. A septum thickness of 2-6 µm.
Remarks. The specimens from the MWL Fm. show no visible apertures in either of the chambers as described by MacNeil and Jones (2006). Although in some cross-sections Minourella cameroni looks identical to Minourella gotlandica, it is difficult to determine the true specimen count of the species. Many specimens of ovummurids were triangular in morphotype, and we believe these may be transverse cross-sectional cuts through Minourella cameroni. The MWL Fm. specimens in comparison to the Hay River Material are (1) lower in population, (2) thinner in septum thickness and generally smaller, (3) longer in test length with maximum test lengths of 298 μm were observed.
Stratigraphical range. Lower Silurian to Upper Devonian
Minourella wenlockensis sp. nov.
Figure 4E-G
zoobank.org/AC9E3A5C-4CE9-4A24-8A14-DF56DD236761
Derivation. After Wenlock Edge, Shropshire UK - the type locality of Wenlock aged strata.
Holotype. SQ1/51. Strefford Quarry, Nodular Packestones of the Off-reef Tract, Much Wenlock Limestone Formation, Homerian.
Paratype. 3MQ/54. Moorwood Quarry, Nodular Packestones of the Off-reef Tract, Much Wenlock Limestone Formation, Homerian.
Material. 858 specimens.
Diagnosis. A species of Minourella with an overall conical test, comprising one large globular dorsal chamber and one small conical ventral chamber with apertures at the anterior end of test and a dorsal secondary aperture. The wall of the microorganism comprises linearly arranged calcite tablets.
Description. Measurements are similar to Minourella gotlandica. The dorsal chamber is globular and can be oval shaped. The ventral chamber is conical with a sharp apex. The aperture is cylindrical and in some cases conical and can be present in either or both dorsal and ventral chambers, simultaneously.
Dimensions. Overall test height of 29-212 μm and an overall test width of 17-136 μm. Large dorsal chamber height of 7-151 μm and a diameter of 10-115 μm with a surrounding wall thickness of 0.8-23 μm. The smaller ventral chamber has a height of 7.4-50.9 μm and a width of 9.6-115.4 μm, the thickness of the wall surrounding the chamber is 0.8-16 μm thick. An aperture ranging from 0.9-10 μm.
Remarks. This morphology is mentioned and described by both Munnecke et al. (2000) and MacNeil and Jones (2006) as a variant of Minourella gotlandica, however, the apex of the ventral chamber is sharp rather than smooth, from this distinct morphological difference is appropriate to assume it is another species belonging to the genus Minourella. Separated from M. gotlandica by its conical ventral chamber as opposed to a globular test. The ventral chamber closes as an abrupt sharp apex rather than smooth one like M. gotlandica. Apertures can be present within either dorsal/ventral chamber or within both simultaneously (see Figure 4E).
Stratigraphical range. Lower Silurian to Upper Devonian.
Genus AROUXINA
Munnecke, Servais, and Vachard, 2000
Derivation of name. After Alain Roux (1944-1993), French specialist on Palaeozoic calcareous algae and microproblematica; genus feminine.
Type Species. Arouxina pluricellata, the only species known.
Diagnosis (Modified from Munnecke et al. (2000). The test is free and multi-chambered composed of a cluster of 1-20 fine hemispherical chambers irregularly arranged, not necessarily in one plane. Chambers are finely joined or can be disjointed. The two initial chambers differ in shape from the others. They resemble those of Ovummurus or Minourella but differ by having a functional aperture on the second chamber. The group of initial chambers is dolioliform to ovoid in shape; the other chamberlets are more or less hemispherical or bulbous in rare occasions. The wall is composed of more or less linearly arranged, small calcite tablets.
Remarks. The MWL Fm. specimens show only 1-6 hemispherical chambers. The joining of the hemispherical chambers is sometimes disjointed, and the hemispheres are sometimes bulbous in shape. The material from Wenlock Edge shows Arouxina resembles Minourella gotlandica rather than Ovummurus duoportius but can sometimes be unclear due to cross-sectional cuts. The variation based on the initial dorsal and ventral chamber morphologies could form a potentially new species with further observations.
Arouxina pluricellata
Munnecke, Servais, and Vachard, 2000
Figure 5A-F
Derivation of name. Latin, pluri -, several; cella, small chamber.
 Material. 10 specimens
Material. 10 specimens
Diagnosis. Test composed of two initial chambers separated by a pseudo-septum and a relatively small to medium number of hemispherical additional chambers. Each chamber presents an apical cylindrical non-protuberant aperture. Wall is calcitic and lamellar typical of the Ovummuridae.
Description. An Ovummuridae characterised by two symmetrical or asymmetrical chambers separated by a septum like-structure. Initial chambers can be equal in size like Ovummurus or bilocular like Minourella. The larger chamber is globular and oval shaped with the long axis parallel to the septum. The small chamber is globular and oval/elongated-oval shaped similar to the dorsal chamber. The aperture within the dorsal chamber is not always present. The initial chambers are surrounded by free and disjointed hemispherical smaller chambers that occasionally build upon each other.
Dimensions. Overall test height of 48-37 μm and an overall test width of 47-137 μm long. Large dorsal chamber height of 14-75 μm and a diameter of 18-67 μm with a surrounding wall thickness of 4-29 μm. The smaller ventral chamber has a height of 3-24 μm and a width of 10-48 μm and the thickness of the wall surrounding the chamber is 0.9-13 μm thick. The septum thickness is 2-31 μm with an aperture ranging from 1-7 μm. The hemispherical chambers are approximately 13-25 μm in diameter and 6-13 μm height, the thickness of the surrounding wall measures between 4-10 μm.
Remarks. Various specimens observed here have fewer than 14 chambers as described by Munnecke et al. (2000). Here, we observe between 2-6 chambers. Munnecke et al. (2000) describe Arouxina pluricellata to resemble Ovummurus, however, we note morphologies that resemble Minourella gotlandica. The chambers are sometimes bulbous rather than hemi-spherical, and specimens that exhibit these bulbous growths tend to have several walls. Munnecke et al. (2000) suggest measurements are similar to those of Ovummurus and Minourella, noting overall chamber lengths of 80-100 μm and total widths of 50-80 μm. It is noted that the Wenlock Edge specimens have longer lengths and width measurements exceeding the 100 μm in length and 80 μm in width that was originally observed. Comparison of the hemispherical chambers is not possible as no measurements were provided by Munnecke et al. (2000).
