THE TRIGEMINAL MUSCLES
 This section describes the muscles of the skull innervated by the trigeminal nerve (cranial nerve 5 [cn5]). The divisions of the trigeminal nerve (cn5) have long been used to infer the homology of muscular units in the adductor chamber (Luther 1914;
Lakjer 1926;
Oelrich 1956;
Ostrom 1962;
Barghusen 1973;
Haas 1973;
Gomes 1974; Schwenk 2000;
Holliday and Witmer 2007). The trigeminal nerve branches anteriorly into the ophthalmic division (cn5.1), anterolaterally into the maxillary division (cn5.2) and laterally into the mandibular division (cn5.3). The muscle tissue between the mandibular division (cn5.3) and the maxillary division (cn5.2) is referred to as the m. Adductor Mandibulae Externus (mAME); the muscle posterior to the mandibular division (cn5.3) is referred to as the m. Adductor Mandibulae Posterior (mAMP); and the muscle between the ophthalmic division (cn5.1) and maxillary division (cn5.2) is grouped as the m. Adductor Mandibulae Internus (mAMI) (Figure 13). This section describes the muscles of the skull innervated by the trigeminal nerve (cranial nerve 5 [cn5]). The divisions of the trigeminal nerve (cn5) have long been used to infer the homology of muscular units in the adductor chamber (Luther 1914;
Lakjer 1926;
Oelrich 1956;
Ostrom 1962;
Barghusen 1973;
Haas 1973;
Gomes 1974; Schwenk 2000;
Holliday and Witmer 2007). The trigeminal nerve branches anteriorly into the ophthalmic division (cn5.1), anterolaterally into the maxillary division (cn5.2) and laterally into the mandibular division (cn5.3). The muscle tissue between the mandibular division (cn5.3) and the maxillary division (cn5.2) is referred to as the m. Adductor Mandibulae Externus (mAME); the muscle posterior to the mandibular division (cn5.3) is referred to as the m. Adductor Mandibulae Posterior (mAMP); and the muscle between the ophthalmic division (cn5.1) and maxillary division (cn5.2) is grouped as the m. Adductor Mandibulae Internus (mAMI) (Figure 13).
Most of the muscles innervated by the trigeminal nerve comprise the main jaw adductor muscles. These have repeatedly been described in varying degrees of detail (Osawa 1898;
Byerly 1925;
Lakjer 1926;
Edgeworth 1935;
Lightroller 1939;
Anderson 1936;
Poglayen-Neuwall 1953;
Ostrom 1962;
Barghusen 1973;
Haas 1973;
Gorniak et al. 1982;
Wu 2003;
Holliday and Witmer 2007).
Byerly (1925),
Gorniak et al. (1982),
Ostrom (1962),
Haas (1973), and
Wu (2003) report having dissected two specimens whereas
Barghusen (1973) examined one. The account of
Anderson (1936) seems largely to be a translation of
Lakjer (1926).
Holliday and Witmer (2007) also rely on previous literature but did dissect a range of other amniotes including several squamates. Similarly,
Abdala and Moro (2003) coded the cranial muscles of Sphenodon for a cladistic analysis of lepidosaurs based on previous descriptions (citing
Lakjer [1926] and
Haas [1973]) and their knowledge of squamate muscles.
Remaining muscles innervated by the trigeminal nerve include the m Constrictor Internus Dorsalis and the m. Constrictor Ventralis Trigemini (Haas 1973).
m. Adductor Mandibulae Externus (mAME)
This is an important muscle group used for applying hard bites when the jaws are nearly closed because it is at this point that most of the muscle fibres are orientated perpendicular (orthogonal) to the long axis of the tooth rows (Byerly 1925;
Gorniak et al. 1982;
Jones 2008). The mAME is subdivided into three portions: the superficialis, medialis, and profundus (Lakjer 1926). In general the muscles originate from the lateral surface and dorsal margins of the adductor chamber. The medialis and profundus attach to an aponeurosis (basal aponeurosis or bodenaponeurosis of
Lakjer [1926] and
Anderson [1936]), a tendinous sheet that extends from the dorsal margins of the surangular and coronoid bones, with the medialis inserting on its lateral surface and the profundus inserting on its medial surface.
Some of the origins of the external adductor muscles as figured by
Holliday and Witmer (2007, p. 7, figure 4A) for Sphenodon do not correspond to previous descriptions. Moreover, as labelled, the deep muscles (m. Adductor Mandibulae Externus Profundus [mAMEP]) are reported as originating superficially to more superficial muscles (m. Adductor Mandibulae Externus Medialis [mAMEM]).
m. Adductor Mandibulae Externus Superficialis (mAMES).
Following Haas (1973, p. 196),
Wu (2003) divided the m. Adductor Mandibulae Externus Superficialis into three portions: the m. Levator anguli oris (mLAO), m. Retractor anguli oris (mRAO), and the m. Adductor Mandibulae Externus Superficialis sensu stricto (mAMESss). For lepidosaurs,
Lakjer (1926) refers to the first two subunits (mLAO + mRAO) as Part 1a and the third (mAMESss) subdivision as Part 1b. This terminology was also used by
Rieppel and Gronowski (1981).
The m. Levator Anguli Oris is generally present in squamates but not in crocodiles or birds (Haas, 1973). In Sphenodon,
Haas (1973) described it as "complex" whereas
Wu (2003, p. 590) described it as a strap-shaped muscle that originates from the medial surface of the postorbital near the postorbital-postfrontal joint via "a weak tendon." It has a near vertical path and inserts on the anteromedial surface of the dorsal margin of the rictal plate ("Mundplatte"), part of a tendinous sheet that is visible at the angle of the mouth and attached to the overlying skin (Oelrich 1956;
Wu 2003). This corresponds closely to the descriptions of
Haas (1973, p. 293) and
Gorniak et al. (1982, figures 1 and 2).
Abdala and Moro (2003) code this muscle as being narrow and triangular without an origin on the jugal. Elsewhere the muscle is described as originating from the fascia of the lower temporal fenestra and/or from the lower temporal bar (Poglayen-Neuwall 1953;
Rieppel and Gronowski 1981). Neither
Lakjer (1926) nor
Byerly (1925) described this muscle in Sphenodon.
