| |
OSTEOLOGY
The osteology of Sphenodon was first described in detail by
Günther (1867) but numerous studies have been made subsquently. Because Sphenodon was once seen as a "basic" or "generalised" amniote,
Romer (1956) used it as his point of reference for a wide range of extinct and extant amniotes. Similarly, as the only extant rhynchocephalian, the skull of Sphenodon is often represented in text books (e.g.,
Kardong 1998, p. 250;
Liem et al. 2001, p. 252). The skull structure of hatchling or near hatchling animals (skull length = 5–15 mm) was investigated by
Howes and Swinnerton (1901),
Werner (1962), and
Rieppel (1992) whereas
Osawa (1898) and, more recently,
Evans (2008) described skulls of animals that were not fully grown. Ten skulls of different size were figured by
Jones (2008), and geometric morphometrics showed that skull growth in Sphenodon is allometric because during ontogeny the postorbital part of the skull expands relative to the orbital and preorbital portions. Several descriptions have also been made of the braincase (Siebenrock 1893,
1894;
Osawa 1898;
Gower and Weber 1998;
Evans 2008), the teeth (Gray 1831;
Günther 1867;
Colenso 1886;
Harrison 1901a,
1901b;
Howes and Swinnerton 1901;
Robinson 1976;
Throckmorton et al. 1981;
Gorniak et al. 1982;
Rieppel 1992;
Jones 2006a,
2006b,
in press), and the axial skeleton (e.g.,
Günther 1867;
Howes and Swinnerton 1901;
Hoffstetter and Gasc 1969). The cranial joints have been examined by
Jones (2006a,
2007) but this will not be discussed in detail here.
The description that follows is based on previous literature but also from examination of numerous specimens from over a dozen collections:
• Auckland Museum, New Zealand (AIM).
• Berlin Museum of Natural History (Museum für Naturkunde) of the Humbold-University, Germany (ZMB).
• Birkbeck College, School of Biological and Chemical Sciences, London, UK (BRK).
• Booth Museum of Natural History, Brighton, UK (BMB).
• Canterbury Museum, Christchurch, New Zealand (CMC).
• Ernst Moritz Arndt University of Greifswald, Zoology Collection, Germany.
• Grant Museum of Zoology, UCL, London, UK (LDUCZ).
• Huntarian Museum and Art Gallery, University of Glasgow, UK (GLAHM).
• Kings College London, Life Sciences, London, UK (KCL).
• The Manchester Museum, University of Manchester, Manchester, UK (MANCH).
• Natural History Museum, London, UK (BMNH).
• Oxford Museum of Natural History, Oxford, UK (OUMNH).
• David Gower Personal Collection, NHM, UK (DGPC).
• University Museum of Zoology, Cambridge, UK (UMZC).
• The Field Museum, Chicago, USA (FMNH).
• Museum of New Zealand Te Papa Tongarewa (Wellington), New Zealand (NMNZ).
• University of Auckland, New Zealand (AU).
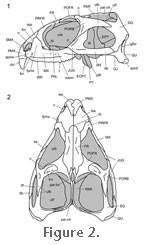 The skull of an adult Sphenodon possesses a short snout, large orbits, and a relatively large postorbital area (Figure 2), and is, in general, approximately 60 mm long from the tip of the rostrum to a point level with the jaw joints (Günther 1867;
Jones 2008). The lacrimal bone is absent (contra
Günther 1867 and
Anderson 1936) in contrast to many squamates where it borders the anterior part of the orbit (e.g.,
Jollie 1960;
Gauthier et al. 1988;
Evans 2008). Correspondingly, in Sphenodon the lacrimal canal runs between the prefrontal and maxilla (Figure 2.1). Note that the term 'lacrimal canal' is still used despite there being no lacrimal gland (e.g.,
Underwood 1970). Above the tooth row is a band of hard tissue referred to here as secondary bone (Harrison 1901a,
1901b;
Jones 2006a,
2006c), and the jaw joint is situated ventral to the long axis of the tooth row (Jones 2008). Both lower and upper temporal fenestrae are present and bounded by lower and upper temporal bars (Günther
1867). The former is composed of the postorbital and squamosal ("mastoid" of
Günther 1867;
Osawa 1898) whereas the latter is composed primarily of the posterior process of the jugal ("zygomatic" of
Günther 1867) with smaller contributions from the squamosal and quadratojugal. In contrast to the prevalent view, current evidence indicates that the lower temporal bar in Sphenodon has been secondarily acquired and does not represent the plesiomorphic condition for lepidosaurs (Whiteside 1983,
1986;
Fraser 1988;
Rieppel 1993;
Reynoso 2000;
Apesteguía and Novas 2003;
Müller 2003;
Wu 2003;
Evans 2003,
2008;
Jones 2006a,
2006c,