Stratigraphical range. Lower Silurian (Homerian)
Genus SAMTLEBENELLA
Munnecke, Servais, and Vachard, 2000
Derivation of name. After Christian Samtleben, University of Kiel, specialist in calcareous nannofossils (coccolithophores) who initiated petrographic studies of the micritic limestones of the Silurian of Gotland; genus feminine.
Type species. Samtlebenella circumcamerata, the only species known.
Original diagnosis. Ovummuridae with an aperture and a pseudo-septum, characterised by its ellipsoidal chamberlets regularly distributed in the wall.
Remarks. Difficult to determine in cross-section and are very similar in appearance to Calcispheres and can only be distinguished by their walls.
Stratigraphical range. Lower Silurian (Homerian).
Samtlebenella circumcamerata
Munnecke, Servais, and Vachard, 2000
Figure 5G-H
Derivation of name. Latin, circum, around; camera, vault, chamber.
Material. 5 specimens
Original diagnosis. A species of Samtlebenella with 10-15 chamberlets regularly distributed in the wall.
Dimensions. Munnecke et al. (2000) noted total lengths of S. circumcamerata to measure between 80-125 µm, and the total width between 50-120 µm and observed between 10-15 chamberlets. Our specimens measure: overall lengths of 84-185 µm and diameters of 96-173 µm with only 1-4 chamberlets observed. The thickness of the wall is noted to vary between 4-27 µm.
Remarks. Specimens are undetermined if they are Samtlebenella circumcamerata as the chambers within the walls are not clear due to their small sizes and possibly could be juvenile specimens. Observations of 1-4 chamberlets only but could be due to cross-sectional cuts. We note specimens of a larger size than those observed at Gotland.
Stratigraphical range. Lower Silurian (Homerian)
Genus NATANTESPRIFMATA gen. nov.
zoobank.org/85D83F35-B299-4020-8305-FA7D4E87B29B
Derivation of name. Latin, natantes, floating; prifmata, prisms; genus; neutral
Type Species. Natantesprifmata rogersi sp. nov., the only species known.
Diagnosis. Ovummuridae with a prismatic test divided into two chambers by a pseudo-septum. The larger dorsal chamber is prismatic in shape. The smaller ventral chamber is conical or pyramidal. Apertures may be present within the central axial plane of the dorsal chamber. The wall is characteristic of Ovummuridae, exhibiting the lamellar brick-like arrangement of calcite tablets.
Remarks. These Ovummuridae specimens do not conform to the specifications of the genera: Minourella, Ovummurus, Arouxina or Samtlebenella because: (1) their chambers are not globular like Minourella or Ovummurus, (2) their chambers are not symmetrical like Ovummurus, (3) they do not exhibit hemispherical chambers around the initial chambers like Arouxina and (4) they do not contain chambers within their walls. On this basis, a new genus has been established to account for the prismatic nature of these variants of Ovummuridae. The ventral chamber is difficult to determine the test shape based on 2D cross sections, hence it is undetermined whether the ventral chamber is of conical or pyramidal nature. It is most likely pyramidal based on the terminations of the wall surrounding the secondary chamber and the pseudo-septum.
There are no known calcareous fossils of a prismatic nature, and neither do Palaeozoic foraminifera show such test geometries. The genus can be compared to Radiolaria and Dinoflagellates for their similarities in polygonal, polyhedron or prismatic test morphologies, however, these microfossil groups are siliceous rather than calcareous. Dinoflagellates such as: Protoperidinium communis and Protoperidinium oceanicum Biffi and Grignani 1983, are polygonal and exhibit the same anterior sharp apex as seen in Natantesprifmata. The pyramidal morphology is seen due to the apical horn in Protoperidinium communis and Protoperidinium oceanicum Biffe and Grignani 1983, rather than a pyramidal-shaped dorsal chamber as seen in these Ovummuridae. However, Protoperidinium are characteristic of having apical and antapical horns that are not seen in Ovummuridae.
The Family Pterocorythidae, described by Haeckel (1881) such as Stichopilium bicorne Haeckel (1887), show similarities to Natantesprifmata as their thorax and external shells are pyramidal when observed proximally due to their polyhedron nature. Similarly, Podobursa mersinensis sp.and Pterocanium prismatium (Lazarus, 1986) can appear prismatic in 2D cross-sections. However, these Radiolaria do not comprise chambers but consist of porate cephalis and commonly more than one postcephalic chamber is present (Haeckel, 1862). Radiolarians do not have the same wall structure that is characteristic of Ovummuridae as their shells are latticed (Haeckel, 1882; Minoura and Chitoku, 1979). They express long spines that emerge from their cephalis, which are not observed within Ovummuridae (Haeckel, 1862). The nassellarian skeletons are usually preserved as microcrystalline calcite.
Stratigraphical Range. Lower Silurian (Homerian).
Natantesprifmata rogersi sp. nov.
Figure 6A-D
zoobank.org/46757E61-FE70-4101-BF76-D970E3BCCA63
 Derivation of name. After Steven Rogers, University of Keele, geologist who initiated the student-led sedimentological and micropalaeontological study of the off-reef tract limestones from the Much Wenlock Limestone Formation, Wenlock Edge, Shropshire, UK; genus masculine.
Derivation of name. After Steven Rogers, University of Keele, geologist who initiated the student-led sedimentological and micropalaeontological study of the off-reef tract limestones from the Much Wenlock Limestone Formation, Wenlock Edge, Shropshire, UK; genus masculine.
Holotype. LST7/0057, Longville Stanway Track, Nodular Packestones of the Off-reef Tract, Much Wenlock Limestone Formation, Homerian.
Paratype. 1MQ/0023, Moorwood Quarry, Nodular Packestones of the Off-reef Tract, Much Wenlock Limestone Formation, Homerian.
Material. 214 specimens
Diagnosis. Same as for genus.
Description. Test composed of two asymmetrical chambers separated by a pseudo-septum. The dorsal chamber is larger and pyramidal and sometimes a cylindrical areal aperture may be present within the central axial plane posterior of the dorsal chamber. The ventral chamber is smaller and pyramidal/conical shaped and extends to terminate at a sharp apex. The walls thicken at the edges of the dorsal chamber. The walls are calcitic and lamellar resembling a brick-like arrangement. The chambers occasionally host clastic grains and pyrite.