 The m. Retractor Anguli Oris (mRAO) is thin, triangular, and mainly originates as a sheet-like tendon from the descending process of the squamosal (Haas 1973;
Wu 2003), although some fibres may arise directly from the posteroventral corner of the lower temporal fenestra at the squamosal-quadratojugal joint (Haas 1973;
Wu 2003). The muscle attaches to the dorsalmost margin of the lateral rictal plate after following an anteroventral
path (Haas 1973;
Wu 2003). This muscle does not originate from the lower temporal bar. The m. Retractor Anguli Oris (mRAO) is thin, triangular, and mainly originates as a sheet-like tendon from the descending process of the squamosal (Haas 1973;
Wu 2003), although some fibres may arise directly from the posteroventral corner of the lower temporal fenestra at the squamosal-quadratojugal joint (Haas 1973;
Wu 2003). The muscle attaches to the dorsalmost margin of the lateral rictal plate after following an anteroventral
path (Haas 1973;
Wu 2003). This muscle does not originate from the lower temporal bar.
 The m. Adductor Mandibulae Externus Superficialis (mAMES) sensu stricto is thought to have been small in early amniotes (Heaton 1979;
Rieppel and Gronowski 1981) but it can be very well developed in squamates (Haas 1973;
Gomes 1974;
Rieppel and Gronowski 1981). According to
Wu (2003), the m. Adductor Mandibulae Externus Superficialis (mAMES) sensu stricto may be further divided into an anterolateral component and a smaller posteromedial component.
Gorniak et al. (1982) made a similar, but not necessarily equivalent division (e.g.,
Gorniak et al. 1982,
table 1) The m. Adductor Mandibulae Externus Superficialis (mAMES) sensu stricto is thought to have been small in early amniotes (Heaton 1979;
Rieppel and Gronowski 1981) but it can be very well developed in squamates (Haas 1973;
Gomes 1974;
Rieppel and Gronowski 1981). According to
Wu (2003), the m. Adductor Mandibulae Externus Superficialis (mAMES) sensu stricto may be further divided into an anterolateral component and a smaller posteromedial component.
Gorniak et al. (1982) made a similar, but not necessarily equivalent division (e.g.,
Gorniak et al. 1982,
table 1)
 based on differences in fibre length. based on differences in fibre length.
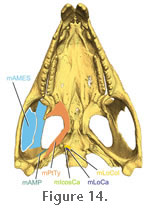
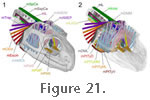
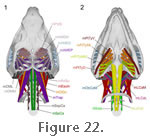
The muscle originates from the fascia of the lower temporal fenestra and also directly from the medial surface of the upper temporal bar (postorbital+squamosal), the neighbouring edges of the squamosal, and the posteromedial surface of the postorbital bar (jugal+postrobital) (Figure 14,
Figure 15,
Figure 16,
Figure 17,
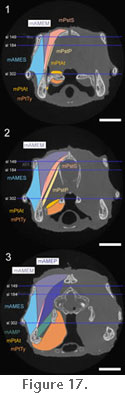 Figure 18,
Figure 19,
Figure 20,
Figure 21,
Figure 22, and
Figure 23) (Edgeworth 1935;
Anderson 1936;
Poglayen-Neuwall 1953;
Haas 1973;
Gorniak et al. 1982;
Wu 2003). The presence of a thin but strong fascia ("Sehnenplatte" of
Poglayen Neuwall 1953) that provides an attachment surface for the mAMES was confirmed by direct examination of specimens (e.g., left side of BMNH1922.6.16.2, right side of FMNH270560 skull, right side of BMNH 1969.2204) (Figure 20). Otherwise the attachment to the medial surface of the postorbital bar is fleshy (Haas 1973). When the jaws are closed the fibres are oriented with a near perpendicular angle to the long axis of the lower jaw (Haas 1973;
Wu 2003). Note that
Haas (1973) and
Lakjer (1926) both considered fibres arising from the medial surface of the upper temporal bar to be part of the mAMEM rather than the mAMES. Figure 18,
Figure 19,
Figure 20,
Figure 21,
Figure 22, and
Figure 23) (Edgeworth 1935;
Anderson 1936;
Poglayen-Neuwall 1953;
Haas 1973;
Gorniak et al. 1982;
Wu 2003). The presence of a thin but strong fascia ("Sehnenplatte" of
Poglayen Neuwall 1953) that provides an attachment surface for the mAMES was confirmed by direct examination of specimens (e.g., left side of BMNH1922.6.16.2, right side of FMNH270560 skull, right side of BMNH 1969.2204) (Figure 20). Otherwise the attachment to the medial surface of the postorbital bar is fleshy (Haas 1973). When the jaws are closed the fibres are oriented with a near perpendicular angle to the long axis of the lower jaw (Haas 1973;
Wu 2003). Note that
Haas (1973) and
Lakjer (1926) both considered fibres arising from the medial surface of the upper temporal bar to be part of the mAMEM rather than the mAMES.
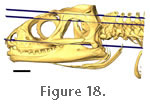 Their
distinction was made because they define the mAMES as only comprising fibres arising from the fascia of the lower temporal fenestra. Their
distinction was made because they define the mAMES as only comprising fibres arising from the fascia of the lower temporal fenestra.
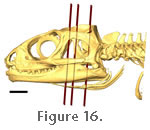
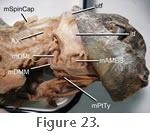
Insertion occurs extensively on the lateral surface of the lower jaw: namely the lateral surface of the dentary, lateral surface of the coronoid bone, and lateral surface of the surangular
(Figure 16,
Figure 17,
Figure 21,
Figure 23,
Figure 24, and
Figure 25;
Anderson 1936;
Rieppel and Gronowski 1981;
Gorniak et al. 1982). According to some authors, a subset of deep fibres may also attach on the lateral surface of the basal aponeurosis (Haas 1973;
Wu 2003). Examination of specimen BMNH 1969.2204 confirms that the mAMES does have a large attachment on the lateral surface of the dentary (Figure
23)
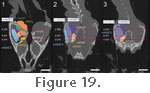 but
also suggests that no part of it originates from the lower temporal bar. When
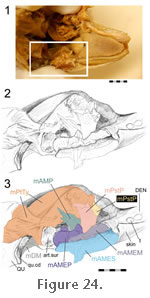
sectioned, the muscle is quite distinct from the other adductor muscles (Figure 24). This muscle does not leave an obvious shelf on the dentary bone as found in many squamates and as suggested for sauropterygians (Figure 5) (Rieppel 2002). but
also suggests that no part of it originates from the lower temporal bar. When
sectioned, the muscle is quite distinct from the other adductor muscles (Figure 24). This muscle does not leave an obvious shelf on the dentary bone as found in many squamates and as suggested for sauropterygians (Figure 5) (Rieppel 2002).
