|
RESULTS
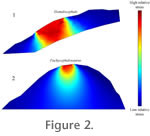 Control: Stress Distribution inSize-Normalized Dome Models Control: Stress Distribution inSize-Normalized Dome Models
Figure 2 depicts results of an analysis with 2D models of cranial dorsa of Homalocephale and Pacycephalosaurus scaled to the same length and with the same impact forces. In the Homalocephale simulation (Figure 2.1), high-magnitude von Mises stresses extend through the dome, reaching the brain cavity (Figure 2.1). In the Pachycephalosaurus analysis, von Mises stresses diminish rapidly away from the point of impact (Figure 2.2), despite application of force to a smaller area than in the flatter-headed Homalocephale.
Stress Distribution in Homalocephale
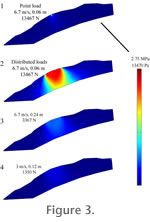 Homalocephale results (Figure 3) reveal that the skull could unambiguously handle impact force at closing speeds of 6.7m/s to 3m/s. The one exception occurred at 6.7m/s with a deceleration distance of 0.06 m. Under these conditions, the skull model was not able to fully dissipate stress in the region dorsal to the brain, resulting in a well-defined area of high tensile stress adjacent to the endocranial cavity. Much higher peak stresses occurred at point impacts than at distributed impacts, but these are focused at the locus of impact and fall below the failure strength of bone. Based on these results, it is reasonable to conclude that Homalocephale would have been able to handle head-butting behavior over most of the velocity and deceleration ranges tested, but that high speed impacts would have been more problematic. Homalocephale results (Figure 3) reveal that the skull could unambiguously handle impact force at closing speeds of 6.7m/s to 3m/s. The one exception occurred at 6.7m/s with a deceleration distance of 0.06 m. Under these conditions, the skull model was not able to fully dissipate stress in the region dorsal to the brain, resulting in a well-defined area of high tensile stress adjacent to the endocranial cavity. Much higher peak stresses occurred at point impacts than at distributed impacts, but these are focused at the locus of impact and fall below the failure strength of bone. Based on these results, it is reasonable to conclude that Homalocephale would have been able to handle head-butting behavior over most of the velocity and deceleration ranges tested, but that high speed impacts would have been more problematic.
Stress Distribution in 3D Pachycephalosaurus Models
 Results for Pachycephalosaurus wyomingensis depict a skull that could withstand high impact forces. Stresses are low with a simple point application of the calculated impact force, and
with much higher applied forces the stress still falls below the limits of bone strength. Results for Pachycephalosaurus wyomingensis depict a skull that could withstand high impact forces. Stresses are low with a simple point application of the calculated impact force, and
with much higher applied forces the stress still falls below the limits of bone strength.
 At 6.7 m/s (with maximum von Mises stress of 14660 Pa:
Figure 4.1) and 3 m/s, stress would dissipate throughout the skull before it would reach the braincase (Figure 4.2). Strain decreased dramatically in magnitude as it was diverted laterally (Figure 4.3) and anteroposteriorly. If high stress was seen to reach the braincase, a high magnitude of stress would be prevalent throughout the images. A similar pattern of stress distribution occurs under the side-impact loading regime (Figure 5), with stress diminishing deep to the site of impact, and anterior and posterior to it. At 6.7 m/s (with maximum von Mises stress of 14660 Pa:
Figure 4.1) and 3 m/s, stress would dissipate throughout the skull before it would reach the braincase (Figure 4.2). Strain decreased dramatically in magnitude as it was diverted laterally (Figure 4.3) and anteroposteriorly. If high stress was seen to reach the braincase, a high magnitude of stress would be prevalent throughout the images. A similar pattern of stress distribution occurs under the side-impact loading regime (Figure 5), with stress diminishing deep to the site of impact, and anterior and posterior to it.
Strain Results for the Subadult Pachycephalosaurine
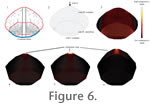 Figure 6.1–6.2 depict the modeled specimen and sagittal section mesh, incorporating histological zones I, II, and III, of the subadult pachycephalosaurine.
Figure 6.3 shows the distribution of compressive strain in the model. Cancellous zone II shows higher overall compressive strain than either of the contiguous zones of compact bone. Figure 6.1–6.2 depict the modeled specimen and sagittal section mesh, incorporating histological zones I, II, and III, of the subadult pachycephalosaurine.
Figure 6.3 shows the distribution of compressive strain in the model. Cancellous zone II shows higher overall compressive strain than either of the contiguous zones of compact bone.
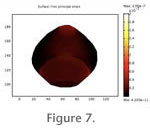 Differences in strain magnitude between zones are especially evident in when the results are animated (Figure 7). Differences in strain magnitude between zones are especially evident in when the results are animated (Figure 7).
Greater strain in the cancellous zone also results in models with varying thicknesses of keratinous integument. However, strain within all three zones decreases when the load is applied to the integument (as would occur in life), and magnitudes of bone strain are inversely proportional to modeled thickness of the keratinous covering (Figure 6.4-6.6).
Phylogenetic Results and Character Optimization
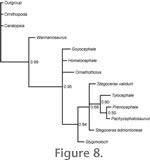 Figure 8 depicts the 50% majority rule consensus tree from the phylogenetic analysis. The topology is similar to that presented by
Maryánska et al. (2004), with a polytomy for Goyocephale, Homalocephale, Ornatotholus, and a clade consisting of more derived taxa. Relationships within this clade are similar to those reported in the consensus tree of
Maryánska et al. (2004), but the seemingly high posterior probability values are low by molecular phylogenetics standards. Figure 8 depicts the 50% majority rule consensus tree from the phylogenetic analysis. The topology is similar to that presented by
Maryánska et al. (2004), with a polytomy for Goyocephale, Homalocephale, Ornatotholus, and a clade consisting of more derived taxa. Relationships within this clade are similar to those reported in the consensus tree of
Maryánska et al. (2004), but the seemingly high posterior probability values are low by molecular phylogenetics standards.
 The negligible 50% posterior probability support for a Pachycephalosaurus-Prenocephale grouping indicates a polytomy between these taxa and Tylocephale, as recovered by
Maryánska et al. (2004). The negligible 50% posterior probability support for a Pachycephalosaurus-Prenocephale grouping indicates a polytomy between these taxa and Tylocephale, as recovered by
Maryánska et al. (2004).
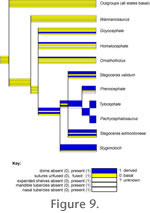
Figure 9 maps characters related to dome mechanical and species recognition functions onto the consensus cladogram. Characters identifiable as for species recognition, such as an elaborate posterior cranial shelf and cranial and mandibular tubercles, generally increase in prominence in taxa with greater vaulting and addition of other bones to the frontoparietal dome. Stygimoloch is a notable exception to the trend. The
pattern provides tentative evidence against the hypothesis that the dome became reduced as other species recognition features evolved, except in the case of Stygimoloch.
|