2008; contra
Herrel et al. 1998,
2007;
Kardong 1998, p. 249;
Hildebrand and Goslow 2001, p. 129;
Liem et al. 2001, p. 90;
Pough et al. 2005, p. 343). The skull of an adult Sphenodon possesses a short snout, large orbits, and a relatively large postorbital area (Figure 2), and is, in general, approximately 60 mm long from the tip of the rostrum to a point level with the jaw joints (Günther 1867;
Jones 2008). The lacrimal bone is absent (contra
Günther 1867 and
Anderson 1936) in contrast to many squamates where it borders the anterior part of the orbit (e.g.,
Jollie 1960;
Gauthier et al. 1988;
Evans 2008). Correspondingly, in Sphenodon the lacrimal canal runs between the prefrontal and maxilla (Figure 2.1). Note that the term 'lacrimal canal' is still used despite there being no lacrimal gland (e.g.,
Underwood 1970). Above the tooth row is a band of hard tissue referred to here as secondary bone (Harrison 1901a,
1901b;
Jones 2006a,
2006c), and the jaw joint is situated ventral to the long axis of the tooth row (Jones 2008). Both lower and upper temporal fenestrae are present and bounded by lower and upper temporal bars (Günther
1867). The former is composed of the postorbital and squamosal ("mastoid" of
Günther 1867;
Osawa 1898) whereas the latter is composed primarily of the posterior process of the jugal ("zygomatic" of
Günther 1867) with smaller contributions from the squamosal and quadratojugal. In contrast to the prevalent view, current evidence indicates that the lower temporal bar in Sphenodon has been secondarily acquired and does not represent the plesiomorphic condition for lepidosaurs (Whiteside 1983,
1986;
Fraser 1988;
Rieppel 1993;
Reynoso 2000;
Apesteguía and Novas 2003;
Müller 2003;
Wu 2003;
Evans 2003,
2008;
Jones 2006a,
2006c,
2008; contra
Herrel et al. 1998,
2007;
Kardong 1998, p. 249;
Hildebrand and Goslow 2001, p. 129;
Liem et al. 2001, p. 90;
Pough et al. 2005, p. 343).
Premaxillae, nasals, frontals, and parietals are paired although the seam between the parietals can be indistinct posteriorly (Figure 2.2;
Günther 1867;
Siebenrock 1893,
1894;
Jones 2006a). Anteriorly the seam is interrupted by a lenticular parietal foramen (Günther 1867). In life this accommodates the so-called third-eye (Dendy 1911;
Robb 1977). In adults the medial edges of the parietal combine to form a crest (Günther 1867;
Jones 2008), and the posterolateral margins slope ventrolaterally to form a shelf. The posterolateral process interlocks with the medial process of the squamosal to create a posterior temporal bar. Supratemporal bones are absent (but see
Rieppel 1992).
In about 25% of the specimens examined (n = 40) (e.g., Sphenodon specimens BMNH1844.102911, MANCH C.1206.49, KCL x12, LDUCZ x146;
Jones 2006a) a small fontanelle is present at the junction between the nasals and frontals. The postfrontals are relatively large compared to fossil rhynchocephalians (Jones 2006a) and meet the parietals and frontals medially (Günther 1867). Laterally the postfrontals interlock with the postorbitals (Jones 2006a). The latter bone is a triradiate element that bears a trough on its dorsal surface bounded by two ridges running oblique to the midline axis (Jones 2006a). The strength of the trough and ridges varies intraspecifically, usually (but not always) being better defined in larger skulls (Jones 2006a).
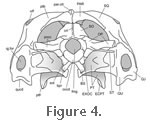
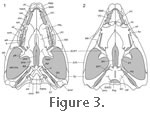 The dorsal portion of the braincase is composed of the prootic, opisthotic, and supraoccipital bones. The supraoccipital bears a posterodorsally projecting crest ("crista supraoccipitalis" of
Siebenrock 1894, p. 310). Ventral to this is the foramen magnum, which is laterally bounded by the exoccipitals. More lateral still, the paroccipital processes of the opisthotics extend towards the posteromedial bases of the squamosals (Gower and Weber 1998) (Figure 3,
Figure 4). The dorsal portion of the braincase is composed of the prootic, opisthotic, and supraoccipital bones. The supraoccipital bears a posterodorsally projecting crest ("crista supraoccipitalis" of
Siebenrock 1894, p. 310). Ventral to this is the foramen magnum, which is laterally bounded by the exoccipitals. More lateral still, the paroccipital processes of the opisthotics extend towards the posteromedial bases of the squamosals (Gower and Weber 1998) (Figure 3,
Figure 4).
