| |
SUMMARY OF RESULTS
General Observations
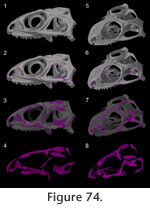 The above description illustrates that a wide variety of joint types can be found in the skull of Sphenodon. For the most part, they can be grouped into the three main categories of abutments, overlaps and interdigitations, but the number and distribution of these joint types across the skull show distinct characteristics. The medial edges of midline bones tend to be dorsoventrally expanded, which increases the surface area of contact (e.g., vomers, parietals). These midline joints are mainly abutments (e.g., interparietal), although some show limited interdigitation (e.g., vomers, pterygoids). In contrast, joints along the sides of the skull involve extensive overlaps (e.g., maxilla-jugal, postorbital-squamosal) (Appendix 2;
Figure 74,
Figure 75,
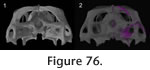
Figure 76,
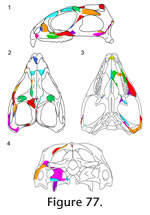
Figure 77). Some of these overlap joints are relatively simple contacts (e.g., internasal) whereas others have more complex interlocking arrangements (e.g., frontal-parietal, palatine-prefrontal); joints where a tongue-shaped process sits in a basin-like depression are common (e.g., jugal-maxilla, postfrontal-postorbital, nasal-frontal, postorbital-squamosal). Less frequent in Sphenodon, in contrast to most early tetrapods (e.g.,
Kathe 1995), are joints in which bones meet along the same plane. Nevertheless, some planar joints are found in the skull roof (e.g., nasal-frontal, frontal-parietal), palate (palatine-pterygoid, palatine-vomer) or lateral surfaces (e.g., palatine-maxilla, ectopterygoid-jugal). Interdigitated joints are rare in Sphenodon and essentially limited to the interpterygoid and the prefrontal-palatine joints. Extensive Type-C interdigitations like those found in mammals, chelonians and crocodiles (e.g.,
Iordansky 1973;
Gaffney 1979;
Jaslow 1990) are absent. The above description illustrates that a wide variety of joint types can be found in the skull of Sphenodon. For the most part, they can be grouped into the three main categories of abutments, overlaps and interdigitations, but the number and distribution of these joint types across the skull show distinct characteristics. The medial edges of midline bones tend to be dorsoventrally expanded, which increases the surface area of contact (e.g., vomers, parietals). These midline joints are mainly abutments (e.g., interparietal), although some show limited interdigitation (e.g., vomers, pterygoids). In contrast, joints along the sides of the skull involve extensive overlaps (e.g., maxilla-jugal, postorbital-squamosal) (Appendix 2;
Figure 74,
Figure 75,
Figure 76,
Figure 77). Some of these overlap joints are relatively simple contacts (e.g., internasal) whereas others have more complex interlocking arrangements (e.g., frontal-parietal, palatine-prefrontal); joints where a tongue-shaped process sits in a basin-like depression are common (e.g., jugal-maxilla, postfrontal-postorbital, nasal-frontal, postorbital-squamosal). Less frequent in Sphenodon, in contrast to most early tetrapods (e.g.,
Kathe 1995), are joints in which bones meet along the same plane. Nevertheless, some planar joints are found in the skull roof (e.g., nasal-frontal, frontal-parietal), palate (palatine-pterygoid, palatine-vomer) or lateral surfaces (e.g., palatine-maxilla, ectopterygoid-jugal). Interdigitated joints are rare in Sphenodon and essentially limited to the interpterygoid and the prefrontal-palatine joints. Extensive Type-C interdigitations like those found in mammals, chelonians and crocodiles (e.g.,
Iordansky 1973;
Gaffney 1979;
Jaslow 1990) are absent.
