| |
RESULTS
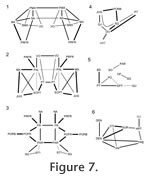 In Sphenodon there are as many as 113 cranial joints. Of these, 106 are paired and 7 are midline (interpremaxillary, internasal, interfrontal, interparietal, intervomer, interpalatine and interpterygoid) (Figure 7,
Table 4). This number does not include the symphysial joint between the lower jaws, joints between the braincase bones, and paired jaw joints between the quadrate and articular. The joints of the skull are divided between seven categories or units, depending on their location within the skull (Figure 7,): rosral joints (20 in all), palatal joints (21 to 25), roofing joints (16 plus), temporal joints (14 plus, 2 mainly fused), metakinetic joints (14), intraoccipital joints and mandibular joints (14 plus 4 fused). The latter two categories will not be discussed in detail here. The joints of most individual bones are restricted to one unit but those of the pterygoid and prefrontal are distributed between three. In Sphenodon there are as many as 113 cranial joints. Of these, 106 are paired and 7 are midline (interpremaxillary, internasal, interfrontal, interparietal, intervomer, interpalatine and interpterygoid) (Figure 7,
Table 4). This number does not include the symphysial joint between the lower jaws, joints between the braincase bones, and paired jaw joints between the quadrate and articular. The joints of the skull are divided between seven categories or units, depending on their location within the skull (Figure 7,): rosral joints (20 in all), palatal joints (21 to 25), roofing joints (16 plus), temporal joints (14 plus, 2 mainly fused), metakinetic joints (14), intraoccipital joints and mandibular joints (14 plus 4 fused). The latter two categories will not be discussed in detail here. The joints of most individual bones are restricted to one unit but those of the pterygoid and prefrontal are distributed between three.
Rostral Joints
 The rostral (or facial) joints include those joints surrounding the anterior part of the skull. The rostral unit is connected to the roofing unit by the nasals and prefrontals and to the palatal unit by the maxillae, prefrontals and vomers. The rostral (or facial) joints include those joints surrounding the anterior part of the skull. The rostral unit is connected to the roofing unit by the nasals and prefrontals and to the palatal unit by the maxillae, prefrontals and vomers.
Interpremaxillary
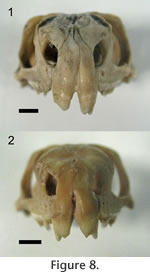
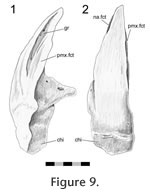
In Sphenodon the premaxillae contact one another along their longest axis. In external view the dorsal portion of the joint has a seam that is sagittally orientated and generally straight, although a slight sigmoid kink is often visible (e.g., LDUCZ x036, LDUCZ x343, LDUCZ x723). This sigmoid kink is particularly large in the juvenile specimen LDUCZ x1176. In the ventral portion of the joint the external seam widens to form an ovoid gap between the anterior edges of the two premaxillae (e.g., DGPC2). In life this is filled by a plug of soft tissue (e.g., LDUCZ x036) (Figure 8). The premaxilla does not have a palatal shelf and therefore the seam is short in ventral view, limited to the alveolar rim. The joint is generally a butt joint (e.g., YPM 11419) but some texture is apparent, consisting of dorsoventrally directed striations. In AIM LH0617 there is a more obvious groove running the length of the facet surface (Figure 9), and CT scans of LDUCZ x036 demonstrate that the kink in the external seam corresponds to an overlap between the two bones. Examination of this joint is not possible in DGPC1.
 Premaxilla-maxilla Premaxilla-maxilla
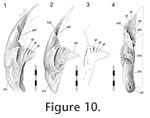
In Sphenodon the anterior end of the maxilla loosely overlaps the lateral process of the premaxilla below the naris, with a 'recessed scarf' joint (Figure 2.10). In dorsal view, the seam runs posteromedially from near the anterolateral corner of the external naris. In lateral view the seam is generally straight, running dorsoventrally from the ventral margin of the naris to the ventral edge of the tooth row (e.g., LDUCZ x036). However, the anterior process of the maxilla is often curved anteroventrally (e.g., DGPC1, LDUCZ x343, LDUCZ x723, YPM 11419), widening the seam between the two bones. Disarticulated specimens demonstrate that there is an initial butting contact anteriorly (a vertical wall at about 90º to the outer cranial surface and about 25% of the mediolateral thickness of the premaxilla). Posterior to this, there is a scarf joint where the angle of overlap approaches 45º. The facet on the premaxilla is generally flat although there is vertical fluting in specimen BMNH.K without corresponding fluting on the maxilla. The posterior process of the premaxilla is relatively long but tapers symmetrically reducing the area of contact with the maxilla (Figure 10). The joint does not seem to be closely apposed, and in life probably involves a substantial amount of soft tissue, particular at its ventral portion. Examination of the joint in CT section of YPM 9194 and LDUCZ x036 confirms this observation.
 Without considering soft tissue, this joint shape appears to restrict the posterior part of the premaxilla from rotating laterally, and the anterior end of the maxilla from rotating medially. In the articulated skull, the paired premaxillae are held between the maxillae, but not firmly. This joint (without soft tissue) would not restrict anterior movement of the premaxilla or posterior movement of the maxilla and neither would it prevent vertical or downward movement of either bone. Because movement of the maxilla is restricted by several other bones (nasal, prefrontal, palatine, jugal, ectopterygoid) freedom at this joint has greater implications for the premaxilla although the maxilla is sometimes found to be displaced in dried specimens (e.g., LDUCZ x343). Without considering soft tissue, this joint shape appears to restrict the posterior part of the premaxilla from rotating laterally, and the anterior end of the maxilla from rotating medially. In the articulated skull, the paired premaxillae are held between the maxillae, but not firmly. This joint (without soft tissue) would not restrict anterior movement of the premaxilla or posterior movement of the maxilla and neither would it prevent vertical or downward movement of either bone. Because movement of the maxilla is restricted by several other bones (nasal, prefrontal, palatine, jugal, ectopterygoid) freedom at this joint has greater implications for the premaxilla although the maxilla is sometimes found to be displaced in dried specimens (e.g., LDUCZ x343).
 Premaxilla-nasal Premaxilla-nasal
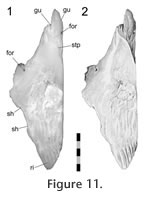
The premaxilla overlaps the anterior process of the nasal in a 'recessed scarf' joint (Figure 2.10) in which the anterior end of the nasal process slots into a 'pocket' in the back of the premaxilla (at least in DGPC1). In dorsal view, the nasal processes of the paired premaxillae appear to be pinched between the nasals with seams that run posteromedially from the dorsal margin of the naris toward the midline. The internal structure of the joint is complex (e.g., LDUCZ x343, YPM 11419, DGPC1, AUP 11883). First, the anterior process of the nasal, which extends beneath the premaxilla, is triangular and directed anterolaterally, so the hidden anterior processes of the nasals do not meet along the midline but diverge (Figure 11,
Figure 12,
Figure 13). Second, the facet on the nasal is sunk or recessed as in a 'recessed scarf' joint.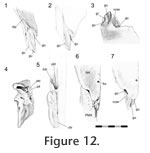 It is deep medially but shallows laterally (Figure 11.2,
Figure 12.1). As a result the paired premaxillae are wedged against each other. Third, the anterior tip of the pointed nasal process fits into a pocket in the posteroventral surface of the premaxilla (Figure 10.4,
Figure 12.4, 12.5). Hence, the nasal has both dorsal and ventral facets for the premaxilla (Figure 12.2, 12.7). The posterior wall of the pocket is fairly low, and the interior contains five pits that are probably related to nutrient supply (Figure 12.4). It is deep medially but shallows laterally (Figure 11.2,
Figure 12.1). As a result the paired premaxillae are wedged against each other. Third, the anterior tip of the pointed nasal process fits into a pocket in the posteroventral surface of the premaxilla (Figure 10.4,
Figure 12.4, 12.5). Hence, the nasal has both dorsal and ventral facets for the premaxilla (Figure 12.2, 12.7). The posterior wall of the pocket is fairly low, and the interior contains five pits that are probably related to nutrient supply (Figure 12.4).