Dimensions. Overall test height of 44-126 μm and an overall test width of 34-120 μm long. Large dorsal chamber height of 5-71 μm and a diameter of 7-61 μm with a surrounding wall thickness of 0.9-15 μm. The smaller ventral chamber has a height of 3-48 μm and a width of 5-50 μm and the thickness of the wall surrounding the chamber is 0.5-11 μm. A septum thickness of 0.5-10 and an aperture width ranging from 0.9-10 μm.
Remarks. Specimens of Natantesprifmata rogersi appear quadrilateral or resemble kites in cross-section. Their walls can include pyrite as can their chambers. Sometimes clastic material is present within their dorsal or ventral chambers.
Stratigraphical range. Lower Silurian (Homerian).
Genus HARTONELLA gen. nov.
zoobank.org/68D3F0D5-A429-4F3E-9B75-7B99E61B9A01
Derivation of name. After Harton Hollow, Shropshire, UK. The name of the locality the genus was first found from, genus feminine.
Type species. Hartonella oblonga sp. nov., the only species known.
Diagnosis. Elongated oval shaped test comprising of two asymmetrical chambers. The larger dorsal chamber is elongate, oval shaped separated by a pseudo-septum from the ventral oval to hemispherical chamber. The ventral chambers apex can be either smooth or sharp. An aperture is present within the central axis of the posterior of the dorsal chamber. The wall is comprised of calcite tablets that are in a lamellar arrangement.
Remarks. These Ovummuridae specimens do not conform to the specifications of the genera: Minourella, Ovummurus, Arouxina or Samtlebenella because: (1) their chambers are not globular like Minourella or Ovummurus and are elongate and tubular, (2) their chambers are not symmetrical like Ovummurus, (3) they do not exhibit hemispherical chambers around the initial chambers like Arouxina and (4) they do not contain chambers within their walls like Samtlebenella. Therefore, based upon their morphological differences and measurements, a new genus has been established to account for the elongate nature of these variants of Ovummuridae.
The genus is comparable to other microfossils, such as foraminifera; lagenoid forms such as Nodosaria sp. (Loeblich and Tappan, 1964) have straight chambers similar to these elongate Ovummuridae variants. However, they also have characteristic planispirally coiled or uncoiled chambers which are not observed in Ovummuridae. Nodosariacea comprise uni-multilamellar chambers, and their tests are finely perforated, neither which are distinctive of the Ovummuridae (Loeblich and Tappan, 1964 and Minoura and Chitoku, 1979). Similarly, foraminifera such as: Earlandia ex gr. minor (Rauzer-Chernousova, 1948) and Miliolata Nubeculariina Calcivertelloidea: Calcivertella sp. Cushman and Waters, 1928 in cross section can have elongated tubular appearances similarly seen to these ovummurid variants. Although, they share characteristics in their undivided tubular test chambers, Calcivertella sp. walls are porcelaneous, and they have a streptospirally coiled test (Vachard et al., 2018).
Hartonella is anatomically comparable to chitinozoans, in particular, Conochitinia proboscifera Eiesenack, 1937. They are both tubular and elongated nature. However, unlike the chitinozoans, Ovummuridae are not chitinous, and their tests are formed of calcite tablets.
Stratigraphical range. Lower Silurian (Homerian).
Hartonella oblonga sp. nov.
Figure 6E-H
zoobank.org/B2257655-19E8-447F-A3FF-CBB4647C386C
Derivation of name. Latin, oblonga, elongated; genus neutral.
Holotype. 1HH/055, Harton Hollow, Nodular Packestones of the Off-reef Tract, Much Wenlock Limestone Formation, Homerian.
Paratype. 1HH/153, Harton Hollow, Nodular Packestones of the Off-reef Tract, Much Wenlock Limestone Formation, Homerian.
Material. 228 specimens.
Diagnosis. same as for genus.
Description. Test composed of two asymmetrical chambers that are elongate oval shaped. The dorsal chamber is elongated to tubular in shape and contains a cylindrical areal aperture at the posterior. The ventral chamber is hemispherical to conical in shape and can have either a sharp or smooth apex. The wall comprises lamellar arranged calcite tablets resembling the structure of a brick wall.
Dimensions. Overall test height of 24.1-296.8 μm and an overall test width of 22.1-258.1 μm long. Large dorsal chamber height of 17.6-111.3 μm and a diameter of 14.7-103.9 μm with a surrounding wall thickness of 0.5-19.2 μm. The smaller ventral chamber has a height of 2.9-32.3 μm and a width of 2.8-38.5 μm and the thickness of the wall surrounding the chamber is 0.5-9.7 μm thick. A septum thickness of 0.5-19.2 and an aperture width ranging from 0.9-3.2 μm.
Remarks. Specimens are different from Minourella gotlandica as they are more elongated, and their geometries are not due to longitudinal cross-sectional cuts. Variation is observed in their measurements that do not fit within the range of M. gotlandica. The apex of the ventral chamber can be either smooth or sharp. There is the possibility of establishing further species from specimens observed here, especially based on the shape of the ventral chamber.
Stratigraphical range. Lower Silurian (Homerian).
Genus MUNNECKELLA gen. nov.
zoobank.org/61267617-5BEC-4844-BED1-4B591F2CC20F
Derivation of name. After Axel Munnecke, Friedrich-Alexander-University of Erlangen-Nürnberg, instrumental in the erection of the Ovummuridae, observed the first three-chambered variants of Ovummuridae; genus feminine.
Type species. Munneckella tribuscamera sp. nov., the only known species.
Original diagnosis (Modified from Munnecke et al., 2000). Ovummuridae characterised by a three-chamber organisation. The morphological differences include variation in the thickness of the wall, the two pseudo-septal structures and the shape of the three chambers. The three chambers can be symmetrical and asymmetrical in size. The wall is a lamellar arrangement of calcite tablets. There are two pseudo-septums present and a cylindrical aperture can be present in either pole of the test.
Remarks. This morphotype differs from Ovummurus and Minourella in showing three (not two) chambers, divided by two pseudo-septa as originally described by Munnecke et al. (2000). The variation in chamber size between the three chambers suggests the potential for erecting further species. This could potentially be a different life cycle stage as suggested by MacNeil and Jones (2006), however, the number of observations, and the lack of any transitional evidence found, lead us to conclude that they to belong to a new species.