 The palate of Sphenodon is vaulted and composed of paired vomers, palatines, and pterygoids (Figure 3.1). All of these bones meet in the midline as visible in dorsal view (Figure 3.2). A lateral process of the pterygoid and ventral process of the ectopterygoid contribute to a substantial pterygoid flange (Günther 1867;
Jones 2006a). The posterior process of the pterygoid laps the medial surface of the quadrate forming a dorsoventrally deep quadrate-pterygoid wing (Günther 1867;
Jones 2006a). Against the lateral surface of the pterygoid, and adjacent to the anterior edge of the quadrate, sits a broad based epipterygoid ("columella" of
Günther 1867). The palate of Sphenodon is vaulted and composed of paired vomers, palatines, and pterygoids (Figure 3.1). All of these bones meet in the midline as visible in dorsal view (Figure 3.2). A lateral process of the pterygoid and ventral process of the ectopterygoid contribute to a substantial pterygoid flange (Günther 1867;
Jones 2006a). The posterior process of the pterygoid laps the medial surface of the quadrate forming a dorsoventrally deep quadrate-pterygoid wing (Günther 1867;
Jones 2006a). Against the lateral surface of the pterygoid, and adjacent to the anterior edge of the quadrate, sits a broad based epipterygoid ("columella" of
Günther 1867).
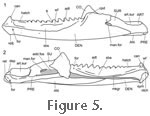 The lower jaw is robust and has a conspicuous mandibular foramen (Günther 1867;
Osawa 1898;
Baur 1891;
Throckmorton et al. 1981). The dentary extends posteriorly to a point level with the articular surface (Figure 5). Posterior to the tooth row the dentary also expands dorsally into the coronoid process (cpd = coronoid process of the dentary,
Jones 2006c) and braces the coronoid bone. The latter is large and contacts both the prearticular and angular ventrally. The lower jaw is robust and has a conspicuous mandibular foramen (Günther 1867;
Osawa 1898;
Baur 1891;
Throckmorton et al. 1981). The dentary extends posteriorly to a point level with the articular surface (Figure 5). Posterior to the tooth row the dentary also expands dorsally into the coronoid process (cpd = coronoid process of the dentary,
Jones 2006c) and braces the coronoid bone. The latter is large and contacts both the prearticular and angular ventrally.
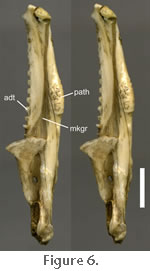 Note that the angular bone was described and labelled as a splenial bone by early authors (e.g.,
Günther 1867;
Osawa 1898,
Baur 1891). The dorsal surface of the articular is saddle-shaped and expanded antero-posteriorly to be about two and a half times the length of the bi-lobate articular surface of the quadrate (e.g., DGPC2). Pathologies on the lower margin of the lower jaw are not infrequent (e.g.,
Figure 6; BMNHD.405; LDUCZ x036, LDUCZ x146; LDUCZ x343; KCL X12;
Robb 1977, figure 5) and probably result from infections following injuries sustained during fighting with conspecifics (Evans 1983;
Daugherty and Cree 1990). On the labial surface of the dentary below the tooth row a skirt of secondary bone develops ontogenetically. This
bony skirt has an obvious ventral edge in adults (Harrison 1901a,
1901b;
Jones 2006a) and compensates for wear from the maxillary teeth (Robinson 1976;
Fraser 1988). Note that the angular bone was described and labelled as a splenial bone by early authors (e.g.,
Günther 1867;
Osawa 1898,
Baur 1891). The dorsal surface of the articular is saddle-shaped and expanded antero-posteriorly to be about two and a half times the length of the bi-lobate articular surface of the quadrate (e.g., DGPC2). Pathologies on the lower margin of the lower jaw are not infrequent (e.g.,
Figure 6; BMNHD.405; LDUCZ x036, LDUCZ x146; LDUCZ x343; KCL X12;
Robb 1977, figure 5) and probably result from infections following injuries sustained during fighting with conspecifics (Evans 1983;
Daugherty and Cree 1990). On the labial surface of the dentary below the tooth row a skirt of secondary bone develops ontogenetically. This
bony skirt has an obvious ventral edge in adults (Harrison 1901a,
1901b;
Jones 2006a) and compensates for wear from the maxillary teeth (Robinson 1976;
Fraser 1988).