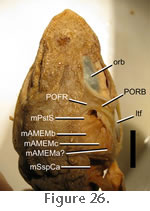
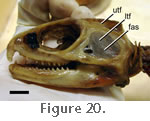 m. Adductor Mandibulae Externus Medialis (mAMEM). Lakjer (1926) referred to the m. Adductor Mandibulae Externus Medialis as Part 2. In general terms the mAMEM originates from the posteromedial boundary of the upper temporal fenestra (posterior to the m. Pseudotemporalis Superficialis) m. Adductor Mandibulae Externus Medialis (mAMEM). Lakjer (1926) referred to the m. Adductor Mandibulae Externus Medialis as Part 2. In general terms the mAMEM originates from the posteromedial boundary of the upper temporal fenestra (posterior to the m. Pseudotemporalis Superficialis)
 (Figure 15,
Figure 16,
Figure 17,
Figure 18,
Figure 19,
Figure 21,
Figure 22,
Figure 26 and
Figure 27).
Abdala and Moro (2003) code the area of origin as simply "parietal and squamosal." Insertion takes place on the lateral surface of the aponeurosis of the lower jaw (Anderson 1936;
Haas 1973;
Gorniak et al. 1982;
Wu 2003) (Figure 16,
Figure 17,
Figure 18,
Figure 19,
Figure 24, and
Figure 25). (Figure 15,
Figure 16,
Figure 17,
Figure 18,
Figure 19,
Figure 21,
Figure 22,
Figure 26 and
Figure 27).
Abdala and Moro (2003) code the area of origin as simply "parietal and squamosal." Insertion takes place on the lateral surface of the aponeurosis of the lower jaw (Anderson 1936;
Haas 1973;
Gorniak et al. 1982;
Wu 2003) (Figure 16,
Figure 17,
Figure 18,
Figure 19,
Figure 24, and
Figure 25).
 Details of the insertions seem to differ between individuals.
Wu (2003) was unable to distinguish any obvious subdivisions but
Gorniak et al. (1982) described three separate portions in the two specimens available to them (Figure 26): Details of the insertions seem to differ between individuals.
Wu (2003) was unable to distinguish any obvious subdivisions but
Gorniak et al. (1982) described three separate portions in the two specimens available to them (Figure 26):
1.
 The ventrolateral head (mAMEMa) – originates from the posterolateral and posteroventral surfaces of the parietal and from the anterior surface of the dorsal process of the squamosal, inserting into the anterior and central sections of the basal aponeurosis. The ventrolateral head (mAMEMa) – originates from the posterolateral and posteroventral surfaces of the parietal and from the anterior surface of the dorsal process of the squamosal, inserting into the anterior and central sections of the basal aponeurosis.
2. The anteromedial head (mAMEMb) – originates on the dorsolateral surface of the parietal crest and inserts into the anterodorsal extension of the basal aponeurosis.
3. The posterior head (mAMEMc) – originates from the posterolateral surface of the parietal and the anterodorsal surface of the squamosal, inserting into anterior and central portions of the basal aponeurosis dorsal to the insertion of the ventrolateral head.
 Haas (1973) listed five subdivisions but the last two subdivisions are probably included within the m. Adductor Mandibulae Externus Superficialis sensu stricto of other authors. Haas (1973) listed five subdivisions but the last two subdivisions are probably included within the m. Adductor Mandibulae Externus Superficialis sensu stricto of other authors.
1. 2a, an anteromedial head (mAMEM2a) – originates from the posteromedial boundary of the upper temporal fenestra (possibly equivalent to mAMEMb of
Gorniak et al. 1982).
2. 2b, a posterior head (mAMEM2b) – originates from the posteromedial boundary of the upper temporal fenestra (possibly equivalent to equivalent to mAMEMc of
Gorniak et al. 1982).
3. 2c, (mAMEM2c) – originates from the squamosal portion of the upper temporal bar (elsewhere treated as part of the mAMES).
4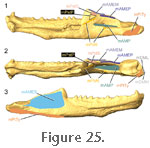 . 2d, (mAMEM2d) –
Haas (1973) simply states that it originates from "the upper temporal arch" but see below (elsewhere this head is treated as part of the mAMES [e.g.,
Poglayen-Neuwall 1953]). . 2d, (mAMEM2d) –
Haas (1973) simply states that it originates from "the upper temporal arch" but see below (elsewhere this head is treated as part of the mAMES [e.g.,
Poglayen-Neuwall 1953]).
5. 2e, (mAMEM2e) –
Haas (1973) simply states that it originates from "the upper temporal arch" but see below (elsewhere this head is treated as part of the mAMES [e.g.,
Poglayen-Neuwall 1953]).
 Of the five divisions outlined,
Haas (1973) only labelled the first four in his figure 10 (p. 300). Furthermore, as noted by
Gorniak et al. (1982, p. 327), the subdivisions that are labelled do not juxtapose to one another as described in the text. Contrary to the figure captions (e.g.,
Haas 1973, p. 300, figure 10a), we suggest that "MAMEB" does not correspond to 2b but instead to 2a, "MAMEC" refers to 2b rather than 2c; and "MAMEA" refers to 2c and not 2a. The label "MAMED" probably does refer to 2d, but in Haas (1973, p. 300, figure 10b) there appears to be a distinct muscle portion between the parts labelled "MAEMD" and "MAEMA". This may represent the muscle described as 2e, or alternatively 2d if "MAEMD" represents 2e. This suggests the mAMEMd and mAMEMe of
Haas (1973) arise from the squamosal at the posterior part of the upper temporal bar. Of the five divisions outlined,
Haas (1973) only labelled the first four in his figure 10 (p. 300). Furthermore, as noted by
Gorniak et al. (1982, p. 327), the subdivisions that are labelled do not juxtapose to one another as described in the text. Contrary to the figure captions (e.g.,
Haas 1973, p. 300, figure 10a), we suggest that "MAMEB" does not correspond to 2b but instead to 2a, "MAMEC" refers to 2b rather than 2c; and "MAMEA" refers to 2c and not 2a. The label "MAMED" probably does refer to 2d, but in Haas (1973, p. 300, figure 10b) there appears to be a distinct muscle portion between the parts labelled "MAEMD" and "MAEMA". This may represent the muscle described as 2e, or alternatively 2d if "MAEMD" represents 2e. This suggests the mAMEMd and mAMEMe of
Haas (1973) arise from the squamosal at the posterior part of the upper temporal bar.