The genus can only be compared to a limited number of microfossils; three-chambered calcareous microfossils are rare. In cross section, foraminifera such as: Globorotalia centralis Cushman and Bermudez, 1937, can appear three-chambered, however, Globorotalia centralis are agglutinated, and their dimensions, microstructure and number of individual chambers are completely different from those observed in Munneckella (Tappan and Loeblich, 1988). Ovummuridae are distinctive for the lamellar arrangement of calcite tablets in their walls and are not agglutinated.
Spores such as the Permian aged Platysaccus papilionis Potonié and Klaus, 1954, Lueckisporites virkkiae Potonié and Klaus, 1954 and Striatopodocarpites fusus (Balme and Hennelly, 1955) Potonié, 1958 exhibit the most similar geometry to the three-chambered organisation of Munneckella (Balme and Hennelly, 1955 and Potonie, 1958). However, spores such as: Striatopodocarpites fusus have bisaccate morphology in which the three compartmentalisation is a product of wing-like features rather than three chambers as observed within Munneckella. The walls observed in Muneckella are also calcareous, and its structural arrangement is different to those of spores. Nannoplankton such as coccolithophores (e.g., Zygodiscaceae, Hay and Mohler, 1967) can have a similar three-chamber organisation as Munneckella. Isthmolithus recurves Deflandre and Fert, 1954, is characteristic of two parallel transverse bars that form a three-compartment organisation. Although morphologically similar, these nannofossils do not share the same wall structure.
Stratigraphical range. Lower Silurian to Upper Devonian.
Munneckella tribuscamera sp. nov.
Figure 7A-D
zoobank.org/9075F00B-3A8D-4ED3-A416-8EC8EBBB1ABD
Derivation of name. Latin Tribus, three and camera - chamber, vault; genus neutral.
 Holotype. 15MQ/0001, Moorwood Quarry, Crinoidal Grainstones of the Off-reef Tract, Much Wenlock Limestone Formation, Homerian.
Holotype. 15MQ/0001, Moorwood Quarry, Crinoidal Grainstones of the Off-reef Tract, Much Wenlock Limestone Formation, Homerian.
Paratype. 17HH/0187, Harton Hollow, Crinoidal Grainstones of the Off-reef Tract, Much Wenlock Limestone Formation, Homerian.
Material. 44 specimens.
Diagnosis. same as for genus.
Description. Oval test split into three asymmetrical chambers. The large central chamber is oval to spherical in shape and is separated from the adjacent chambers by two pseudo-septa. The smaller peripheral chambers are positioned at either polar end of the large central chamber and are oval in shape. The apex of the chambers is smooth. Apertures may occur within the posterior of either, or both, of the smaller chambers. The wall of the test encompasses all three chambers and is characteristic of calcite tablets in a lamellar arrangement. Detrital clasts were observed sporadically within the chambers.
Dimensions. Measurements by Munnecke et al. (2000) gave a test height of 25-140 μm and a width, 20-90 μm. The specimens from the MWL Fm. measured a test height of 25.8-102.0 μm and a test width of 16.1-52.9 μm long. The large dorsal chamber height of 4.8-101.0 μm and a diameter of 8.1-50.0 μm with a surrounding wall thickness of 1.6-16.1 μm. The smaller peripheral chambers have a height of 1.0-96.8 μm and a width of 1.0-52.9 μm and the thickness of the wall surrounding the chamber is 1-8.7 μm thick. A septum thickness of 1.6-17.3 and an aperture width ranging from 0.9-1.9 μm.
Remarks. Measured specimens from the MWL Fm. are comparably smaller than those observed from Gotland. Variation in thickness is observed within the three-chambered variants, in particular a thicker pseudo-septum is observed. Chamber sizes vary with some specimens having equal-sized chambers and other specimens having asymmetrical chamber morphologies. There is a possibility of erecting further species based on the size variations of the chambers.
Stratigraphical range. Lower Silurian (Homerian) - Devonian (Frasnian).
Open Nomenclature
Gen. et. Sp. Indet. 1
Figure 8A
Material. 6 specimens.
Description. A bilocular test comprising of one large and one small chamber. The larger dorsal chamber has a sunken apex (heart-shaped). The smaller chamber is conical.
 Dimensions. Test measurements range from: height of 61.3-71.9 μm, width of 63.2-82.7 μm. Dorsal chamber height: 35.5-48.4 μm, diameter: 58.8-71.2 μm, Thickness of the wall surrounding the dorsal chamber: 2.9-4.8 μm. Septum thickness: 1.6-3.8 μm.
Dimensions. Test measurements range from: height of 61.3-71.9 μm, width of 63.2-82.7 μm. Dorsal chamber height: 35.5-48.4 μm, diameter: 58.8-71.2 μm, Thickness of the wall surrounding the dorsal chamber: 2.9-4.8 μm. Septum thickness: 1.6-3.8 μm.
Remarks. Similar to Minourella wenlockensis with its conical ventral chamber however the apex of the large dorsal chamber is slightly sunken. Due to specimen numbers, this morphotype has been left to open nomenclature because only six specimens were found and the full range of morphological variation, therefore, remains unknown. The observations could also possibly have been due to cross-sectional cuts.
Gen. et. Sp. Indet. 2
Figure 8B
Material. 2 specimens
Description. A bilocular prismatic test compartmentalised into two asymmetrical chambers. The larger dorsal chamber is pentagonal. The ventral chamber is conical.
Dimensions. Test measurements range from: height of 59.6-67.3 μm, width of 43.3-65.4 μm. Thickness of the wall surrounding the dorsal chamber: 2.9-4.8 μm. Septum thickness: 2.9-3.8 μm.
Remarks. The variant appears cubic in cross-section and could be a specimen of Natantesprifmata rogersi, however, they are cubic rather than prismatic. This morphotype has been left to open nomenclature because only two specimens were found and the full extent of the morphological variation still remains unknown.
Gen. et. Sp. indet. 3
Figure 8C
Material. 20 specimens
Description. A bilocular pyramidal test that comprises one large and one small chamber. Both the dorsal and ventral chambers are conical. The pseudo-septum separates the two chambers. No apertures were observed.
Dimensions. Test measurements range from: height of 51.0-96.8 μm, width of 60.6-96.2 μm. Dorsal chamber height: 14.4-86.6 μm, diameter: 33.7-82.7 μm, Thickness of the wall surrounding the dorsal chamber: 1.0-9.6 μm. Ventral chamber height: 1.9-38.4 μm, diameter: 1.9-38.4 μm, Thickness of the wall surrounding the ventral chamber: 1.0-6.5 μm. Septum thickness: 1.0-8.0 μm.