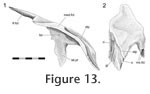 Due to bisection of the skull only the lateral half the premaxilla-nasal joint is known for DGPC1. However, the available portion shows the presence of three gutters on the anterior process of the nasal that run parallel to the long axis of the process itself (anterolaterally). The posterior ends of the lateral and medial gutters are visible on the dorsal surface but excavate the surface laterally and medially, respectively. The central gutter is more visible ventrally and terminates at the tip of the anterior nasal process. On the ventral surface of the process the three gutters are separated by a shallow groove and a concavity. The largest and most lateral of the gutters (Figure 12.1, 12.2) corresponds to a tubercle on the ventral surface of the premaxilla above the pocket (Figure 10.2, 10.4,
Figure 12.5, 12.6). The two smaller gutters on the nasal also interlock with ridges inside the pocket of the premaxilla. The medial parts of the facets, as seen in BMNH.K, bear longitudinal grooves and ridges (Figure 13). In the juvenile Sphenodon, LDUCZ x1176, the nasals appear to abut the edge of the premaxillary pocket but do not enter it. Due to bisection of the skull only the lateral half the premaxilla-nasal joint is known for DGPC1. However, the available portion shows the presence of three gutters on the anterior process of the nasal that run parallel to the long axis of the process itself (anterolaterally). The posterior ends of the lateral and medial gutters are visible on the dorsal surface but excavate the surface laterally and medially, respectively. The central gutter is more visible ventrally and terminates at the tip of the anterior nasal process. On the ventral surface of the process the three gutters are separated by a shallow groove and a concavity. The largest and most lateral of the gutters (Figure 12.1, 12.2) corresponds to a tubercle on the ventral surface of the premaxilla above the pocket (Figure 10.2, 10.4,
Figure 12.5, 12.6). The two smaller gutters on the nasal also interlock with ridges inside the pocket of the premaxilla. The medial parts of the facets, as seen in BMNH.K, bear longitudinal grooves and ridges (Figure 13). In the juvenile Sphenodon, LDUCZ x1176, the nasals appear to abut the edge of the premaxillary pocket but do not enter it.
This joint would prevent the posterior end of the premaxilla from rotating posteroventrally but some anterior rotation would be possible without soft tissue (again as seen in LDUCZ x343). The butting wall of the 'recessed scarf' and edge of the premaxillary pocket would obstruct posterodorsal movement of the premaxilla but would not restrict anteroventral movement. The small grooves and gutters on the facets, and the shape of the premaxillary pocket would inhibit mediolateral movement and increase surface area for soft tissue. The change in depth of the scarf joint would also discourage sideways movements between the nasal and premaxilla. Moreover, in an articulated skull, medial movement of one premaxilla would be prevented by the other premaxilla.
Internasal
 The nasals meet along the midline between the premaxillae and frontals. The external seam is usually straight (LDUCZ x343, LDUCZ x723, NMNZ RE0385) but it can also be curved (LDUCZ x1176), irregular (LDUCZ x146) or slightly sigmoid (LDUCZ x343; BMB 100225). The length of the seam also varies in comparison with the posterior extent of the premaxillae or interorbital width; it may be relatively long (LDUCZ x721, BMB 101668, LDUCZ x343) or short (LDUCZ x723, KCL x12, BMNH 1972.1499) (Figure 14;
Jones and Lappin 2009, figure 4D). The medial edge of the anterior process for the premaxilla, seen only in disarticulated specimens, provides further contact area between the nasals (Figure 13.1). Nevertheless, contact with the premaxillae and frontals occurs across a far greater surface area. In general the internasal is a butt joint although very small shelves of bone may 'invade' the adjoining nasal ventrally. In one specimen (BMNH 1985.1212) the centre of the left nasal exhibits a large pathological hole from which seams extend anteriorly and posteriorly. The nasals meet along the midline between the premaxillae and frontals. The external seam is usually straight (LDUCZ x343, LDUCZ x723, NMNZ RE0385) but it can also be curved (LDUCZ x1176), irregular (LDUCZ x146) or slightly sigmoid (LDUCZ x343; BMB 100225). The length of the seam also varies in comparison with the posterior extent of the premaxillae or interorbital width; it may be relatively long (LDUCZ x721, BMB 101668, LDUCZ x343) or short (LDUCZ x723, KCL x12, BMNH 1972.1499) (Figure 14;
Jones and Lappin 2009, figure 4D). The medial edge of the anterior process for the premaxilla, seen only in disarticulated specimens, provides further contact area between the nasals (Figure 13.1). Nevertheless, contact with the premaxillae and frontals occurs across a far greater surface area. In general the internasal is a butt joint although very small shelves of bone may 'invade' the adjoining nasal ventrally. In one specimen (BMNH 1985.1212) the centre of the left nasal exhibits a large pathological hole from which seams extend anteriorly and posteriorly.
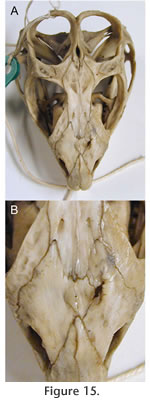 Possibly as a response, the midline internasal seam has partially fused (Figure 15). Alternatively the right nasal may have grown to compensate for the left and a suture subsequently developed within it. In hatchling skulls there is a fontanelle between the nasals and frontals (Howes and Swinnerton 1901;
Rieppel 1992;
Jones and Lappin 2009, figure 4). As previously reported remains of it can be found in a number of adult skulls, and it may be over 1 mm in diameter (e.g., MANCH C120649, AMPC1, UCMZ 2614, KCL x12) (Jones et al. 2009). Possibly as a response, the midline internasal seam has partially fused (Figure 15). Alternatively the right nasal may have grown to compensate for the left and a suture subsequently developed within it. In hatchling skulls there is a fontanelle between the nasals and frontals (Howes and Swinnerton 1901;
Rieppel 1992;
Jones and Lappin 2009, figure 4). As previously reported remains of it can be found in a number of adult skulls, and it may be over 1 mm in diameter (e.g., MANCH C120649, AMPC1, UCMZ 2614, KCL x12) (Jones et al. 2009).
Nasal-prefrontal
In dorsal view the external seam of this joint is sub-parallel to the premaxillary-nasal joint, being anterolaterally directed and generally straight before it disappears under the maxilla (Figure 5.2,
Figure 6,
Figure 14,
Figure 15). In lateral view (with the maxilla removed) the seam continues between the ventrolateral process of the nasal and lateral process of the prefrontal, travelling at first anterolaterally and then ventrally. This ventral part of the seam (hidden in articulated skulls by the facial process of the maxilla) exhibits some intraspecific variation; it may be sigmoid as in DGPC1 (Figure 16,
Figure 17.1) or almost straight as in YPM 11419. The form of the nasal-prefrontal joint changes substantially along its distance, and it seems logical to divide it into two parts; a posterior portion (best seen in dorsal view) and an anterior portion (best seen in lateral view with the maxilla removed).
The posterior portion of the joint consists of a weak slot joint (Figure 16). In DGPC1, the posterior edge of nasal overlaps a short shelf or lappet of bone from the prefrontal. This shelf in turn meets the large anterior process of the underlying frontal. A smaller longitudinal nasal shelf also projects under the prefrontal shelf for a small distance, creating a small narrow slot joint posteriorly. Midway along the posterior portion of the joint in DGPC1 the seam is wide, and contact between the two bones is lost.