Haas (1973) included divisions 2d and 2e within the m. Adductor Mandibulae Externus Medialis (mAMEM) because they both insert on the lateral surface of the basal aponeurosis, the diagnostic criterion used by
Lakjer (1926). However, other authors consider these portions to be part of the m. Adductor Mandibulae Externus Superficialis (e.g.,
Gorniak et al. 1982;
Wu 2003). We follow the latter interpretation. The upper temporal fenestra is covered by a sheet of soft tissue (right side of BMNH 1969.2204, right side of one FMNH 270560 skull) but the muscles do not originate from it.
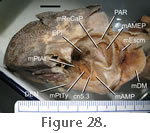
 m. Adductor Mandibulae Externus Profundus (mAMEP).
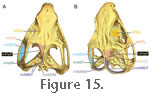
Lakjer (1926) referred to the mAMEP as Part 3. In general, it arises deep to the mAMEM and inserts on the medial surface of the basal aponeurosis posterior to the insertion of the m. Pseudotemporalis Superficialis (mPstS) (Figure 15,
Figure 16,
Figure 17,
Figure 18,
Figure 19,
Figure 21,
Figure 22,
Figure 27, and
Figure 28) (Haas 1973;
Gorniak et al. 1982;
Wu 2003) . The mAMEP is usually described as having two distinct heads of origin (e.g.,
Haas 1973;
Wu 2003;
Gorniak
et al. 1982), but there is disagreement as to the positions of their origins, which could mean that they are not actually equivalent. m. Adductor Mandibulae Externus Profundus (mAMEP).
Lakjer (1926) referred to the mAMEP as Part 3. In general, it arises deep to the mAMEM and inserts on the medial surface of the basal aponeurosis posterior to the insertion of the m. Pseudotemporalis Superficialis (mPstS) (Figure 15,
Figure 16,
Figure 17,
Figure 18,
Figure 19,
Figure 21,
Figure 22,
Figure 27, and
Figure 28) (Haas 1973;
Gorniak et al. 1982;
Wu 2003) . The mAMEP is usually described as having two distinct heads of origin (e.g.,
Haas 1973;
Wu 2003;
Gorniak
et al. 1982), but there is disagreement as to the positions of their origins, which could mean that they are not actually equivalent.
 According to
Haas (1973), the two heads include a posteroventral head (3a) originating from the posterodorsal process of the prootic, and a dorsal head (3b) originating from the posterolateral surface of the upper temporal fenestra (anterior surface of the parietal squamosal process and part of the posteromedial surface of the squamosal). By contrast, neither of the portions described by
Wu (2003) include the prootic as a site of origin. Instead he lists a main portion with fibres that originate from the posterior wall of the upper temporal fenestra and a posteroventral head that originates from the anterolateral surface of the medial process of the squamosal and the dorsolateral surface of the quadrate, posterolateral to the origin of the m. Adductor Mandibulae Posterior. The situation according to
Gorniak et al. (1982) is different again, with both portions, "lateral" and "medial", described as arising from the prootic, more specifically from its posterolateral and anterolateral surfaces respectively, although the lateral head is figured as if arising from the squamosal (Gorniak et al. 1982, p. 328, figure 2). According to
Haas (1973), the two heads include a posteroventral head (3a) originating from the posterodorsal process of the prootic, and a dorsal head (3b) originating from the posterolateral surface of the upper temporal fenestra (anterior surface of the parietal squamosal process and part of the posteromedial surface of the squamosal). By contrast, neither of the portions described by
Wu (2003) include the prootic as a site of origin. Instead he lists a main portion with fibres that originate from the posterior wall of the upper temporal fenestra and a posteroventral head that originates from the anterolateral surface of the medial process of the squamosal and the dorsolateral surface of the quadrate, posterolateral to the origin of the m. Adductor Mandibulae Posterior. The situation according to
Gorniak et al. (1982) is different again, with both portions, "lateral" and "medial", described as arising from the prootic, more specifically from its posterolateral and anterolateral surfaces respectively, although the lateral head is figured as if arising from the squamosal (Gorniak et al. 1982, p. 328, figure 2).
Direct examination of BMNH.1969.2204 shows that part of the m. Adductor Mandibulae Externus Profundus attaches to the posteroventral edge of the posterior temporal bar (parietal and squamosal) and loops anteriorly into the adductor chamber (Figure 28). This perhaps corresponds to the lateral head of
Gorniak et al. (1982) and part of the dorsal (3b) head of
Haas (1973). The majority of the muscle attaches to the fascia of the posterior temporal bar. There are also fibres visible near the top of the quadrate, which is consistent with the posteroventral part of the m. Adductor Mandibulae Externus Profundus as observed by
Wu (2003, p. 591). However, these fibres may instead correspond to the m. Protractor Pterygoidei (Ostrom 1962;
Haas 1973).
It is difficult to reconcile the differences between descriptions of the m. Adductor Mandibulae Externus Profundus but there is a general consensus that the muscle originates in the posterodorsal corner of the adductor chamber and attaches to the medial surface of the basal aponeurosis (Poglayen-Neuwall 1953;
Haas 1973;
Gorniak et al. 1982;
Wu 2003). Discrepancies found in descriptions and observations may stem from intraspecific variation.
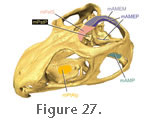
m. Adductor Mandibulae Internus (mAMI)
The m. Adductor Mandibulae Internus comprises the m. Pseudotemporalis and the m. Pterygoideus. These muscles lie between the ophthalmic nerve (cn5.1) and the maxillary nerve (cn5.2), anteromedial to the m. Adductor Mandibulae Externus.
Pseudotemporalis (mPst). The m. Pseudotemporalis seems to correspond, at least in part, to the "pterygoideus externus" of
Byerly (1925) who considered this muscle important for closing the mouth, but also for protraction of the lower jaw.
Gorniak et al. (1982) demonstrated that the m. Pterygoideus Typicus is more important for the latter role. The m. Pseudotemporalis is usually divided into superficial and deep parts (e.g.,
Anderson 1936;
Gorniak et al. 1982;
Barghusen 1973;
Holliday and Witmer 2007), although
Haas (1973, p. 301) cautions that they are not "sharply separated" from each other.
The m. Pseudotemporalis Superficialis (mPstS) originates in the anterior third of the upper temporal fenestra (Figure 15,
Figure 16,
Figure 17,
Figure 18,
Figure 19,
Figure 21,
Figure 22,
Figure 26, and
Figure 27): namely the anterolateral surface of the parietal and posterior surface of the postfrontal (Anderson 1936;
Haas 1973;
Barghusen 1973; BMNH 1972.1223).