Remarks. This morphotype differs from any Ovummuridae specimens observed or described. Specimens may potentially be transverse cross sectional cuts through Minourella cameroni. As there is so much variation and not great enough similarity to known specimens we have left this to open nomenclature.
Gen. et. Sp. indet. 4
Figure 8D
Material. 4 specimens
Description. A bilocular test that is open ended at its posterior. The larger dorsal chamber is open at its posterior rather differently than other apertures observed from Ovummuridae. The smaller ventral chamber can be conical or globular.
Dimensions. Test measurements range from: height of 86.6-192.4 μm, width of 48.1-74.1 μm. Thickness of the wall surrounding the dorsal chamber: 1.0-5.7 μm, thickness of the wall surrounding the ventral chamber: 1.0-3.8 μm. Septum thickness: 1.0-5.7 μm.
Remarks. Similar to Minourella gotlandica and Minourella wenlockensis. Left to open nomenclature as unsure if observations are due to cross-sectional cuts, however, the wall flares out to produce flanges that have a much wider opening than typical Ovummuridae apertures, these may not be due to cross-sectional cuts.
Gen. et. Sp. indet. 5
Figure 8E-F
Material. 24 specimens
Description (Modified from MacNeil and Jones, 2006). Globular Ovummuridae characterised by one small ventral chamber and one large dorsal chamber. Septum partly divides the large dorsal chamber into two sub-chambers, connected by a simple aperture. The septum is undulating and protrudes into the dorsal chamber to form an arcuate member that fully or partially splits the test into two chambers.
Dimensions. MacNeil and Jones (2006) noted measurements of this ovummurid variant to measure: overall test height of: 135-200 μm high, 85-130 μm wide, wall 9-14 μm thick; small chamber 15-25 μm high, 25-40 μm wide; large chamber 95-145 μm high, 70-105 μm wide; septum 50-75 μm long; septum. The specimens from Wenlock Edge range from: height of 33.9-250.5 μm and width of 45.2-298 μm. Dorsal chamber height: 22.6-230.9 μm and diameter: 27.9-282.3 μm. The thickness of the wall surrounding the dorsal chamber: 1.0-10.6 μm. A ventral chamber height: 3.8-19.4 μm and diameter: 3.2-43.3 μm. The thickness of the wall surrounding the ventral chamber: 1.0-8.1 μm. Septum thickness: 1.0-8.1 μm.
Remarks. These specimens are distinct from M. gotlandica because of their larger test size and partial division of their dorsal chamber by a prominent septum. Their globular test shape distinguishes them from M. cameroni, which is conical. The Wenlock Edge material in comparison to the Hay River specimens is comparably larger. They have been left under open nomenclature because the full range of morphological variation still remains unknown.
DISCUSSION
A total of 6591 specimens of Ovummuridae from the MWL Fm., exposed around Wenlock Edge are reported in this study. From the specimens observed, several previously known and several new morphotypes of Ovummuridae were noted. The new morphologies were observed in great numbers and exhibit obvious morphological variations from the Ovummuridae taxa already established such as: Minourella gotlandica and Ovummurus duoportius (Minoura and Chitoku, 1979 and Munnecke et al., 2000). These new morphologies did not fit within the range of measurement dimensions or morphologies previously noted by Minoura and Chitoku (1979), Munnecke et al. (2000) and MacNeil and Jones (2006). This resulted in the establishment of three new genera; Munneckella, Hartonella and Natantesprifmata and four new species: Munneckella tribuscamera, Hartonella oblonga, Natantesprifmata rogersi and Minourella wenlockensis.
Observations of several Ovummuridae species within the off-reef tract of the MWL Fm. are indicative of a diverse ecosystem environment within an open carbonate platform (Ray et al., 2010). There is a clear abundance of Ovummuridae towards the off-reef tract with a large and diverse population of ovummurids observed. The MWL Fm. comprises three lithofacies: Tabular Wackestones and Mudstones, Nodular/Bioclastic Packstones and Crinoidal Grainstones; deposited in three slightly different shallow marine environments. Our results show ovummurids were observed in the greatest numbers in the Nodular/Bioclastic Packstone, relative to the other lithofacies. In prior studies, it had been noted by Blackburn (2016) that no ovummurids were observed within the tabular lithofacies, however, by using the reflected light methodology of Hussain et al. (2019), a total of 476 specimens have since been found. Similarly, MacNeil and Jones (2006) note that their observations suggest ovummurids preferred off-reef environments. It is possible that the greater numbers of Ovummuridae observed within the Nodular/Bioclastic Packstone facies is influenced by cementation, diagenetic alteration or differences in hydraulic concentration of these allochthonous organisms, however, no obvious evidence of these processes has been noted. We, therefore, propose that this is a valid representation for the distribution of Ovummuridae in the MWL Fm.
BIOLOGICAL AFFINITIES AND PHYLOGENETIC RELATIONSHIPS OF OVUMMURIDAE
Various observations of Ovummuridae have provided some insight into possible phylogenetic relations that the species may have. Previously, the affinity of Ovummuridae had been compared to many foraminifera (e.g., parathuramminoids, calipionellids and Calcisphaerulida sensu Flügel) by Munnecke et al. (2000). These comparisons were based on the similarities in morphology, but it is highlighted that the Ovummuridae have distinctive walls formed of calcite tablets. The Ovummuridae have also been compared to dinoflagellate cysts based on similarities of the ultrastructure of the walls by Servais et al. (2009).
A possible taxonomical relation to bryozoans is suggested from the results of this study. On the basis of their distinctive wall structure. Ovummuridae are characteristic for their lamellar arrangement of calcite tablets in the ultra-structure of their walls; resembling that of a brick-wall (Minoura and Chitoku, 1979; Munnecke et al., 2000). When compared to the walls of a bryozoan, they are very similar and many observed ovummurid specimens are in close proximity to bryozoans. It is possible that they could be bryozoan polyps, and although this does not explain the range of morphotypes, it could explain their planktonic mode of life.
Minoura and Chitoku (1979) note fine-grained clastic material within the chambers and sometimes within the septum-like structure of Ovummuridae specimens. They suggest a potential relationship with encrusting algae, which have been known to grow on clastic material. MacNeil and Jones (2006) similarly compared the lifestyle of Ovummuridae to an encrusting algae based on quartz grains within the internal chambers of several specimens. Clastic grains were observed within the chambers of some specimens studied here, but not to such an extent that it would be considered a characteristic feature.