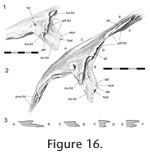 These facts are not evident in LDUCZ x036, LDUCZ x343 or DGPC2 where externally the prefrontal may appear to encroach upon the nasal. In BMNH.K, nasal-prefrontal contact is also retained, and there appears to be a much longer shelf from the nasal underlapping the prefrontal. Medial to the facial process of the maxilla the nasal overlaps a triangular shelf on the prefrontal. This shelf is continuous with the lateral process of the prefrontal that continues ventrally. Overall the posterior section of the joint is not very strong, but would resist some dorsoventral movement between the bones. These facts are not evident in LDUCZ x036, LDUCZ x343 or DGPC2 where externally the prefrontal may appear to encroach upon the nasal. In BMNH.K, nasal-prefrontal contact is also retained, and there appears to be a much longer shelf from the nasal underlapping the prefrontal. Medial to the facial process of the maxilla the nasal overlaps a triangular shelf on the prefrontal. This shelf is continuous with the lateral process of the prefrontal that continues ventrally. Overall the posterior section of the joint is not very strong, but would resist some dorsoventral movement between the bones.
The anterior portion of the nasal-prefrontal joint is associated with the maxilla-nasal joint. In a lateral view (with the maxilla removed), the lateral wing of the prefrontal can be seen to overlap about a third of the lateral wing of the nasal (Figure 16,
Figure 17,
Figure 18). The dorsal portion of this overlap is a 'recessed scarf' joint (Figure
16) but more ventrally the abutting wall on the nasal diminishes so that the contact more closely resembles a lap joint (Figure
16).
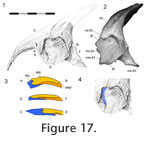 At this same point in DGPC1, two small projections (tabs) from the nasal facet increase the overlap distance. Ventral to this, the edge of the prefrontal bears a medially directed fold on to which an expanded foot-like part of the nasal process sits, effectively overlapping the prefrontal and producing a small anteroposteriorly directed butt joint. The ventral tips of the nasal and prefrontal lateral facets barely touch. At this same point in DGPC1, two small projections (tabs) from the nasal facet increase the overlap distance. Ventral to this, the edge of the prefrontal bears a medially directed fold on to which an expanded foot-like part of the nasal process sits, effectively overlapping the prefrontal and producing a small anteroposteriorly directed butt joint. The ventral tips of the nasal and prefrontal lateral facets barely touch.
The naso-prefrontal joint, although complicated, does not appear strong even when the overlap is substantial.
 The bone is fairly thin and unbuttressed, and some of the detail may be subject to intraspecific variation. Nevertheless the joint would obstruct downward movement of the nasal relative to the prefrontal and lateral rotation of the posterior end of the nasal. Consequently, the joint would also obstruct upward movements of the prefrontal and medial rotation of its posterior end. Anterolateral and posteromedial movements along the joint would also be inhibited (at least in DGPC1). It would not prevent upward or medial movement of the nasal and correspondingly neither would it prevent downward or lateral movement of the prefrontal. The bone is fairly thin and unbuttressed, and some of the detail may be subject to intraspecific variation. Nevertheless the joint would obstruct downward movement of the nasal relative to the prefrontal and lateral rotation of the posterior end of the nasal. Consequently, the joint would also obstruct upward movements of the prefrontal and medial rotation of its posterior end. Anterolateral and posteromedial movements along the joint would also be inhibited (at least in DGPC1). It would not prevent upward or medial movement of the nasal and correspondingly neither would it prevent downward or lateral movement of the prefrontal.
Maxilla-septomaxilla
In Sphenodon the septomaxilla is a small curved bone. Its posterolateral surface rests against the dorsomedial edge of the maxilla at the base of the nares (e.g., DGPC1, LDUCZ x036).
Premaxilla-septomaxilla
A small anterior portion of the septomaxilla rests against the dorsal surface of the premaxillary lateral process. No obvious facet can be found on either bone.
Vomer-septomaxilla
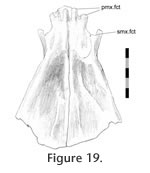 The anterior portion of the vomer has a dorsally expanded lateral edge. The anterior border of this raised edge bears a C-shaped embayment, making a hook-like process that accepts the anteromedial end of the septomaxilla or at least soft tissue associated with it (Figure 19). The anterior portion of the vomer has a dorsally expanded lateral edge. The anterior border of this raised edge bears a C-shaped embayment, making a hook-like process that accepts the anteromedial end of the septomaxilla or at least soft tissue associated with it (Figure 19).
Maxilla-nasal
The external seam of this joint occurs between the lateral part of the nasal and the anterodorsal edge of the facial process of the maxilla. In lateral view the posterior part may initially be directed anteriorly but after a short distance it turns ventrally as the facial process curves toward the base of the naris (e.g.,
Figure 5).
As described above, the long ventrolateral process of the nasal is partly overlain by the prefrontal but the remaining part of the faceted process is overlain by the facial process of the maxilla (Figure 17,
Figure 18). Hence, the variation in the degree and shape of the maxilla-nasal contact is related to that found in the prefrontal-nasal overlap, for example in DGPC1 the maxilla-nasal contact is greatest ventrally whereas in YPM 11419 it is more evenly distributed. Anterodorsally there is also a slot arrangement peripheral to the main overlap (Figure 17.3). Here a short but wide, posteriorlydirected projection from the nasal slots into a dorsoventrally directed groove on the anterior edge of the maxilla facial process. This arrangement results in a triple overlap, from the medial to lateral surface: nasal-maxilla, maxilla-nasal, nasal-maxilla. In a lateral view of DGPC1, with the maxilla removed, the lateral surface of the nasal is slightly recessed compared to that of the prefrontal (Figure 17.3), creating a dorsoventrally orientated trough. Correspondingly the anterior portion of the facial process has a medial bulge. The maxillary facet of the nasal bears parallel striations directed posteroventrally but similar striations are not obvious on the medial facet of the maxilla. This joint restricts anterior movement of the maxilla and to some extent would restrict lateral or medial rotation of the tooth row. Posteroventral movement of the maxilla is not prevented by this joint alone.
Maxilla-prefrontal
There are two points of contact between the maxilla and prefrontal, one on either side of the large foramen with a sloping ledge that accommodates the lacrimal canal. The most anterior seam begins at the anterior margin of the lacrimal foramen and follows the curved, but occasionally crenulated, outline of the facial process of the maxilla until it reaches the nasal. The second more posterior seam is essentially straight and runs between the posterior margin of the lacrimal duct and the junction with the palatine.
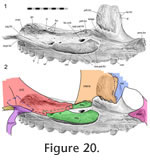 The anterior contact involves a large lateral facet on the prefrontal, which is overlapped by the majority of the facial process of the maxilla (Figure 17,
Figure 20). As mentioned above the prefrontal itself overlaps a portion of the nasal lateral facet and hence contributes to a triple bone overlap at this junction, although the skull is not exceptionally thick here. The prefrontal's lateral facet has anteroventrally directed striations, and one of these ends in a foramen. Similarly orientated, but less obvious, striations are visible on the facet of the maxilla (Figure 20). The anterior contact involves a large lateral facet on the prefrontal, which is overlapped by the majority of the facial process of the maxilla (Figure 17,
Figure 20). As mentioned above the prefrontal itself overlaps a portion of the nasal lateral facet and hence contributes to a triple bone overlap at this junction, although the skull is not exceptionally thick here. The prefrontal's lateral facet has anteroventrally directed striations, and one of these ends in a foramen. Similarly orientated, but less obvious, striations are visible on the facet of the maxilla (Figure 20).