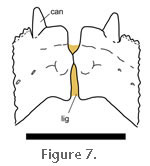 The additional bone also strengthens the lower jaw by increasing its cross-sectional area (Jones 2006a). On the lingual surface is a more planar apron of secondary bone. The symphysial surface is comma-shaped, generally smooth, and located ventral to a notch that has also been found in some fossil rhynchocephalians (Evans et al. 2001). In life the symphysis is maintained by a fibrous ligament that permits some accommodating movement between the left and right lower jaws (Figure 7) (Günther 1867, p. 600;
Robinson 1976, p. 54;
Schwenk 2000, p. 189). The additional bone also strengthens the lower jaw by increasing its cross-sectional area (Jones 2006a). On the lingual surface is a more planar apron of secondary bone. The symphysial surface is comma-shaped, generally smooth, and located ventral to a notch that has also been found in some fossil rhynchocephalians (Evans et al. 2001). In life the symphysis is maintained by a fibrous ligament that permits some accommodating movement between the left and right lower jaws (Figure 7) (Günther 1867, p. 600;
Robinson 1976, p. 54;
Schwenk 2000, p. 189).
The teeth of Sphenodon are fused to the crest of the jaw bone with an acrodont implantation (Günther 1867;
Robinson 1976;
Augé 1997;
Kieser et al. 2008). In adults the premaxilla bears a single large chisel-like tooth bearing two cusps that when worn can superficially resemble the incisor tooth of some mammals (Günther 1867;
Newman 1878;
Howes and Swinnerton 1901;
Robinson 1976). It changes ontogenetically as hatchlings possess three distinct teeth (Günther 1867;
Howes and Swinnerton 1901;
von Wettstein 1931;
Werner 1962;
Rieppel 1992). The anterior part of the maxillary tooth row comprises a set of hatchling teeth that alternate in size (Harrison 1901a,
1901b;
Howes and Swinnerton 1901;
Werner 1962;
Robinson 1976;
Rieppel 1992;
Reynoso 2003). This can be so highly worn in adults that individual teeth are not discernable. Some of the hatchling teeth are also replaced by secondary teeth or caniniforms (Robinson 1976;
Reynoso 2003). The more posteriorly situated dentition consists of adult (or "additional") teeth individually added to the rear of the tooth row during ontogeny. These teeth are larger than the hatchling teeth. They are conical and stout but with a worn medial surface and a posterolingually directed flange (Robinson 1976;
Throckmorton et al. 1981;
Gorniak et al. 1982;
Rieppel 1992;
Jones 2006a).
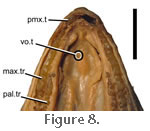 A large row of teeth is present on the lateral margin of the palatine nearly parallel to the teeth on the maxilla (Figure 3.2) so that the dentary teeth fit between the two upper rows when the jaws close. The enlarged palatine tooth row is something
Gray (1831) noted in the first description of Sphenodon (incorrectly identified as an agamid) and represents a unique feature of Rhynchocephalia amongst extant amniotes (Evans 2003;
Jones 2007). The teeth on the palatine are to some extent mirror images of those on the maxilla: conical and stout but with a worn lateral surface and a posterolabial directed flange. Anteriorly juveniles possess a row of hatchling dentition that alternates in size (e.g., OUMNH 700) whereas adults possess a conical caniniform (Robinson 1976;
Jones 2006a). A vestigial tooth may be found on each of the vomers in juveniles and occasionally in adult individuals of Sphenodon (e.g., specimens AMPC1, LDUCZ
x1176 (previously referred to as x804), UCLGMZ x343, BMB100225, OUMNH 4911).