STRATIGRAPHIC DISTRIBUTION
The stratigraphic distribution of the Ovummuridae stretches from the lower Silurian to the Upper Permian (Munnecke et al. (2001). The majority of specimens have been found from Silurian material (including the several thousand reported here), this is likely due to the excellent preservation of carbonates from Gotland and Wenlock Edge (UK). The numbers of Ovummuridae in this study support the suggestion of Munnecke et al. (2001) that these microfossils were possibly abundant in carbonate platforms throughout the Palaeozoic, but are so far largely underreported.
Observing new and existing species of Ovummuridae from the MWL Fm. requires the adjustment of the known stratigraphical extent of the family, its genera and species. This study has added four new species from the Homerian (Silurian) and has increased the known stratigraphical distribution of the previously reported species Minourella cameroni (MacNeil and Jones, 2006). M. cameroni was first noted within the Late Devonian carbonate ramps of the Upper Escarpment and Alexandra Fm. in the Hay River deposits (MacNeil and Jones, 2006). Previously there had been no additional observations of this species, and this is the first observation of M. cameroni in the MWL Fm. outside its type locality; a total count of 27 specimens (if cross-sectional cuts are taken into consideration) were noted. This, therefore, extends the known stratigraphical range of M. cameroni from a first appearance in Homerian, (Wenlock) Silurian to last known appearance in the Frasnian-Famennian, Upper Devonian.
The stratigraphical extent of A. pluricellata remains Homerian (Wenlock) Silurian in age, but its palaeogeographical range now includes the Silurian of Wenlock Edge. S. circumcamerata has a slight stratigraphical range increase from Sheinwoodian (lower Wenlock) to Homerian (upper Wenlock).
COMPARISON TO THE SILURIAN OF GOTLAND: AN INTERPRETATION OF PALAEOGEOGRAPHY
The Silurian of Gotland during the Homerian was situated close to the equator and was covered by a warm, epicontinental sea known as the Baltic Basin forming a series of stacked carbonate platforms (Poprawa et al., 1999 and Calner et al., 2004). The warm waters of the Baltic Basin accommodated reefs that formed the majority of the rock formations that form the island today (Calner et al., 2004). The MWL Fm. is equivalent to the upper Slite, Mulde/Halla and Klinteburg beds of the Silurian of Gotland strata and are of Homerian of age (Munnecke et al., 2000 and Calner et al., 2004). The sedimentology of the MWL Fm. in comparison to the Gotland beds is very similar, in which the Slite Beds are equivalent to the Tabular lithofacies, Mulde/Halla beds are equivalent to the Nodular Lithofacies and the Klinteberg Beds are equivalent to the Crinoidal Grainstone Lithofacies (Calner et al., 2004; Blackburn, 2016).
The findings of Munnecke et al., (2000) compared to our findings from Wenlock Edge show a very similar dominance of M. gotlandica as well as the presence of O. duoportius. Munnecke et al. (2000) reported seven morphotypes of Ovummuridae; this study recognises 10 morphotypes within the MWL Fm. The limestones of the MWL Fm. contain the same ovummurid diversity as those of Gotland with observed species including: A. pluricellata, S. circumcamerata and the three-chambered variant now established as M. tribuscamera. We note lower numbers of A. pluricellata and S.circumcamerata but a higher population of M. tribuscamera (n=44) at Wenlock Edge. Additional taxa that are not observed from Gotland are also reported here. M. cameroni was previously only reported from the Late Devonian deposits of Canada by MacNeil and Jones (2006). In addition, a new taxon, Natantesprifmata (kite-shaped morphology) is present in the MWL Fm.
Munnecke et al., (2000) found M. gotlandica within the Slite, Klinteburg, Mulde and Halla beds and reported observations of six specimens of O. duoportius within the overlying Hemse beds (Gorstian, Ludlow) and underlying Upper Visby (Sheinwoodian). A. pluricellata was found within the Slite beds but also the Upper Visby beds and Slite beds and report only finding S. circumcamerata in the Upper Visby Beds (Munnecke et al., 2000). In contrast, almost all species were present throughout all three lithofacies within the MWL Fm. The tabular lithofacies of the MWL Fm. are barren of M. cameroni but do contain S. circumcamerata. Whilst the crinoidal lithofacies lack S.circumcamerata, they are abundant with specimens of M. cameroni. O. duoportius within the MWL Fm. appear continuously throughout the lithofacies, which does not seem to occur within the Homerian beds of Gotland where they are reported from within Hemse and Upper Visby beds (Munnecke et al., 2000).
This variation in assemblage could be due to palaeoenvironmental, palaeoecological and/or taphonomic factors. Munnecke et al., (2001) note Ovummuridae have a planktonic mode of life as they can survive within a range of environments recognised from the strata of Gotland from extremely shallow marine lagoonal environments to distal shelf settings within the photic zone. The MWL Fm. deposits are from an off-reef environment similar to the environment recognised by MacNeil and Jones (2006) for the Hay River material.
Populations of ovummurid species that are observed from Gotland: M. gotlandica, O. duoportius, A. pluricellata and S. circumcamerata and M. tribuscamera seem to be found around the Baltic Basin stretching towards the Avalonian shelf margin where Wenlock Edge was situated and explains why observations of the same population are seen. Whereas, the populations of ovummurids that are only observed at Wenlock Edge and not in Gotland include: M. wenlockensis, M. cameroni, N. rogersi and H. oblonga and are found to dominate the Avalonia shelf margins. The continental collision of Laurentia and Avalonia and Baltica during the Caledonian orogeny (Ordovician - Early Devonian) increased the area of marine-flooded continental shelf and could possibly explain the observations of M. cameroni within the Hay River Material (situated in Laurussia) at the end of the Devonian (Cocks and Torsvik, 2002; MacNeil and Jones, 2006).
The platform was a ramp type that became steeper through time and eventually transformed into a distally steepening ramp (Calner et al., 2004). By contrast, The MWL Fm. was deposited on the Midland platform as an open carbonate shelf (Wright and Burchette, 1996 and Ratcliffe and Thomas, 1999). The difference in ovummurid assemblages observed between the MWL Fm. and the Silurian of Gotland could possibly be related to the palaeogeographical location and their varying carbonate environments. Alternatively, observation methodology may have played a role in the number and diversity of Ovummuridae observed. Here we used a reflected light microscopy method (Hussain et al., 2019) with very little SEM, MacNeil and Jones (2006) used mixed transmitted light microscopy and SEM and Munnecke et al. (2001) used exclusively SEM.