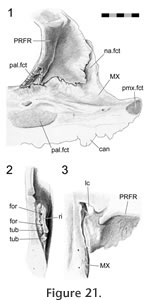 These internal striations parallel the anteroventrally directed grooves that can sometimes be seen on the external surface of the facial process of the maxilla. The ventral margin of the lateral facet (prefrontal) is dentate (Figures 17.1,
Figure 21.1), bearing planar triangular
projections which fit into corresponding depressions and recesses on the medial
surface of the maxilla (Figure 20,
Figure 21.1). The posterodorsal margin of the prefrontal facet is a deep wall which abuts and may occasionally overlap the dorsal margin of the maxilla very slightly. The external seam may also appear "slightly interdigitated" (sensu
Herring 1972, figure 1). In cross-section the joint most closely resembles a 'stepped joint' (Figure 2.3) but the seam's morphology and facet texture in some individuals also indicates some subtle Type-B interdigitation. This joint would prevent the facial process of the maxilla from rotating medially and would also restrict any posterodorsal movement. These internal striations parallel the anteroventrally directed grooves that can sometimes be seen on the external surface of the facial process of the maxilla. The ventral margin of the lateral facet (prefrontal) is dentate (Figures 17.1,
Figure 21.1), bearing planar triangular
projections which fit into corresponding depressions and recesses on the medial
surface of the maxilla (Figure 20,
Figure 21.1). The posterodorsal margin of the prefrontal facet is a deep wall which abuts and may occasionally overlap the dorsal margin of the maxilla very slightly. The external seam may also appear "slightly interdigitated" (sensu
Herring 1972, figure 1). In cross-section the joint most closely resembles a 'stepped joint' (Figure 2.3) but the seam's morphology and facet texture in some individuals also indicates some subtle Type-B interdigitation. This joint would prevent the facial process of the maxilla from rotating medially and would also restrict any posterodorsal movement.
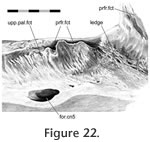 The posterior maxilla-prefrontal joint involves the posteroventral process of the prefrontal, which is associated with the maxilla-palatine and prefrontal-palatine joints (Figure
21). The ventrolateral edge of this process sits in a short groove on the dorsal surface of the maxilla just behind the facial process. The surface of the groove is not smooth in DGPC1 but bears two tubercles and two foramina (Figure 21.2). The groove is bounded medially by a small ridge (Figure 22). The fit is not tight and so may involve substantial soft tissue. The posterior maxilla-prefrontal joint involves the posteroventral process of the prefrontal, which is associated with the maxilla-palatine and prefrontal-palatine joints (Figure
21). The ventrolateral edge of this process sits in a short groove on the dorsal surface of the maxilla just behind the facial process. The surface of the groove is not smooth in DGPC1 but bears two tubercles and two foramina (Figure 21.2). The groove is bounded medially by a small ridge (Figure 22). The fit is not tight and so may involve substantial soft tissue.
Premaxilla-vomer
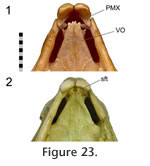 The right and left premaxilla-vomer joints can effectively be treated as a single horizontal joint between the paired premaxillae and paired vomers. The posterior surface of the conjoined premaxillae, cleaned of soft tissue, bears little evidence of its relationship with the vomers (e.g., DGPC1, DGPC2), and no facet is visible. The anterior end of each vomer bifurcates into two prongs but can be separated from the premaxilla by a notable distance (occasionally equal to the width of the vomerine anterior process e.g., NMNZ RE0385) (Figure 23.1). However, as seen in uncleaned skulls (e.g., LDUCZ x343, LDUCZ x1176) and CT data (YPM 9192), the posterior surfaces of the premaxillae and anterior tips of the vomers are connected by a thick sheet of soft tissue (Figure 23.2). This observation demonstrates the problems associated with inferring soft tissue from fossils. Each vomer also has a more posteriorly placed lateral prong that articulated with the small septomaxilla. The right and left premaxilla-vomer joints can effectively be treated as a single horizontal joint between the paired premaxillae and paired vomers. The posterior surface of the conjoined premaxillae, cleaned of soft tissue, bears little evidence of its relationship with the vomers (e.g., DGPC1, DGPC2), and no facet is visible. The anterior end of each vomer bifurcates into two prongs but can be separated from the premaxilla by a notable distance (occasionally equal to the width of the vomerine anterior process e.g., NMNZ RE0385) (Figure 23.1). However, as seen in uncleaned skulls (e.g., LDUCZ x343, LDUCZ x1176) and CT data (YPM 9192), the posterior surfaces of the premaxillae and anterior tips of the vomers are connected by a thick sheet of soft tissue (Figure 23.2). This observation demonstrates the problems associated with inferring soft tissue from fossils. Each vomer also has a more posteriorly placed lateral prong that articulated with the small septomaxilla.
Palatal Joints
 This palatal unit is the largest in the skull and is linked to the rostral unit by the paired maxillae, prefrontals and vomers, to the roofing unit by the prefrontals and to the temporal unit by the jugals and pterygoids. This palatal unit is the largest in the skull and is linked to the rostral unit by the paired maxillae, prefrontals and vomers, to the roofing unit by the prefrontals and to the temporal unit by the jugals and pterygoids.
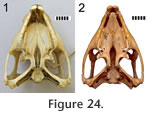
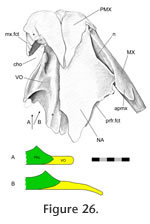
Vomer-palatine
The anterior tongue-like edge of the palatine overlaps the posterodorsal surface of the vomer (Figure 24). The facet on the vomer is scarfed with the slope directed anteromedially (Figure 25,
Figure 26).
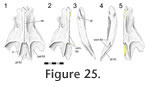 The amount of overlap depends on the vomer's posterior extent (which can be seen in ventral view). The amount of overlap depends on the vomer's posterior extent (which can be seen in ventral view).
 In some specimens the overlap is small with a vomerine-palatine ventral seam that runs anteromedially from the edge of the choana before turning posteromedially (LDUCZ x343 left; LDUCZ x036, left). In other specimens the overlap is larger where the ventral seam travels medially before turning sharply posteriorly to be parallel with the midline (e.g., LDUCZ x1176); larger again when the entire course is roughly posteromedial (LDUCZ x343; LDUCZ x146; LDUCZ x036, right; LDUCZ x343 right); and most extensive when the seam runs posteromedially before turning posteriorly to reach the junction with the pterygoids (DGPC2, NMNZ RE0385
Jones et al. 2009, figure 3.1). The facet on the vomer also includes a slot or pocket that accommodates the lateral edge of the anterior palatine process. The vomer laps the palatine early in ontogeny when the skull is no more than 6 mm in length (Howes and Swinnerton 1901, skull length = about 5.6 mm;
Werner 1962, about 6 mm, 5.4 mm according to
Bellairs and Kamal 1981, p. 118). In some specimens the overlap is small with a vomerine-palatine ventral seam that runs anteromedially from the edge of the choana before turning posteromedially (LDUCZ x343 left; LDUCZ x036, left). In other specimens the overlap is larger where the ventral seam travels medially before turning sharply posteriorly to be parallel with the midline (e.g., LDUCZ x1176); larger again when the entire course is roughly posteromedial (LDUCZ x343; LDUCZ x146; LDUCZ x036, right; LDUCZ x343 right); and most extensive when the seam runs posteromedially before turning posteriorly to reach the junction with the pterygoids (DGPC2, NMNZ RE0385
Jones et al. 2009, figure 3.1). The facet on the vomer also includes a slot or pocket that accommodates the lateral edge of the anterior palatine process. The vomer laps the palatine early in ontogeny when the skull is no more than 6 mm in length (Howes and Swinnerton 1901, skull length = about 5.6 mm;
Werner 1962, about 6 mm, 5.4 mm according to
Bellairs and Kamal 1981, p. 118).
Vomer-pterygoid
In Sphenodon the anterior tips of the paired pterygoids overlap the posterior ends of the paired vomers medially (Figure 25;
Jones et al. 2009, figure 3.2). This joint is separated from the vomer-palatine joint by a small ridge of bone (e.g., AIM LH0617; AUP 11883) but the posterior part underlies the palatine-pterygoid joint. In ventral view the vomer and pterygoid are separated by a short seam, which may be parallel to the coronal plane (LDUCZ x143), oblique to the sagittal plane (DGPC2) or 'V' shaped (LDUCZ x036).