Howes (1890) and
Harrison (1901a, p. 162) did not think these teeth ever entered the oral cavity, but
Siebenrock (1894, p. 310) observed that the teeth of dry specimens were long enough to do so. Examination of specimen BMNH 1972.1223 confirms the suggestion of
Siebenrock (1893,
1894) (Figure 8). A large row of teeth is present on the lateral margin of the palatine nearly parallel to the teeth on the maxilla (Figure 3.2) so that the dentary teeth fit between the two upper rows when the jaws close. The enlarged palatine tooth row is something
Gray (1831) noted in the first description of Sphenodon (incorrectly identified as an agamid) and represents a unique feature of Rhynchocephalia amongst extant amniotes (Evans 2003;
Jones 2007). The teeth on the palatine are to some extent mirror images of those on the maxilla: conical and stout but with a worn lateral surface and a posterolabial directed flange. Anteriorly juveniles possess a row of hatchling dentition that alternates in size (e.g., OUMNH 700) whereas adults possess a conical caniniform (Robinson 1976;
Jones 2006a). A vestigial tooth may be found on each of the vomers in juveniles and occasionally in adult individuals of Sphenodon (e.g., specimens AMPC1, LDUCZ
x1176 (previously referred to as x804), UCLGMZ x343, BMB100225, OUMNH 4911).
Howes (1890) and
Harrison (1901a, p. 162) did not think these teeth ever entered the oral cavity, but
Siebenrock (1894, p. 310) observed that the teeth of dry specimens were long enough to do so. Examination of specimen BMNH 1972.1223 confirms the suggestion of
Siebenrock (1893,
1894) (Figure 8).
The dentary teeth almost invariably demonstrate wear, particularly on their labial and lingual surfaces (Robinson 1976;
Reynoso 1996). The majority of teeth on the dentary are pyramidal with an anteriorly positioned apex that possesses anterolingual and anterolateral flanges (Robinson 1976;
Throckmorton et al. 1981;
Gorniak et al. 1982;
Jones 2006a,
2006b,
2008,
in press,
Jones et al., 2009). However, as for the maxilla and palatine, there is a set of hatchling teeth anteriorly. Again this can be obscured or obliterated by wear in adults and is usually partially replaced by a conical caniniform tooth (Harrison 1901a,
1901b;
Howes and Swinnerton
1901; Robinson 1976;
Reynoso 2003). The anterodorsal tip of each lower jaw may bear an oblique wear facet from where it impacts the back of the corresponding chisel-like premaxillary tooth during the prooral jaw shearing (e.g., OMNH 908; OMNH 4911;
Jones 2006a).
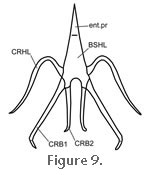 The hyobranchial skeleton is a remnant of the visceral skeleton (splanchnocranium) and is associated with the tongue and throat muscles. The structure in Sphenodon has been described several times (e.g.,
Osawa 1898;
Fürbringer 1922;
von Wettstein 1931,
1932;
Romer 1956;
Tanner and Avery 1982). It comprises a relatively broad midline basihyal composed of cartilage (Figure 9). The basihyal has three anterior projections, of which the median entoglossal process supports the tongue. The two shorter anterolateral processes are connected to a pair of long thin ceratohyals that curve dorsolaterally around the pharynx. Extending posterolaterally from the central body of the basihyal are two pairs of projections: the bony first ceratobranchials, and the shorter and more medially located second ceratobranchials (Osawa 1898;
Fürbringer 1922;
Rieppel 1978;
Tanner and Avery 1982). Comparison of previous illustrations (e.g.,
Osawa 1898;
Fürbringer 1922;
Edgeworth 1935;
Rieppel 1978;
Tanner and Avery 1982) The hyobranchial skeleton is a remnant of the visceral skeleton (splanchnocranium) and is associated with the tongue and throat muscles. The structure in Sphenodon has been described several times (e.g.,
Osawa 1898;
Fürbringer 1922;
von Wettstein 1931,
1932;
Romer 1956;
Tanner and Avery 1982). It comprises a relatively broad midline basihyal composed of cartilage (Figure 9). The basihyal has three anterior projections, of which the median entoglossal process supports the tongue. The two shorter anterolateral processes are connected to a pair of long thin ceratohyals that curve dorsolaterally around the pharynx. Extending posterolaterally from the central body of the basihyal are two pairs of projections: the bony first ceratobranchials, and the shorter and more medially located second ceratobranchials (Osawa 1898;
Fürbringer 1922;
Rieppel 1978;
Tanner and Avery 1982). Comparison of previous illustrations (e.g.,
Osawa 1898;
Fürbringer 1922;
Edgeworth 1935;
Rieppel 1978;
Tanner and Avery 1982)
 suggests that there may be some intraspecific variation in the length of different components. suggests that there may be some intraspecific variation in the length of different components.
The skull and neck are connected by the atlanto-occipital joint which involves both the first and second vertebrae (atlas and axis respectively) (axis = epistropheus of
Günther 1867,
Osawa 1898, and
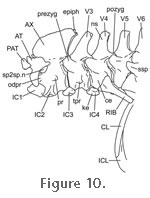
Nishi 1916) (Figure 10).