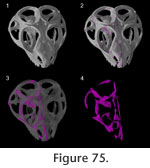 Two different joints may occur between the same pair of bones (e.g., palatine and maxilla, prefrontal and maxilla) and, as in other tetrapods (e.g.,
Clack 2002), joint morphology can change along the suture (e.g., the palatine-pterygoid suture). At one point in a joint 'bone X' may overlap 'bone Y' but at another part of the suture the opposite occurs (e.g., nasal-prefrontal, prefrontal-frontal, intervomerine, postfrontal-frontal, frontal-parietal). As in other taxa (e.g.,
Moss 1957;
Clack, 2002;
Sun et al. 2004), some joints bear a complex external seam but internally the contacting surfaces are relatively smooth (e.g., ectopterygoid-jugal joint). Two different joints may occur between the same pair of bones (e.g., palatine and maxilla, prefrontal and maxilla) and, as in other tetrapods (e.g.,
Clack 2002), joint morphology can change along the suture (e.g., the palatine-pterygoid suture). At one point in a joint 'bone X' may overlap 'bone Y' but at another part of the suture the opposite occurs (e.g., nasal-prefrontal, prefrontal-frontal, intervomerine, postfrontal-frontal, frontal-parietal). As in other taxa (e.g.,
Moss 1957;
Clack, 2002;
Sun et al. 2004), some joints bear a complex external seam but internally the contacting surfaces are relatively smooth (e.g., ectopterygoid-jugal joint).
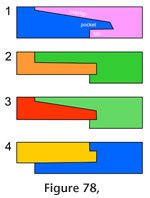
Many joints, particularly along the dorsal and ventral margins of the skull, involve three or, occasionally, four bones (Figure 77, e.g., jugal-maxilla-ectopterygoid, frontal-postfrontal-parietal, jugal-squamosal-quadratojugal). This arrangement can result in strong triple overlaps (e.g., the maxilla-prefrontal-nasal, vomer-palatine-pterygoid), although similar complex overlaps can occur between two bones where one possesses a pocket or tab into which the other slots (Figure 78) (e.g., premaxilla-nasal, palatine-vomer, nasal-maxilla, parietal-postfrontal, and to some extent, the palatine-prefrontal and squamosal-postorbital).
 Following disarticulation of the skull, relatively few bones can be rearticulated so that they remain locked together in most orientations (e.g., the postfrontal and postorbital or the frontals and parietals). Most cranial joints require soft tissue and/or the presence of other bones
to maintain the articulation (e.g., palatine-maxilla or premaxilla-palatine). Indeed, the surfaces of a few bones bear no trace of facets and are maintained entirely by soft tissue (e.g., the premaxilla-vomerine joint). This characteristic reflects the amount of soft tissue in many of the cranial joints of this genus and explains the rarity of articulated Holocene specimens of Sphenodon (Gill, personal commun., 2006). Following disarticulation of the skull, relatively few bones can be rearticulated so that they remain locked together in most orientations (e.g., the postfrontal and postorbital or the frontals and parietals). Most cranial joints require soft tissue and/or the presence of other bones
to maintain the articulation (e.g., palatine-maxilla or premaxilla-palatine). Indeed, the surfaces of a few bones bear no trace of facets and are maintained entirely by soft tissue (e.g., the premaxilla-vomerine joint). This characteristic reflects the amount of soft tissue in many of the cranial joints of this genus and explains the rarity of articulated Holocene specimens of Sphenodon (Gill, personal commun., 2006).
 Facet surfaces, where present, are generally smooth but can be ridged (e.g., nasal-frontal), pitted (e.g., pterygoid-ectopterygoid) or striated (e.g., maxilla-jugal) to maximise the attachment area for collagen fibres. Within a single joint there may be variation in the location of texture (e.g., jugal-postorbital, maxilla-jugal). Usually the texture is reflected on opposing facet surfaces but not always (e.g., prefrontal-maxilla). Facet surfaces, where present, are generally smooth but can be ridged (e.g., nasal-frontal), pitted (e.g., pterygoid-ectopterygoid) or striated (e.g., maxilla-jugal) to maximise the attachment area for collagen fibres. Within a single joint there may be variation in the location of texture (e.g., jugal-postorbital, maxilla-jugal). Usually the texture is reflected on opposing facet surfaces but not always (e.g., prefrontal-maxilla).