Wu (2003) also reported that some fibres come from the posterodorsal tip of the epipterygoid. Its posterior fibres are partly overlain by the m. Adductor Mandibulae Externus Medialis (Anderson 1936;
Gorniak et al. 1982).
Byerly (1925),
Anderson (1936) and
Gorniak et al. (1982) described the area of origin as extending onto the prootic and the posterolateral edge of the lateral process of the postorbital, but this has not been confirmed or accepted by subsequent workers such as
Wu (2003).
The muscle inserts into the medial surface of the basal aponeurosis with some fibres converging on a tendinous extension in a bipennate manner (Anderson 1936;
Haas 1973).
Wu (2003) also reported that some fibres insert directly into the posteromedial edge of the coronoid process (Figure 24 and
Figure 25.1). This may correspond to the point of insertion listed by
Byerly (1925) for the "pterygoideus externus."
Gorniak et al. (1982) described the insertion in the most detail and referred to three sets of fibres:
1. Anterior fibres. These insert via a tendinous extension to the medial surface of the anterior part of the basal aponeurosis.
2. Central fibres. These insert on the anterior section of the tendinous extension, ventral and posterior to the anterior fibres.
3. Posterior fibres. These attach directly into the medial surface of the anterior and central portions of the basal aponeurosis.
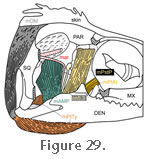 The m. Pseudotemporalis profundus (mPstP) is described by
Haas (1973) as "entirely fleshy" and by
Wu (2003) as being thin and rectangular (Figure 29). It is closely associated with the m. Adductor Mandibulae Posterior (mAMP) (Lakjer 1926;
Haas 1973). Both
Haas (1973) and
Gorniak et al. (1982) described two parts for this muscle, with one effectively overlying the other: The m. Pseudotemporalis profundus (mPstP) is described by
Haas (1973) as "entirely fleshy" and by
Wu (2003) as being thin and rectangular (Figure 29). It is closely associated with the m. Adductor Mandibulae Posterior (mAMP) (Lakjer 1926;
Haas 1973). Both
Haas (1973) and
Gorniak et al. (1982) described two parts for this muscle, with one effectively overlying the other:
1. The external layer (long lateral part of
Gorniak et al.
1982) originates from the anterolateral edge of the parietal and epipterygoid, and also separately from the posterodorsal expansion of the epipterygoid (Haas 1973;
Gorniak et al.
1982).
2. The internal layer (short medial part of
Gorniak et al. 1982) originates from the membranous wall of the braincase (Haas 1973).
Gorniak et al. (1982) also described some fibres originating from the anterior part of the epipterygoid.
Wu (2003) agreed with previous descriptions but did not subdivide the muscle and reported no contribution from the parietal.
There is general agreement that insertion takes place on the medial surface of the lower jaw on or below the coronoid bone (Figure 24 and
Figure 25) (Anderson 1936;
Haas 1973;
Gorniak et al. 1982;
Wu 2003).
Haas (1973) described a long area of attachment on the lingual surface of the lower jaw from the anterior end of the coronoid process, along the prearticular edge of the adductor fossa, to the level of the mandibular foramen. This corresponds to the description of
Gorniak et al. (1982, p. 330 and figure 2).
Wu (2003, p. 592) again records insertion on the medial surface of the coronoid but also to "the anterior part the medial surface of the surangular," presumably above the adductor fossa.
m. Pterygoideus (mPt). The m. Pterygoideus is composed of two parts; a large fleshy m. Pterygoideus Typicus and a smaller anteriorly placed m. Pterygoideus Atypicus (Anderson 1936;
Barghusen 1973;
Haas 1973;
Gorniak et al. 1982;
Wu 2003). The latter is unknown in any extant squamates (Haas 1973;
Gomes 1974). In general this muscle group is most important when the gape is large (Byerly 1925;
Olson 1961;
Jones 2008), but it also plays an important role during protraction of the lower jaw (Gorniak et al. 1982).
The m. Pterygoideus Typicus (mPtTy) is large, complex, and conspicuous, bulging ventrally below the posterior end of the lower jaw (Figure 14,
Figure 16,
Figure 17,
Figure 18,
Figure 19,
Figure 22.2, and
Figure 23;
Byerly 1925;
Haas 1973;
Gorniak et al. 1982;
Wu 2003; FMNH 270560, BMNH 1969.2204). According to
Gorniak et al. (1982) it is the heaviest muscle in the skull (Table 2). Elsewhere, this muscle is referred to as the "pterygoideus ventralis" (e.g.,
Holliday and Witmer 2007), "pterygoideus posterior" (e.g.,
Schumacher 1973b;
Busbey 1989), or the "pterygoideus internus" (Byerly 1925).
Gorniak et al. (1982) provided the most detailed account of this muscle in Sphenodon and considered it in three parts:
1.
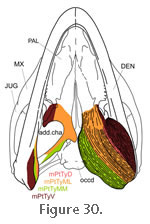 A dorsal (or deep) (mPtTyD) part that originates from the medial surface of the pterygoid along its anterodorsal margin (Figure 30,
Figure 31, and
Figure 32). It inserts into the dorsomedial surface of the posterior end of the lower jaw (on the dentary) by way of fine tendons and fleshy attachment (Figure 22,
Figure 25.1 and
Figure 30). A dorsal (or deep) (mPtTyD) part that originates from the medial surface of the pterygoid along its anterodorsal margin (Figure 30,
Figure 31, and
Figure 32). It inserts into the dorsomedial surface of the posterior end of the lower jaw (on the dentary) by way of fine tendons and fleshy attachment (Figure 22,
Figure 25.1 and
Figure 30).
2.