Intervomerine
The ventral and dorsal seams for this midline joint are generally straight suggesting a simple butt joint (e.g.,
Jones et al. 2009). However, disarticulated material shows that, at least in some cases, it can be more complex. In specimen AIM LH0617, a small anteromedial shelf from the left vomer slots into a groove in the right vomer, centrally the medial edge of the left is overlapped slightly by the medial edge of the right, and posteriorly the medial edge of the right is overlapped by the left (Figure 25). In the central portion of the joint the medial margins of the vomers are dorsoventrally expanded, increasing the contact area between them.
Prefrontal-palatine
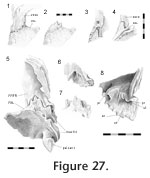 As previously reported the ventral process of the prefrontal meets the palatine with a wide joint (Bolt 1974). Viewed in posterodorsal aspect (through the orbit) this seam travels from its junction with the maxilla in a primarily dorsomedial direction. It is often interdigitated (e.g., LDUCZ x036) although not always (e.g., DGPC 2). Isolated bones demonstrate that this joint is extremely complex (Figure 27,
Figure 28). The anterolateral end of the palatine bears a facet facing posterodorsally at an angle of approximately 45° from the horizontal plane (level with the long axes of the maxillary tooth rows) (Figure 28). The posterior edge of the facet is recessed with a stepped (Figure 2.3) and jagged (zig-zagged) border (Type-B interdigitation,
Figure 2.4). The posterior process of the prefrontal sits in this sloping depression with a broadly corresponding posterior edge (Figure 27.1, 27.2). The anterior part of the facet on the palatine is also jagged, and this sits against a step on the underside of the prefrontal (Figure 27.6, 27.7). Visible in anteroventral view, a process from the prefrontal sits in a notch on the palatine medial to three projections from the latter (Figure 27.6, 27.7). The ventrolateral part of the ventral process of the prefrontal (that articulates with the maxilla) wraps around the palatine to form a longitudinal slot (Figure 27.6, 27.8). The facet on the prefrontal is somewhat striated laterally (Figure 27.8). As previously reported the ventral process of the prefrontal meets the palatine with a wide joint (Bolt 1974). Viewed in posterodorsal aspect (through the orbit) this seam travels from its junction with the maxilla in a primarily dorsomedial direction. It is often interdigitated (e.g., LDUCZ x036) although not always (e.g., DGPC 2). Isolated bones demonstrate that this joint is extremely complex (Figure 27,
Figure 28). The anterolateral end of the palatine bears a facet facing posterodorsally at an angle of approximately 45° from the horizontal plane (level with the long axes of the maxillary tooth rows) (Figure 28). The posterior edge of the facet is recessed with a stepped (Figure 2.3) and jagged (zig-zagged) border (Type-B interdigitation,
Figure 2.4). The posterior process of the prefrontal sits in this sloping depression with a broadly corresponding posterior edge (Figure 27.1, 27.2). The anterior part of the facet on the palatine is also jagged, and this sits against a step on the underside of the prefrontal (Figure 27.6, 27.7). Visible in anteroventral view, a process from the prefrontal sits in a notch on the palatine medial to three projections from the latter (Figure 27.6, 27.7). The ventrolateral part of the ventral process of the prefrontal (that articulates with the maxilla) wraps around the palatine to form a longitudinal slot (Figure 27.6, 27.8). The facet on the prefrontal is somewhat striated laterally (Figure 27.8).
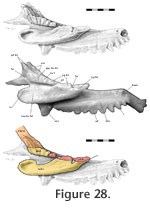 The prefrontal would be restrained from moving anteroventrally or medially and the palatine from moving posterodorsally or laterally. This joint would also stop the lower end of the prefrontal rotating anteriorly and the medial edge of the palatine from rotating laterally. The interlocking processes are not tight and would therefore allow some movement but the arrangement would provide a large surface area for soft tissue in many potential orientations. Overall this joint could be decribed as a stepped overlap with some Type-B interdigitation but some of the interlocking parallel to the bone surface also resembles a slot joint or Type-A interdigitation. The prefrontal would be restrained from moving anteroventrally or medially and the palatine from moving posterodorsally or laterally. This joint would also stop the lower end of the prefrontal rotating anteriorly and the medial edge of the palatine from rotating laterally. The interlocking processes are not tight and would therefore allow some movement but the arrangement would provide a large surface area for soft tissue in many potential orientations. Overall this joint could be decribed as a stepped overlap with some Type-B interdigitation but some of the interlocking parallel to the bone surface also resembles a slot joint or Type-A interdigitation.
Maxilla-palatine
In Sphenodon the palatine has two lateral processes separated by a foramen for the maxillary division of the trigeminal nerve (cranial nerve 5) and associated blood vessels. This foramen is marked "mf" in
Jones et al. (2009, figure 3.2). Both of these processes are involved in the joint with the maxilla (Figure 20,
Figure 28,
Figure 29).
 The upper maxilla-palatine joint is a loose butt contact with some very weak vertical Type-B interdigitation. In lateral view (disarticulated) the upper process of the palatine is rectangular and fluted, in DGPC1 this comprises five or six ridges which are directed posteroventrally in the anterior part and anteroventrally in the posterior part (Figure
28). Two grooves in this fluting probably relate to the presence of foramina. The corresponding facet on the maxilla for the upper lateral process of the palatine bears subtle anteroventrally directed striations (Figure 22). These striations are not obviously reflected on the palatine's maxillary facet. The upper maxilla-palatine joint is a loose butt contact with some very weak vertical Type-B interdigitation. In lateral view (disarticulated) the upper process of the palatine is rectangular and fluted, in DGPC1 this comprises five or six ridges which are directed posteroventrally in the anterior part and anteroventrally in the posterior part (Figure
28). Two grooves in this fluting probably relate to the presence of foramina. The corresponding facet on the maxilla for the upper lateral process of the palatine bears subtle anteroventrally directed striations (Figure 22). These striations are not obviously reflected on the palatine's maxillary facet.
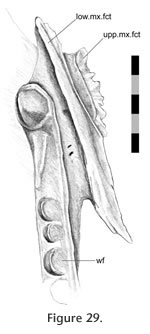
The lower maxilla-palatine joint is primarily a loose butt joint. The lower lateral process of the palatine stems from a point above the anterior half of the palatine tooth row (Figure 29).
 It expands anterolaterally and posteriorly to form a large process with an anterolaterally facing ovoid facet. This broad facet sits loosely against a depression in the maxilla almost dorsal to the maxillary tooth row so that the majority of the facet is orientated in between the long axis of the maxilla and long axis of the maxillary tooth row (Figure 5,
Figure 30). Ventral to the facet on the maxilla is a trough with a slight rim which is particularly distinct anteriorly (Figure 30). In life the space between the trough and lower palatine process is filled with soft tissue and the joint appears to be very well supported (e.g., LDUCZ x723). There is a distinct lack of sculpture on either the maxilla or palatine for the lower maxilla-palatine joint but anterodorsally the lower facet on the palatine bears a shallow tubercle and depression that corresponds to a depression and ridge on the maxilla (Figure 20,
Figure 28). The posterodorsal corners of both palatine processes contact the jugal and are discussed below. It expands anterolaterally and posteriorly to form a large process with an anterolaterally facing ovoid facet. This broad facet sits loosely against a depression in the maxilla almost dorsal to the maxillary tooth row so that the majority of the facet is orientated in between the long axis of the maxilla and long axis of the maxillary tooth row (Figure 5,
Figure 30). Ventral to the facet on the maxilla is a trough with a slight rim which is particularly distinct anteriorly (Figure 30). In life the space between the trough and lower palatine process is filled with soft tissue and the joint appears to be very well supported (e.g., LDUCZ x723). There is a distinct lack of sculpture on either the maxilla or palatine for the lower maxilla-palatine joint but anterodorsally the lower facet on the palatine bears a shallow tubercle and depression that corresponds to a depression and ridge on the maxilla (Figure 20,
Figure 28). The posterodorsal corners of both palatine processes contact the jugal and are discussed below.