CONCLUSION
Based on a novel application of reflected light microscopy of polished and etched thin sections, this study introduces a detailed report of the Ovummuridae of the MWL Fm. 6951 individual specimens were observed, highlighting the usefulness of the cheap and efficient reflected light microscopy approach used. Several new taxa are introduced and further information on most known Ovummuridae provided, namely:
(1) The establishment of three new genera based on observations of new ovummurid morphotypes; Natantesprifmata, Hartonella and Munneckella and subsequently four new species: Natantesprifmata rogersi, Munneckella tribuscamera, Hartonella oblonga and Minourella wenlockensis.
(2) The first observations and therefore the first reporting of Minourella cameroni and Arouxina pluricellata outside of their type locality; extending the known stratigraphical range of M. cameroni to the Homerian (Wenlock), Silurian.
(3) Adjustment of the biostratigraphical distribution of Minourella cameroni from previously known from the Late Devonian to now having a stratigraphical range from the Homerian (Silurian) to the Frasnian (Late Devonian).
(4) The abundance and diversity of Ovummuridae reported from the off-reef tract of the Wenlock carbonate platform (and the relative reduction within the reef tract) indicates these microfossils most likely had an affinity to this environment.
The large number of specimens observed in this study, indicate that the method deployed here is both time and cost effective, however, images produced are often less clear or well-defined. The common occurrence of Ovummuridae from the Much Wenlock Limestone Formation, a “classic” and well-studied formation, highlights the importance of revisiting these lithologies.
ACKNOWLEDGEMENTS
The authors would like to thank D. Wilde of Keele University for his expertly crafted thin sections and technical assistance. T. Servais and an anonymous reviewer are thanked for their constructive and enthusiastic reviews. A. Jeffery and J. Pringle are thanked for reading over a draft of the manuscript. The author S.L. Rogers would like to highlight that R. Hussain erected the taxonomic names outlined in this work in her master’s thesis. S.L. Rogers had no input in the choice of names and is very humbled.
REFERENCES
Balme, B.E. and Hennelly, J.P.F. 1955. Bisaccate sporomorphs from Australian Permian coals. Australian Journal of Botany, 3(1):89-98. https://doi.org/10.1071/BT9550089
Bassett, M.G. 1976. A critique of diachronism, community distributions and correlation of the Wenlock-Ludlow boundary. Lethaia, 9:207-18. https://doi.org/10.1111/j.1502-3931.1976.tb00967.x
Bassett, M.G. 1989. The Wenlock Series in the Wenlock area, p. 51-73. In Holland, C.H. and Bassett, M.G. (eds.), A global standard for the Silurian System. Cardiff: National Museum of Wales, Geological Series no. 9.
Biffi, U. and Grignani, D. 1983. Peridinioid dinoflagellate cysts from the Oligocene of the Niger Delta, Nigeria. Micropaleontology, 29:126-45. https://doi.org/10.2307/1485563
Blackburn, J. 2016. Sedimentology and depositional environment of the off-reef Much Wenlock Limestone Formation in Shropshire and Herefordshire, UK. Keele University. (MSc Dissertation).
Calner, M., Jeppsson, L., and Munnecke, A. 2004. The Silurian of Gotland--Part I: Review of the stratigraphic framework, event stratigraphy, and stable carbon and oxygen isotope development. Erlanger geologische Abhandlungen, Sonderband, 5:113-131.
Cocks, L.R.M. and Torsvik, T.H. 2002. Earth geography from 500 to 400 million years ago: a faunal and palaeomagnetic review. Journal of the Geological Society, 159(6):631-644. https://doi.org/10.1144/0016-764901-118
Crosfield, M.C. and Johnston, M.S. 1914. A study of Ballstone and the associated beds in the Wenlock Limestone of Shropshire. Proceedings of the Geologists' Association, 25(3):193-225. https://doi.org/10.1016/S0016-7878(14)80051-5
Cushman, J.A. and Bermúdez, P.J. 1937. Further new species of foraminifera from the Eocene of Cuba. Contributions from the Cushman Laboratory for Foraminiferal Research, 13(1):1-29.
Cushman, J.A. and Waters, J.A. 1928. Upper Paleozoic Foraminifera from Sutton County, Texas. Journal of Paleontology, 2(4):358-371.
Deflandre, G. and Fert, C. 1954. Observations sur les coccolithophoridés actuels et fossiles en microscopie ordinaire et électronique. Annales de paléontologie, 1:59,
Eisenack, A. 1937. Neue Mikrofossilien des baltischen Silurs. IV. Palaeontologische Zeitschrift, 13:74-118
Fry, C.R., Ray, D.C., Wheeley, J.R., Boomer, I., Jarochowska, E., and Loydell, D.K. 2017. The Homerian carbon isotope excursion (Silurian) within graptolitic successions on the Midland Platform (Avalonia), UK: implications for regional and global comparisons and correlations. GFF, 139(4):301-313. https://doi.org/10.1080/11035897.2017.1388280
Haeckel, E. 1862. “Die” Radiolarien (Rhizopoda radiaria): eine Monographie. Hauptbd (Vol. 2). Reimer.
Haeckel, E. 1881. Metagenesis und Hypogenesis von Aurelia aurita: Ein Beitrag zur Entwickelungsgeschichte und zur Teratologie der Medusen. G. Fischer.
Haeckel, E. 1887. Report on the Radiolaria collected by HMS Challenger during the years 1873-1876. Report of the Voyage of HMS Challenger.
Hay, W.W. and Mohler, H.P. 1967. Calcareous nannoplankton from early Tertiary rocks at Pont Labau, France, and Paleocene-early Eocene correlations. Journal of Paleontology, 41(6):1505-1541.