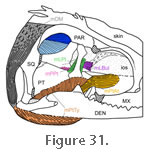 A middle part (the largest part) (mPtTy: mPtTyML and mPtTyML) that originates on the ventral and medial surfaces of the ectopterygoid-pterygoid process, the medial margin and posteroventral half of the pterygoid, and the anteromedial
process of the quadrate (Figure 30,
Figure 31, and
Figure 32). The anterior limit of origin is marked by the "medial pterygoid crest," which is visible in a ventral view of a dried skull (Barghusen 1973;
Gorniak et al. 1982) (Figure 3.1,
Figure 14, and
Figure 22). It inserts on the posteromedial surface of the lower jaw on the dentary, just anterior to the insertion of the dorsal portion (of the mPtTyp)
(Figure 22,
Figure 25 and
Figure 30). A middle part (the largest part) (mPtTy: mPtTyML and mPtTyML) that originates on the ventral and medial surfaces of the ectopterygoid-pterygoid process, the medial margin and posteroventral half of the pterygoid, and the anteromedial
process of the quadrate (Figure 30,
Figure 31, and
Figure 32). The anterior limit of origin is marked by the "medial pterygoid crest," which is visible in a ventral view of a dried skull (Barghusen 1973;
Gorniak et al. 1982) (Figure 3.1,
Figure 14, and
Figure 22). It inserts on the posteromedial surface of the lower jaw on the dentary, just anterior to the insertion of the dorsal portion (of the mPtTyp)
(Figure 22,
Figure 25 and
Figure 30).
3.
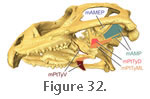 A ventrolateral part (mPtTyV) that originates via a tendon from the lateral and ventral surfaces of the ectopterygoid-pterygoid process (Figure
30 and Figure 32) and inserts on the medial, ventral, and ventrolateral surfaces of the posterior third of the dentary
(Figure 22,
Figure 25.1 and
Figure 30). A ventrolateral part (mPtTyV) that originates via a tendon from the lateral and ventral surfaces of the ectopterygoid-pterygoid process (Figure
30 and Figure 32) and inserts on the medial, ventral, and ventrolateral surfaces of the posterior third of the dentary
(Figure 22,
Figure 25.1 and
Figure 30).
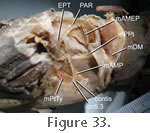 The relationship between the posterolateral surface of the quadrate-pterygoid wing and the mPtTyp (middle part) is unclear. Consistent with
Gorniak et al. (1982), several authors (e.g.,
Barghusen 1973;
Haas 1973;
Wu 2003) describe and figure the m. Pterygoideus Typicus (middle part) as originating on the ventrolateral part of the quadrate. However, in specimen BMNH 1969.2204, some of the muscle previously considered part of the m. Pterygoideus Typicus (e.g.,
Figure 31,
Wu 2003, figure 8) can be seen to lie posterior to the mandibular division of the trigeminal nerve and a boundary of transparent connective tissue (Figure 28 and
Figure 33). The relationship between the posterolateral surface of the quadrate-pterygoid wing and the mPtTyp (middle part) is unclear. Consistent with
Gorniak et al. (1982), several authors (e.g.,
Barghusen 1973;
Haas 1973;
Wu 2003) describe and figure the m. Pterygoideus Typicus (middle part) as originating on the ventrolateral part of the quadrate. However, in specimen BMNH 1969.2204, some of the muscle previously considered part of the m. Pterygoideus Typicus (e.g.,
Figure 31,
Wu 2003, figure 8) can be seen to lie posterior to the mandibular division of the trigeminal nerve and a boundary of transparent connective tissue (Figure 28 and
Figure 33).
 Therefore, this portion of muscle strictly represents a portion of m. Adductor Mandibular Posterior (Figure 32). It also appears to insert into the mandibular fossa rather than more ventrally. Therefore, this portion of muscle strictly represents a portion of m. Adductor Mandibular Posterior (Figure 32). It also appears to insert into the mandibular fossa rather than more ventrally.
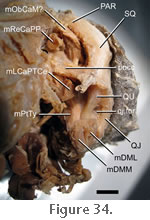
The relationship between the posteromedial surface of the quadrate-pterygoid wing and the middle part of the m. Pterygoideus Typicus is also unclear. Examination of BMNH 1969.2204 suggests that only the ventral edge provides a site of origin (Figure 34), whereas one of the skulls registered as FMNH 270560 and CT data of specimen YPM 9194 suggest that fibres originate from the entire anteroventral corner (Figure 17.3 and
Figure 19.1).
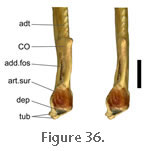
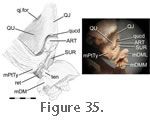 Specimens BMNH 1969.2204 and BMNH 1972.1223 show an additional, previously unreported, insertion for posterior fibres of the m. Pterygoideus Typicus onto the dorsal surface of the lower jaw in a depression behind the articular surface and in front of the m. Depressor Mandibulae (Figure 35). Near this location in specimen PCDG2 there is a tubercle that might be associated with the attachment of muscle fibres (Figure 36). This aspect of the m. Pterygoideus Typicus seems previously to have been overlooked and would probably add stability (via an opposing force) during protrusions of the lower jaw. When activated the majority of the m. Pterygoideus Typicus will aid in closing the jaw, but the action of these posteriormost fibres will provide an opposite, opening force. Specimens BMNH 1969.2204 and BMNH 1972.1223 show an additional, previously unreported, insertion for posterior fibres of the m. Pterygoideus Typicus onto the dorsal surface of the lower jaw in a depression behind the articular surface and in front of the m. Depressor Mandibulae (Figure 35). Near this location in specimen PCDG2 there is a tubercle that might be associated with the attachment of muscle fibres (Figure 36). This aspect of the m. Pterygoideus Typicus seems previously to have been overlooked and would probably add stability (via an opposing force) during protrusions of the lower jaw. When activated the majority of the m. Pterygoideus Typicus will aid in closing the jaw, but the action of these posteriormost fibres will provide an opposite, opening force.
 The m. Pterygoideus Atypicus (mPtAt) is probably equivalent to the pterygoideus anterior, dorsalis, or lateralis in archosaurs (Witmer 1987;
Holliday and Witmer 2007). It is not found in squamates (Haas 1973;
Wu 2003), and in Sphenodon it is quite small compared to the m. Pterygoideus Typicus, being less than 10% the weight (e.g.,
Gorniak et al. 1982;
Wu 2003). Correspondingly it seems to have been overlooked by some workers (e.g.,
Byerly 1925). Nevertheless, the path of this muscle is very different from any other muscle in the skull and probably plays in important role in jaw movement (Gorniak et al. 1982). The m. Pterygoideus Atypicus (mPtAt) is probably equivalent to the pterygoideus anterior, dorsalis, or lateralis in archosaurs (Witmer 1987;
Holliday and Witmer 2007). It is not found in squamates (Haas 1973;
Wu 2003), and in Sphenodon it is quite small compared to the m. Pterygoideus Typicus, being less than 10% the weight (e.g.,
Gorniak et al. 1982;
Wu 2003). Correspondingly it seems to have been overlooked by some workers (e.g.,
Byerly 1925). Nevertheless, the path of this muscle is very different from any other muscle in the skull and probably plays in important role in jaw movement (Gorniak et al. 1982).