Maxilla-jugal
 In dorsal view the seam arcs posterolaterally from the junction with the palatine to the margin of the orbit. In lateral view, the seam is sigmoid: running posteriorly from the edge of the orbit, turning first posteroventrally and then posteriorly again along the base of the lower temporal bar. In ventral view the seam is 'V'-shaped, running posteromedially and then anteromedially before meeting the ectopterygoid. In dorsal view the seam arcs posterolaterally from the junction with the palatine to the margin of the orbit. In lateral view, the seam is sigmoid: running posteriorly from the edge of the orbit, turning first posteroventrally and then posteriorly again along the base of the lower temporal bar. In ventral view the seam is 'V'-shaped, running posteromedially and then anteromedially before meeting the ectopterygoid.
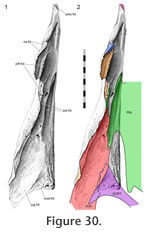
The jugal slots into a long concavity in the maxilla that broadens posteriorly and is bounded laterally by the sub-orbital margin (Figure
30). The long axis of the cavity is directed anteromedially but it is asymmetrical with a greater lateral component. At the posterior end of the concavity the lateral wall flexes medially and then laterally thus producing a longitudinal ridge (Figure
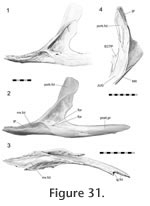
20) that slots into a wide groove along the lateral surface of the jugal (Figure 31).
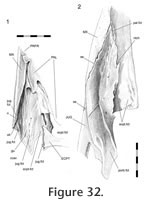 The ridge and groove are less pronounced in LDUCZ x1176 than in DGPC1 (Figure 32). Posteriorly this ridge has a rugose surface with convoluted striae and gutters directed anterodorsally, anteriorly and anteroventrally. Anteriorly the ridge is more sharply defined (at least in DGPC1) and shelf-like. Above it, anteriorly, there are three distinct slits (elongate foramina) (Figure
20). In dorsal view the base of the maxillary concavity can be seen to possess small gutters that are generally orientated anteromedially, particularly on the lateral side (Figure
30). Correspondingly the maxillary facet of the jugal is roughened, particularly in lateral view (Figure 31). The facet also displays several neuro-vascular foramina, some of which are at the anterior end of gutters. Two of these foramina, situated at the anterior end of the concavity, are very large (Figure
30). This joint would prevent anterior, lateral and ventral movement of the jugal and posterior, medial and dorsal movement of the maxilla. In a hatchling (e.g., FMNH 65905) the maxilla overlaps the jugal less extensively (Rieppel 1992). The ridge and groove are less pronounced in LDUCZ x1176 than in DGPC1 (Figure 32). Posteriorly this ridge has a rugose surface with convoluted striae and gutters directed anterodorsally, anteriorly and anteroventrally. Anteriorly the ridge is more sharply defined (at least in DGPC1) and shelf-like. Above it, anteriorly, there are three distinct slits (elongate foramina) (Figure
20). In dorsal view the base of the maxillary concavity can be seen to possess small gutters that are generally orientated anteromedially, particularly on the lateral side (Figure
30). Correspondingly the maxillary facet of the jugal is roughened, particularly in lateral view (Figure 31). The facet also displays several neuro-vascular foramina, some of which are at the anterior end of gutters. Two of these foramina, situated at the anterior end of the concavity, are very large (Figure
30). This joint would prevent anterior, lateral and ventral movement of the jugal and posterior, medial and dorsal movement of the maxilla. In a hatchling (e.g., FMNH 65905) the maxilla overlaps the jugal less extensively (Rieppel 1992).
Jugal-ectopterygoid
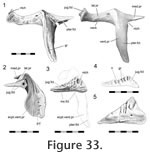 The ectopterygoid is a 'T'- shaped bone composed of a long ventral process, a slightly shorter lateral process and an even shorter medial process (Figure 33). The proximal portion of the lateral process was termed the 'neck' for the fossil rhynchocephalian Gephyrosaurus by
Evans (1980). The lateral process expands to become two-thirds the width of the neck and ends in a triangular face but there is variation in its exact dimensions as well as the surface texture. The face tends to being shallower in smaller individuals (Figure 33) and the dorsal edge may be ribbed and complex. The ectopterygoid is a 'T'- shaped bone composed of a long ventral process, a slightly shorter lateral process and an even shorter medial process (Figure 33). The proximal portion of the lateral process was termed the 'neck' for the fossil rhynchocephalian Gephyrosaurus by
Evans (1980). The lateral process expands to become two-thirds the width of the neck and ends in a triangular face but there is variation in its exact dimensions as well as the surface texture. The face tends to being shallower in smaller individuals (Figure 33) and the dorsal edge may be ribbed and complex.
This lateral process of the ectopterygoid plugs into a shallow depression on the medial surface of the jugal above the edge of the maxilla-jugal seam (Figure 34). The apex of the facet on the jugal coincides with the medial ridge of the postorbital bar (Figure 34). In DGPC1, the apex of this facet bears a small process that coincides with a groove on the ectopterygoid. As inferred from seam morphology this feature is also found in DGPC2, LDUCZ x146 (left) but not in LDUCZ x343, LDUCZ x036, or LDUCZ x146 (right). In a dorsal view of the articulated skull, the slightly crenulated seam extends posterolaterally from the anterior junction with the maxilla, and its posterior tip may turn ventromedially (DGPC1; DGPC2; LDUCZ x146, left). In posterior view, the seam passes ventrolaterally before turning ventromedially and meeting the maxilla. The section of seam visible in posterior view may also be interdigitated (e.g., LDUCZ x146, right).
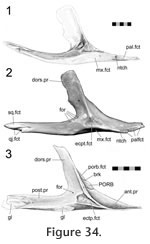 This joint would resist medial and dorsoventral movement of the jugal and conversely lateral and dorsoventral movement of the ectopterygoid. The contact appears strong with a large contact surface relative to the size of the ectopterygoid, anteriorly there is some slight interdigitation. This joint would resist medial and dorsoventral movement of the jugal and conversely lateral and dorsoventral movement of the ectopterygoid. The contact appears strong with a large contact surface relative to the size of the ectopterygoid, anteriorly there is some slight interdigitation.
Palatine-jugal
This joint primarily involves the anteromedial edge of the jugal abutting against the posterodorsal edge of the lower lateral palatine process (Figure 28,
Figure 32). In addition the tip of the jugal extends anteriorly beyond the maxillary foramen contacting the upper lateral palatine process (e.g., DGPC1, LDUCZ x723, LDUCZ x343 and possibly LDUCZ x1176) for a relatively short distance. Viewed dorsally, the majority of the seam runs posterolaterally but the posterior extremity of this seam turns medially.
The joint is essentially a perpendicular butt contact resisting medial movement of the jugal and lateral movement of the palatine. However, in DGPC1 (Figure 32.2,
Figure 34) the facet on the jugal is slightly concave and, as reflected in the seam, the posterior end of the joint is notched so that the jugal hooks behind the palatine (Figure 32). This arrangement of the bones would restrict posterior movement of the palatine and anterior movements of the jugal as well as some mediolateral movements. Apart from the upper facet of the palatine, which bears two vertical ridges, the facets involved lack obvious sculpture (Figure
28).
 Palatine-pterygoid Palatine-pterygoid
This joint is associated with the intervomerine, vomer-palatine, and vomer-pterygoid joints. In ventral view the seam generally runs posterolaterally from the junction with the vomer, although its anterior and posterior ends maybe more sagittally directed (Jones et al. 2009). In dorsal view the anterior end of the seam can be slightly convoluted and in posterior view the small posterior section of the joint has an 'S' shaped seam.
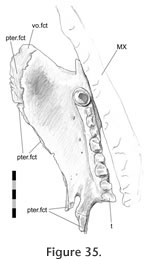
This joint can be divided into three parts. The anterior part involves the palatine overlapping the anterior processes of the pterygoid with a shallow scarf joint (Figure 2.5,
Figure 35) so that both palatines meet in the midline on the dorsal surface of the palate (e.g., DGPC2;
Sharrell 1966). However, the tips of the pterygoids remain exposed anteriorly (Jones et al. 2009, figure 3.2).