Holland, C.H., Lawson, J.D., and Walmsley, V.G. 1963. The Silurian rocks of the Ludlow district, Shropshire. Bulletin of British Natural History Museum, series geology, 8:93-171
Hurst, J.M. 1975. The diachronism of the Wenlock Limestone. Lethaia, 8:301-14. https://doi.org/10.1111/j.1502-3931.1975.tb00935.x
Hussain, R., Rogers, S., and Blackburn, J.A. 2019. Reflected Light Microscopy as an Efficient and Cost-effective Method for the Detection of Calcareous Micro-fossils, An Example from the Much Wenlock Limestone Formation, Shropshire, UK. The Student Journal of Natural Sciences. https://doi.org/10.21252/fpdf-8j55
Larsson, K. 1979. Silurian tentaculitids from Gotland and Scania. Fossils and Strata, 11:1-180.
Lazarus, D. 1986. Tempo and mode of morphologic evolution near the origin of the radiolarian lineage Pterocanium prismatium. Paleobiology, 12(2):175-189.
Loeblich, A.R. and Tappan, H. 1964. Foraminiferal classification and evolution. Journal of Geological Society of India, 5:5-40.
MacNeil, A.J. and Jones, B. 2006. Ovummuridae (calcareous microfossils) from a Late Devonian ramp: their distribution, preservation potential, and palaeoecological significance. Journal of Earth Sciences. 43(3):269-280. https://doi.org/10.1139/e05-105
Minoura, N. and Chitoku, T. 1979. Calcareous nannoplankton and problematic microorganisms found in the Late Palaeozoic limestones. Journal of the Faculty of Science, Hokkaido University. Series 4, Geology and mineralogy. 19(1-2):199-212.
Munnecke, A. and Servais, T. 1996. Scanning electron microscope of polished, slightly etched rock surfaces: A method to observe polynomorphs in situ. Palynology, 20(1):163-176. https://doi.org/10.1080/01916122.1996.9989473
Munnecke, A., Servais, T., and Vachard, D. 2000. A new family of calcareous microfossils from the Silurian of Gotland, Sweden. Palaeontology, 43(6):1153-1172. https://doi.org/10.1111/1475-4983.00165
Munnecke, A., Servais, T., and Vachard, D. 2001. New findings and stratigraphical distribution of the Ovummuridae (Palaeozoic calcareous microfossils). Earth and Planetary Sciences, 333(3):179-185. https://doi.org/10.1016/S1251-8050(01)01624-X
Murchinson, R.I. 1833. On the sedimentary deposits which occupy the western parts of Shropshire and Herefordshire, and are prolonged from NE to SW, through Radnor, Brecknock and Caermarthenshire, with descriptions of the accompanying rocks of intrusive or igneous characters. Proceedings of the Geological Society of London, 1:470-477.
Poprawa, P., Šliaupa, S., Stephenson, R., and Lazauskien, J. 1999. Late Vendian-Early Palaeozoic tectonic evolution of the Baltic Basin: regional tectonic implications from subsidence analysis. Tectonophysics, 314:219-239. https://doi.org/10.1016/S0040-1951(99)00245-0
Potonié, R. 1958. Synopsis der Gattungen der Sporae dispersae. T. II. Sporites (Nachträge), Saccites, Aletes, Praecolpates. Polyplicates, Monocolpates: Amt für Bodenforschung Beihefte.
Potonié, R. and Klaus, W. 1954. Einige Sporengattungen des alpinen Salzgebirges. Geologisches Jahrbuch, 68:517-546.
Ratcliffe, K.T. 1988. Oncoids as environmental indicators in the Much Wenlock Limestone Formation of the English Midlands. Journal of the Geological Society, London, 145:117-24. https://doi.org/10.1144/gsjgs.145.1.0117
Ratcliffe, K.T. 1991. Palaeoecology, taphonomy and distribution of brachiopod assemblages from the Much Wenlock Limestone Formation of England and Wales. Palaeogeography, Palaeoclimatology, Palaeoecology, 83:265-293. https://doi.org/10.1016/0031-0182(91)90056-W
Ratcliffe, K.T. and Thomas, A.T. 1999. Carbonate depositional environments in the late Wenlock of England and Wales. Geology Magazine, 136(2):189-204. https://doi.org/10.1017/S0016756899002538
Rauzer-Chernousova, D.M. 1948. Some new species of foraminifera from the Lower Carboniferous deposits of the Moscow Basin. Trudy Instituta Geologicheskikh Nauka, Akademiya Nauk SSSR, 62:227-238.
Ray, D.C. and Thomas, A.T. 2007. Carbonate depositional environments, sequence stratigraphy and exceptional skeletal preservation in the Much Wenlock Limestone Formation (Silurian) of Dudley, England. Palaeontology, 50(1):197-222. https://doi.org/10.1111/j.1475-4983.2006.00607.x
Ray, D.C., Brett, C.E., Thomas, A.T., and Collings, A.V.J. 2010. Late Wenlock sequence stratigraphy in Central England. Geological Magazine, 147:123-144. https://doi.org/10.1017/S0016756809990197
Rogers, S., Blackburn, J., and Price, D. 2017. A new observation of Ovummuridae, from the mid-Silurian (Wenlock) strata of Wenlock Edge, Shropshire, UK: A preliminary report. Proceedings of the Shropshire Geological Society,18:74-79.
Servais, T., Munnecke, A., and Versteegh, G.J. 2009. Silurian calcispheres (Calcitarcha) of Gotland (Sweden): comparisons with calcareous dinoflagellates. Comptes Rendus Palevol, 8(6):527-534. https://doi.org/10.1016/j.crpv.2009.04.002
Scoffin, T.P. 1971. The conditions of growth of the Wenlock reefs in Shropshire. Sedimentology, 17:173-219. https://doi.org/10.1111/j.1365-3091.1971.tb01774.x
Shergold, J.H. and Bassett, M.G. 1970. Facies and faunas at the Wenlock/Ludlow boundary of Wenlock Edge, Shropshire. Lethaia, 3:113-142. https://doi.org/10.1111/j.1502-3931.1970.tb01853.x
Tappan, H. and Loeblich Jr., A.R. 1988. Foraminiferal evolution, diversification, and extinction. Journal of Paleontology, 62(5):695-714.
Vachard, D., Krainer, K., and Schönlaub, H.P. 2018. Lower Serpukhovian (Steshevian) foraminifers and algae from exotic limestone clasts of Nötsch (Eastern Alps, Austria). Geobios, 51(1):75-100. https://doi.org/10.1016/j.geobios.2017.12.005
Wright, V.P. and Burchette, T.P. 1996. Shallow-water carbonate environments, p. 325-394. In Reading, H.L., (ed.), Sedimentary Environments: processes, facies, and stratigraphy. Black-well Science, Oxford.