The m. Pterygoideus Atypicus originates via a tendon from the dorsal surface of the palate (below the eye) close to, or on, the palatine-pterygoid joint (Haas 1973;
Gorniak et al. 1982;
Wu 2003) (Figure 16,
Figure 17,
Figure 18,
Figure 19,
Figure 21,
Figure 27,
Figure 31, and
Figure 37).
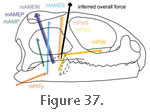 The area of origin may also include the base of the interorbital septum (Haas 1973;
Wu 2003). Posteriorly it extends and loops over the dorsal surface of the pterygoid flange (Figure 21,
Figure 31) (e.g. BMNH 1969.2204;
Haas 1973;
Gorniak et al. 1982;
Wu 2003) as it does in both crocodiles and turtles (Schumacher 1973b;
Holliday and Witmer 2007). The area of origin may also include the base of the interorbital septum (Haas 1973;
Wu 2003). Posteriorly it extends and loops over the dorsal surface of the pterygoid flange (Figure 21,
Figure 31) (e.g. BMNH 1969.2204;
Haas 1973;
Gorniak et al. 1982;
Wu 2003) as it does in both crocodiles and turtles (Schumacher 1973b;
Holliday and Witmer 2007).
The muscle attaches to the lower jaw on or near the coronoid bone, but the exact location appears subject to individual variation.
Haas (1973) observed it to insert below the coronoid,
Poglayen-Neuwall (1953) described it as inserting into the anterior edge of the coronoid, and
Gorniak et al. (1982) described it attaching ventral and posterior to the coronoid (Figure 25.1).
Wu (2003) reported two distinct tendinous insertions of this muscle, and in his account the main part (~80%) of the muscle mass was said to insert onto the posteroventral margin of the coronoid (just above and in front of the adductor fossa), whereas the remaining 20% inserts onto the lower margin of the adductor fossa (Wu 2003, figure17A) (Figure 25.1).
m. Adductor Mandibulae Posterior (mAMP)
This muscle originates from the quadrate and inserts within the adductor fossa of the lower jaw (Haas 1973;
Wu 2003;
Holliday and Witmer 2007). When the jaws are closed the fibres are orientated so that they are nearly vertical (Haas 1973;
Wu 2003).
This muscle was not specifically described by
Gorniak et al. (1982), but
Wu (2003, p. 595) suggested it was figured as part of the m. Adductor Mandibulae Externus Profundus (Gorniak et al.1982, figure 1) because a portion is clearly visible posterior to the course of the mandibular division of the trigeminal nerve (cn5.3). Perhaps significantly, both
Lakjer (1926) and
Haas (1973) noted that the m. Adductor Mandibulae Posterior (mAMP) and m. Adductor Mandibulae Profundus (mAMEP) can appear to be poorly separated. However, in the same figure by
Gorniak et al. (1982) this portion of muscle is drawn as if it attaches to the crest of the lower jaw rather than the adductor fossa, as would be expected according to
Haas (1973) and
Wu (2003). As
Gorniak et al. (1982, figure 1) reported no muscle originating from the quadrate, as would be expected for the mAMP, the muscle may have been absent in the specimens examined or was overlooked entirely.
Abdala and Moro (2003) coded the mAMP as present in Sphenodon.
Barghusen (1973) figured a large area of origin for the mAMP on most of the lateral surface of the pterygoid process of the quadrate (Figure
16, Figure 17.3,
Figure 29, and
Figure 32). In specimen BMNH 1969.2204 this region is largely
free of muscle but this could be because some or all of the muscle was removed during previous dissection (e.g.,
Figure 28 and
Figure 33).
A group of muscle fibres runs from the ventral part of the pterygoid process of the quadrate towards the adductor fossa. This has been considered part of the m. Pterygoideus Typicus in previous descriptions (e.g.,
Figure 29,
Figure 31;
Wu 2003, figure 8). Nonetheless, in specimen BMNH 1969.2204 it is clearly situated posterior to the mandibular division of the trigeminal nerve (Figure 28) and is separated from the m. Pterygoideus Typicus by a portion of transparent connective tissue (Figure 33); it therefore probably represents part of the mAMP (Figure 32). A bundle of muscle fibres attached to the adductor fossa in BMNH 1972.1223 seems to be the insertion of the mAMP as described previously (Haas 1973;
Wu 2003;
Holliday and Witmer 2007).
m. Constrictor Internus Dorsalis (mCID)
This muscle group runs between the braincase and the bones that contribute to the palate. It is divided into three parts: the m. Levator Pterygoidei, the m. Protractor Pterygoidei and the m. Levator Bulbi (Figure 31). In general these muscles connect the braincase to the palate (Haas
1973), but do show individual variation (Table 3;
Ostrom 1962;
Haas 1973;
Wu 2003).
Byerly (1925) did not describe these muscles.
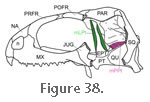 m. Levator Pterygoidei (mLPt). The m. Levator Pterygoidei lies medial to the ophthalmic nerve (cn5.1) (Holliday and Witmer 2007) and has been described as a thin strip of long fibres (Ostrom 1962). It originates on the "ventrolateral surface of the cartilaginous orbitosphenoid medial to the dorsal extremity of the epipterygoid" (Ostrom 1962, p. 733), and inserts on the medial surface of the epipterygoid base and the dorsal and medial surfaces of the pterygoid (Ostrom 1962) (Figure 38).
Poglayen-Neuwall (1953),
Haas (1973) and
Wu (2003) described similar arrangements but with slight differences in fibre length or extent.
Wu (2003) also reported a tendon located posterior and parallel to the m. Levator Pterygoidei (Figure 31).
Lakjer (1926), and correspondingly
Anderson (1936), did not consider this muscle present in Sphenodon but, as
Haas (1973) points out, it does seem to have been figured by
Lakjer (1926) (Table 3). m. Levator Pterygoidei (mLPt). The m. Levator Pterygoidei lies medial to the ophthalmic nerve (cn5.1) (Holliday and Witmer 2007) and has been described as a thin strip of long fibres (Ostrom 1962). It originates on the "ventrolateral surface of the cartilaginous orbitosphenoid medial to the dorsal extremity of the epipterygoid" (Ostrom 1962, p. 733), and inserts on the medial surface of the epipterygoid base and the dorsal and medial surfaces of the pterygoid (Ostrom 1962) (Figure 38).