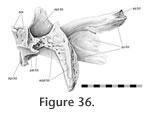 The central part of this joint involves the mediolateral edge of the palatine overlapping the lateral edge of the pterygoid but contact is generally minimal (Figure 35) and may even be lost entirely leaving an elongate fontanelle (e.g., LDUCZ x036 left side). In the posterior part of the joint the lateral pterygoid margin expands dorsoventrally and bears two slot-like recesses running oblique to the midline (Figure 36). Two posterior processes from the palatine insert against these recesses (Figure
37). The central part of this joint involves the mediolateral edge of the palatine overlapping the lateral edge of the pterygoid but contact is generally minimal (Figure 35) and may even be lost entirely leaving an elongate fontanelle (e.g., LDUCZ x036 left side). In the posterior part of the joint the lateral pterygoid margin expands dorsoventrally and bears two slot-like recesses running oblique to the midline (Figure 36). Two posterior processes from the palatine insert against these recesses (Figure
37).
 The contact lies just in front of the pterygoid-ectopterygoid joint and is related to the minor (or even occasional) palatine-ectopterygoid joint. The joint between the more medial process and slot is almost a simple butt contact (Figure 37.4). The joint between the more lateral process and slot is more complicated, in that the posterior tip of the process ends in a cup that wraps around the ventral edge of the slot (Figure 37.4). The contact lies just in front of the pterygoid-ectopterygoid joint and is related to the minor (or even occasional) palatine-ectopterygoid joint. The joint between the more medial process and slot is almost a simple butt contact (Figure 37.4). The joint between the more lateral process and slot is more complicated, in that the posterior tip of the process ends in a cup that wraps around the ventral edge of the slot (Figure 37.4).
In hatchlings the palatines do not appear to overlap the pterygoid extensively (Werner 1962) but the two posterior palatine processes are present in hatchling or near hatchling specimens (Howes and Swinnerton 1901;
Werner 1962).
Interpterygoid
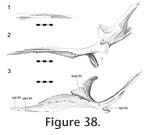 In Sphenodon the paired pterygoids are connected anteriorly along a midline seam which is less than half the total length of the bones (Figure 24.1). In ventral view the seam tends to exhibit low amplitude and long wavelength meandering (e.g., LDUCZ x036). The dorsal view is similar but the waveform may have greater amplitude. The internal structure can be clearly seen in BMNH.K and YPM11419. A medial view of a disarticulated pterygoid reveals long slots and flanges that are anterodorsally inclined from the long axis (Figure 38). These slots and flanges interlock as in Type-A interdigitation (Figure 2.2) but because the bones are relatively thin, contact seems small relative to the overall size of the bones. Therefore, despite being very distinct the joint is not necessarily strong. The meandering of the external seams corresponds to some Type-B interdigitation (Figure 2.4) but it is very subtle by comparison to the obvious Type-A interdigitation. In Sphenodon the paired pterygoids are connected anteriorly along a midline seam which is less than half the total length of the bones (Figure 24.1). In ventral view the seam tends to exhibit low amplitude and long wavelength meandering (e.g., LDUCZ x036). The dorsal view is similar but the waveform may have greater amplitude. The internal structure can be clearly seen in BMNH.K and YPM11419. A medial view of a disarticulated pterygoid reveals long slots and flanges that are anterodorsally inclined from the long axis (Figure 38). These slots and flanges interlock as in Type-A interdigitation (Figure 2.2) but because the bones are relatively thin, contact seems small relative to the overall size of the bones. Therefore, despite being very distinct the joint is not necessarily strong. The meandering of the external seams corresponds to some Type-B interdigitation (Figure 2.4) but it is very subtle by comparison to the obvious Type-A interdigitation.
Interpalatine
The thin dorsomedial edges of the paired palatines meet along the midline above the vomer-pterygoid junction (Sharrel 1966, p. 29;
Jones et al. 2009, figure 3). This arrangement can be appreciated with a dorsal view of the palate, provided the specimen is cleaned of soft tissue (e.g., in DGPC2;
Jones et al. 2009). In ventral view the joint is hidden by the underlying vomers and pterygoids.
 Pterygoid-ectopterygoid Pterygoid-ectopterygoid
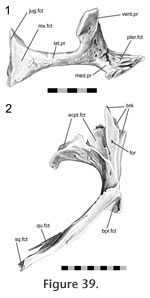
The contact surface is relatively large (described as "extensive" by
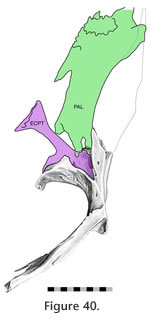
Günther [1867]), and involves both the ventral and medial processes of the ectopterygoid. The posterior surface of the ectopterygoid ventral process sits in a cavity in the descending process of the pterygoid to form the pterygoid flange (Figure 39). The ectopterygoid medial process is smaller than the lateral process but is expanded anteriorly and sits in a triangular recess on the dorsal surface of the pterygoid (Figure 40). The dorsal part of the joint is related to both the palatine-pterygoid joint and the negligible palatine-ectopterygoid joint.
Separating the medial process and the longer ventral process of the ectopterygoid is a small notch that accepts a small sill from the pterygoid. The ventral facet on the pterygoid is curved and bears both pitting and transverse ridging, particularly on its laterally facing surface. The ventral process of the ectopterygoid is curved and exhibits a subtle groove running along its length (Figure 33). Nevertheless the joint between the ectopterygoid and pterygoid is tightly apposed. It would resist anteroventral movements of the pterygoid and posterodorsal movements of the ectopterygoid. This arrangement would prevent the lateral ectopterygoid process from rotating posteriorly and the medial ectopterygoid process from rotating anteriorly but it would not prevent the opposite from occurring (i.e., anterior rotation of the lateral process and posterior rotation of the medial process).
 Palatine-ectopterygoid Palatine-ectopterygoid
In DGPC2, the anterior tip of the ectopterygoid's medial process rests against the dorsolateral surface of the lateral-most palatine projection. Consequently, small slivers of the ectopterygoid (laterally) and pterygoid (medially) are held between the two posterior projections of the palatine (Figure 35,
Figure 37). Only the tips of the posterior palatine projections are involved leaving a space between the bases (Figure 24.2;
Jones et al. 2009, figure 3). In specimen DGPC1 contact between the ectopterygoid and palatine is less certain but may have occurred indirectly through soft tissue related to the pterygoid-palatine joint.
Maxilla-ectopterygoid
The expanded lateral process of the ectopterygoid sits on the dorsal surface of the maxilla above the posterior end of the tooth row (Figure 20,
Figure 30). In dorsal view the seam travels posteromedially from the edge of the jugal. In disarticulated specimens the edges of the facet on the maxilla are not clearly delimited but within the location of the facet the surface bears pitting and ridges suggesting ligamentous tissue. However, this sculpture is not reflected on the ventral facet of the ectopterygoid which is smooth and may have occurred during preparation (Figure 30). This joint prevents the ectopterygoid from moving ventrally and the posterior end of the maxilla from any dorsal movement.
Roofing Joints
 The roofing joints are situated dorsally on the skull and mainly include those of the frontal, postfrontal and parietal. The roofing unit is linked to the rostral unit by the nasal and prefrontal, the palatal unit by the prefrontal, the temporal unit by the postorbital and squamosal, and the metakinetic unit by the parietal. The roofing joints are situated dorsally on the skull and mainly include those of the frontal, postfrontal and parietal. The roofing unit is linked to the rostral unit by the nasal and prefrontal, the palatal unit by the prefrontal, the temporal unit by the postorbital and squamosal, and the metakinetic unit by the parietal.