Variation within Sphenodon
In Sphenodon, sutures in larger skulls usually show a greater degree of overlap and facet texture than those in small skulls. Individual joints generally have the same gross morphology between skulls of similar size but there is variation in the edges of processes, position of seams, facet texture and other minor details. Given the limited sample of juvenile Sphenodon skulls, it is still difficult to disentangle ontogenetic variation from individual variation, but some differences are obvious.
 Adult (e.g., DGPC1, BMNH.K) and juvenile skulls (e.g., UCLGMX 1176) differ from large hatchling skulls (e.g.,
Howes and Swinnerton 1901;
Rieppel 1992) in the following ways: the frontoparietal and interparietal fontanelles have closed; the posterior process of the jugal is more extensively sutured to the quadrate and squamosal and the overlap between the squamosal and postorbital is more extensive. Adult skulls (e.g., BMNH.K, DGPC1) differ from juvenile skulls (e.g., UCLGMX 1176) in the following ways: the interpremaxillary seam is less sigmoid; the ectopterygoid-jugal joint tends to be more interdigitated; the prefrontal-palatine seam tends to be more interdigitated; the jugal dorsal process has a squared off dorsal tip and the parietal crest is dorsally expanded. Between adult skulls (e.g., BMNH.K and DGPC1) the following variations occur: the presence or absence of the postfrontal cleft; the degree to which the vomer underlaps the palatine, the palatine overlaps the pterygoid centrally, the prefrontal overlaps the nasal and the frontal underlaps the nasal; the presence or absence of a nasofrontal fontanelle; the shape of the nasofrontal, postfrontal-frontal and frontal-parietal seams; the level of texture present on the facets of the nasal-frontal joint; the exact arrangement of the jugal-squamosal-quadratojugal joint; and the degree to which the squamosal envelopes the postorbital. Adult (e.g., DGPC1, BMNH.K) and juvenile skulls (e.g., UCLGMX 1176) differ from large hatchling skulls (e.g.,
Howes and Swinnerton 1901;
Rieppel 1992) in the following ways: the frontoparietal and interparietal fontanelles have closed; the posterior process of the jugal is more extensively sutured to the quadrate and squamosal and the overlap between the squamosal and postorbital is more extensive. Adult skulls (e.g., BMNH.K, DGPC1) differ from juvenile skulls (e.g., UCLGMX 1176) in the following ways: the interpremaxillary seam is less sigmoid; the ectopterygoid-jugal joint tends to be more interdigitated; the prefrontal-palatine seam tends to be more interdigitated; the jugal dorsal process has a squared off dorsal tip and the parietal crest is dorsally expanded. Between adult skulls (e.g., BMNH.K and DGPC1) the following variations occur: the presence or absence of the postfrontal cleft; the degree to which the vomer underlaps the palatine, the palatine overlaps the pterygoid centrally, the prefrontal overlaps the nasal and the frontal underlaps the nasal; the presence or absence of a nasofrontal fontanelle; the shape of the nasofrontal, postfrontal-frontal and frontal-parietal seams; the level of texture present on the facets of the nasal-frontal joint; the exact arrangement of the jugal-squamosal-quadratojugal joint; and the degree to which the squamosal envelopes the postorbital.
There are several possible reasons for individual differences in joint morphology between adults of Sphenodon. One is variation in the pattern of mechanical forces transmitted around the skull, which is known to influence joint morphology (e.g.,
Moss 1954,
1961;
Oudhof and Markens 1982;
Nanda and Hickory 1984;
Nash and Kokich 1985;
Mao 2002;
Mao et al. 2003). Variation in the stress regime may be the result of differences in diet (e.g.,
Ostrom 1962;
Dalrymple 1977) so that animals with more abrasive diets experience more stress and therefore possess more complex sutures (Byron et al. 2004). It may also be related to where an individual prefers to bite along its tooth row. Variations in muscle arrangement (e.g.,
Haas 1973;
Gorniak et al. 1981;
Wu 2003;
Jones et al. 2009), potentially linked to differences in genotype or behaviour, may also alter the location and orientation of stresses, as may variations in skull size and proportions linked to ontogeny (Jones 2008).
|