Poglayen-Neuwall (1953),
Haas (1973) and
Wu (2003) described similar arrangements but with slight differences in fibre length or extent.
Wu (2003) also reported a tendon located posterior and parallel to the m. Levator Pterygoidei (Figure 31).
Lakjer (1926), and correspondingly
Anderson (1936), did not consider this muscle present in Sphenodon but, as
Haas (1973) points out, it does seem to have been figured by
Lakjer (1926) (Table 3).
m. Protractor Pterygoidei (mPPt). The m. Protractor Pterygoidei is located lateral to the ophthalmic nerve (cn5.1) in lepidosaurs (Holliday and Witmer 2007) and is generally described as a broad sheet-like muscle (Anderson 1936;
Ostrom 1962;
Haas 1973). It originates from the lateral surface of the braincase (dorsal surface of the basisphenoid basipterygoid process, the anteroventral part of the prootic, and the ventral part of the orbitosphenoid) and inserts along the dorsal surface of the quadrate-pterygoid
wing (Anderson 1936;
Ostrom 1962;
Haas 1973). The coronal section drawn by
Edgeworth (1907, figure 28) suggests that some anterior fibres of the m. Protractor Pterygoidei ("M. spheno-pterygo-quadratus") may also insert on part of the epipterygoid. Both
Ostrom (1962) and
Haas (1973) found this muscle to be present in one specimen but absent, or almost absent, in another (Table 3).
Wu (2003) also found the muscle in at least one specimen (Figure 31) and although its posterior extent was more limited than described by
Ostrom (1962) (Figure 38),
Wu (2003) did report some anterior fibres originating from a tendon next to the m. Levator Pterygoidei. Vestiges of the m. Protractor Pterygoidei may be visible along the dorsal edge of the quadrate in specimen BMNH 1969.2204 (Figure 33).
The m. Protractor Pterygoidei is associated with kinesis in squamates (e.g.,
Haas 1973). As a result, some authors considered that the presence of this muscle in Sphenodon might indicate the potential for some form of kinetic movement (Lakjer 1926;
Anderson 1936). However, kinesis has never been recorded in Sphenodon, and its sutures do not appear to allow substantial movement between the skull bones (e.g.,
Haas 1973;
Jones 2006a,
2007).
Edgeworth (1935) described this muscle as being present in the embryo and usually retained in the adult leading to suggestions that variation in its development might be related to diet (Ostrom 1962) or ontogeny (Haas 1973). More recent work suggests this muscle is invariably present in adult Sphenodon but can easily be overlooked (Peter Johnson pers. comm. 2009).
m. Levator Bulbi (mLBul). The m. Levator Bulbi is the most developed of the three Constrictor Internus Dorsalis muscles (Haas
1973) and is supposedly the least variable (Wu 2003). The m. Levator Bulbi was not described directly by
Ostrom (1962),
Gorniak et al. (1982), or
Wu (2003).
Haas (1973) divided it into a dorsal and smaller ventral portion.
The m. Levator Bulbi Ventralis (mLBulV) was found in only one of the two
specimens examined by
Haas (1973). There it originated from the anterior edge of
the membranous wall of the braincase and inserted on the palatal membrane in front of, and distinct from, the m. Levator Pterygoidei (Figure 31).
The m. Levator Bulbi Dorsalis (mLBulD) has a tendinous origin from the anteromedial border of the epipterygoid anterior to the m. Levator Bulbi Ventralis, not from the membranous wall of the braincase (contra
Lakjer 1926;
Poglayen-Neuwall 1953). From there it loops anteroventrally and anterodorsally before inserting on the lower eyelid (Haas
1973). In some individuals its midpoint (ventralpoint) may consist of an internal tendon ("Zwischensehne"
Poglayen-Neuwall 1953;
Haas 1973, figure 3). An anterior portion may also arise from the pterygoid or palatine joining the main body of the muscle at the point of the tendon (Haas 1973).
m. Constrictor Ventralis Trigemini (mCVT)
This muscle group was used by
Haas (1973) to accommodate the anterior part of the m. Intermandibularis since its lateral edge is innervated by the trigeminal nerve (cn5.3) via the mylohyoid nerve (e.g.,
Byerly 1925;
Poglayen-Neuwall 1953;
Rieppel 1978). Problematically the posterior and medial part of the muscle is innervated by the facial nerve (cn7) but unlike squamates, it is not possible to recognize an anterior and posterior division of the muscle by gross anatomy (Rieppel 1978).
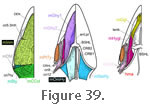 m. Intermandibularis (mInm). The m. Intermandibularis is comprised of transverse fibres that link the anterior three fifths of the lower jaws to one another and wrap around the anterior part of the m. Pterygoid Typicus (Haas 1973;
Rieppel 1978). According to
Rieppel (1978, p. 431) this muscle "grades" into the m. Constrictor Colli posteriorly
(Figure 39). Rieppel's opinion corresponds with the description of
Ruge (1896) but not
Versluys (1898),
von Wettstein (1931,
1932),
Poglayen-Neuwall (1953), or
Haas (1973). This discord may be another example of intraspecific variation or personal perception. The two muscles are certainly separate in prehatchlings at stage P (Edgeworth 1935, figure 459). m. Intermandibularis (mInm). The m. Intermandibularis is comprised of transverse fibres that link the anterior three fifths of the lower jaws to one another and wrap around the anterior part of the m. Pterygoid Typicus (Haas 1973;
Rieppel 1978). According to
Rieppel (1978, p. 431) this muscle "grades" into the m. Constrictor Colli posteriorly
(Figure 39). Rieppel's opinion corresponds with the description of
Ruge (1896) but not
Versluys (1898),
von Wettstein (1931,
1932),
Poglayen-Neuwall (1953), or
Haas (1973). This discord may be another example of intraspecific variation or personal perception. The two muscles are certainly separate in prehatchlings at stage P (Edgeworth 1935, figure 459).
In Sphenodon this muscle may assist in guiding the lower jaws during prooral movement. It is clear from the structure of the jaw joint that in order for the lower jaws to move forwards and backwards, the angle between their long axes must change at the symphysis. Although
Gorniak et al. (1982) did not observe such relative movements, a ligamentous symphysial component may allow it to occur (Günther 1867, p. 600;
Robinson 1976, p. 54).
|