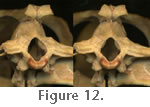
 Also between the occiput and atlas are two small triangular proatlantes (Howes 1890). The atlas consists of an intercentrum and the two halves of the neural arch that extend posteriorly to a zygapophysial joint with the anterodorsal surface of the atlas (LDUCZ x722, LDUCZ x723). The atlas centrum is fused to the axis centrum to form the odontoid process which, in conjunction with the first intercentrum and neural arch bases provides a wide cup-like socket for the ovoid basioccipital condyle (Figure 11 and
Figure 12). Also between the occiput and atlas are two small triangular proatlantes (Howes 1890). The atlas consists of an intercentrum and the two halves of the neural arch that extend posteriorly to a zygapophysial joint with the anterodorsal surface of the atlas (LDUCZ x722, LDUCZ x723). The atlas centrum is fused to the axis centrum to form the odontoid process which, in conjunction with the first intercentrum and neural arch bases provides a wide cup-like socket for the ovoid basioccipital condyle (Figure 11 and
Figure 12).
 The neural spine of the heavily-built axis is greatly expanded antero-posteriorly (Günther 1867;
von Wettstein 1931). The small axis transverse processes are very small and do not bear ribs in adults. The second intercentrum is fused to the axis centrum in adults (Hoffstetter and Gasc 1969; e.g., specimens LDUCZ x036, LDUCZ x723), to form a large saddle-like anterior surface which is convex in the horizontal plane and concave in the vertical plane (Günther 1867, figure 18). This articulates with the atlas intercentrum and allows some rotational movement between the two bones. The neural spine of the heavily-built axis is greatly expanded antero-posteriorly (Günther 1867;
von Wettstein 1931). The small axis transverse processes are very small and do not bear ribs in adults. The second intercentrum is fused to the axis centrum in adults (Hoffstetter and Gasc 1969; e.g., specimens LDUCZ x036, LDUCZ x723), to form a large saddle-like anterior surface which is convex in the horizontal plane and concave in the vertical plane (Günther 1867, figure 18). This articulates with the atlas intercentrum and allows some rotational movement between the two bones.
Post-axis cervical vertebrae (V3-8) possess a centrum that is cylindrical, notochordal (amphicoelous),
has a medioventral ridge, and has expanded anterior and posterior ends (Günther 1867; Hoffstetter and Gasc
1969). The neural arches bear tall neural spines and anterior and posterior zygopophyses (Figure 10) that have a mainly dorso-ventral articulation but there are also small incipient zygosphene-zygantrum joints medially (Hoffstetter and Gasc 1969, p. 237;
Gauthier et al. 1988). Laterally short transverse processes articulate broadly with deep headed ribs (Günther 1867; Hoffstetter and Gasc
1969).
Typically the first ossified rib is attached to the fourth vertebra (Hoffstetter and Gasc
1969) (although part of a double headed rib can be seen in specimen LDUCZ x036 attached to the third vertebra). The rib of the fourth vertebra is small and may be double headed (Hoffstetter and Gasc
1969), single headed (e.g., LDUCZ x036), or may bear a double headed rib on one side only (e.g., YPM 9194; LDUCZ x722). The first ossified rib may also articulate with the intercentrum (Hoffstetter and Gasc 1969;
Al-Hassawi 2004,
2007). The ribs of vertebrae 5, 6, and 7 are also short but have expanded distal ends partly made of cartilage. The rib of the 8th vertebra is long but does not contact the sternum as do the ribs of vertebrae 9 to 12 (Hoffstetter and Gasc
1969). Extending between the ventral surfaces of postaxial vertebrae are smaller intercentra, some of which possess a ventral midline keel (Hoffstetter and Gasc 1969;
Al-Hassawi 2004,
2007).
In adults the pectoral girdle is composed of paired scapulocoracoids, paired clavicles and, ventrally, a slender T-shaped interclavicle (Günther 1867;
Osawa 1898;
Fürbringer 1900;
Howes and Swinnerton 1901;
von Wettstein 1931). Extending from the dorsal edges of the scapulocoracoids are relatively short cartilaginous suprascapulae. Each clavicle attaches to the anterior edge of the scapulocoracoid dorsal to a subtle embayment (Howes and Swinnerton 1901;
Osawa 1898). Medially each clavicle overlaps the anterior edge of the interclavicle (LDUCZ x036, LDUCZ x722).
|