Nasal-frontal
 The naso-frontal seam generally runs posterolaterally from the midline to the junction with the prefrontal (e.g.,
Jones and Lappin 2009, figure 4;
Jones et al. 2009, figure 2). There is some variation in the exact shape of this seam as it may be nearly straight (e.g., BMNH 1844.102911; PCDG2 left side, LDUCZ x723 left side) or more sigmoid (e.g., LDUCZ x036, BMNH.K) travelling from the midline anterolaterally, posterolaterally, laterally and anterolaterally again. The naso-frontal seam generally runs posterolaterally from the midline to the junction with the prefrontal (e.g.,
Jones and Lappin 2009, figure 4;
Jones et al. 2009, figure 2). There is some variation in the exact shape of this seam as it may be nearly straight (e.g., BMNH 1844.102911; PCDG2 left side, LDUCZ x723 left side) or more sigmoid (e.g., LDUCZ x036, BMNH.K) travelling from the midline anterolaterally, posterolaterally, laterally and anterolaterally again.
 Correspondingly, in dorsal or ventral view, the posterior edge of isolated nasals may be either "V" shaped (e.g., PCDG2) or lobate (BMNH.K). Correspondingly, in dorsal or ventral view, the posterior edge of isolated nasals may be either "V" shaped (e.g., PCDG2) or lobate (BMNH.K).
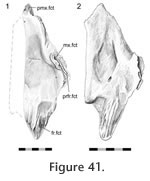
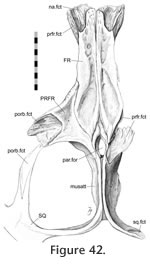
The posterolateral ends of the nasals overlap the anterolateral processes of the frontals with a scarfed tongue-in-groove joint (Figure 2.13,
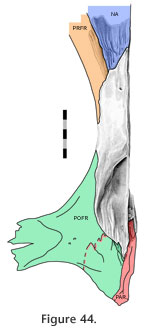
Figure 41,
Figure 42,
Figure 43,
Figure 44). In the disarticulated DGPC1 the medial portions of the bones are not available for study but the structure has been observed in other specimens (e.g., BMNH.K). The contact is particularly extensive along the junction with the prefrontal but diminishes medially (Figure 42,
Figure 44).
 Of 42 skulls examined, a notable midline fontanelle was present between the nasals and frontals in nine (21.4%: specimens AMPC 1, BMNH 1844.102911, KCL x12, LDUCZ x146, MANCH C.1206.49, NMNZ RE0382; OMNH 4911; UMZC 2613, UMZC 2614;
Jones and Lappin 2009;
Jones et al. 2009). In specimen MANCH C.1206.49 (skull length = 54.8 mm) this
fontanelle is 2.8 mm long and 1.5 mm wide. Of 42 skulls examined, a notable midline fontanelle was present between the nasals and frontals in nine (21.4%: specimens AMPC 1, BMNH 1844.102911, KCL x12, LDUCZ x146, MANCH C.1206.49, NMNZ RE0382; OMNH 4911; UMZC 2613, UMZC 2614;
Jones and Lappin 2009;
Jones et al. 2009). In specimen MANCH C.1206.49 (skull length = 54.8 mm) this
fontanelle is 2.8 mm long and 1.5 mm wide.
The anterolateral nasal facet on the frontal is generally scarfed, but it is also concave across its width so that the joint resembles a tongue-in-groove joint (Figure 42,
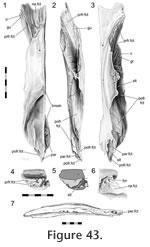
Figure 43,
Figure 44). Correspondingly the posterior process of the nasal is scarfed and convex across its width (Figure 13.1,
Figure 41). In DGPC1 the facet on the frontal possesses several long gutters and ridges that reflect similar texture on the facet of the nasal (Figure 41.1). These are generally orientated sagittally although they have some lateral inclination. Among the gutters there are also two large foramina. Other disarticulated nasals (e.g., BMNH.K, YPM 11419) possess a much smoother facet surface (Figure 41.2).
 This joint would resist dorsal or anterior movements of the frontal and ventral or posterior movements of the nasal. The mediolateral movements of both bones would also be inhibited by the concave shape of the frontal facet and also the sagittally orientated gutters. According to
Rieppel (1992), the nasals overlap the frontals extensively even in the hatchling. This joint would resist dorsal or anterior movements of the frontal and ventral or posterior movements of the nasal. The mediolateral movements of both bones would also be inhibited by the concave shape of the frontal facet and also the sagittally orientated gutters. According to
Rieppel (1992), the nasals overlap the frontals extensively even in the hatchling.
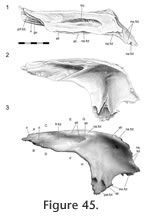
Frontal-prefrontal
In dorsal view the seam travels posteriorly from the nasal to the orbital margin along a roughly parasagittal line with a slight lateral inclination. In lateral view the seam travels posteroventrally before folding back anteroventrally in a "V' shape.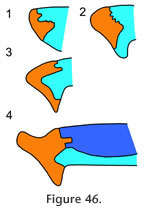 The frontal-prefrontal joint has two components, an anterior part and a posterior part (Figure 42,
Figure 44,
Figure 45,
Figure 46,
Figure 47). In the anterior part, the anterolateral process of the frontal fits inside a deep 'V'-shaped slot in the medial surface of the prefrontal posterior process (Figure 42,
Figure 44,
Figure 45,
Figure 46.3-4). In the posterior part, the posterior process of the prefrontal inserts into a deep cavity in the anterolateral surface of the frontal bone (Figure 45,
Figure 46.1-2,
Figure 47,
Figure 48). Longitudinal ridges can be found dorsally in both parts of this joint (Figure 43.1,
Figure 43.3,
Figure 45.1). The frontal-prefrontal joint has two components, an anterior part and a posterior part (Figure 42,
Figure 44,
Figure 45,
Figure 46,
Figure 47). In the anterior part, the anterolateral process of the frontal fits inside a deep 'V'-shaped slot in the medial surface of the prefrontal posterior process (Figure 42,
Figure 44,
Figure 45,
Figure 46.3-4). In the posterior part, the posterior process of the prefrontal inserts into a deep cavity in the anterolateral surface of the frontal bone (Figure 45,
Figure 46.1-2,
Figure 47,
Figure 48). Longitudinal ridges can be found dorsally in both parts of this joint (Figure 43.1,
Figure 43.3,
Figure 45.1).
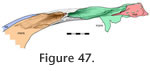 Anteriorly the joint is associated with the nasal-prefrontal and nasal-frontal joints (Figure 46). Both components of the joint combine to form an alternating slot joint. This joint would resist anterior movement of the frontal and posterior movement of the prefrontal but would also prevent differential dorsoventral movements between the bones. Anteriorly the joint is associated with the nasal-prefrontal and nasal-frontal joints (Figure 46). Both components of the joint combine to form an alternating slot joint. This joint would resist anterior movement of the frontal and posterior movement of the prefrontal but would also prevent differential dorsoventral movements between the bones.
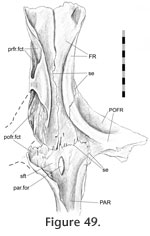 Interfrontal Interfrontal
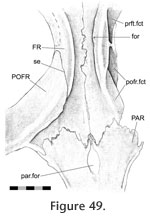 In Sphenodon the frontals generally contact each other sagittally with a long butt joint. Ventrally the seam is generally straight except where it exhibits low amplitude interdigitation centrally between the orbital margins (e.g., LDUCZ x1176, BMNH K, UCMZ2614; AUP 11883) (Figure 48,
Figure 49). Correspondingly, in medial view the mid ventral portion of the facet bears ridges oblique to the long axis of the bone (Figure 43.7) representing some degree of Type-B interdigitation (YPM11419). The interfrontal
suture fully closes in hatchlings between stages S and T when the skull is 9 14
mm long (Howes and Swinnerton 1901;
Schauinsland 1903;
Werner 1962;
Rieppel 1992;
Jones and Lappin 2009). In Sphenodon the frontals generally contact each other sagittally with a long butt joint. Ventrally the seam is generally straight except where it exhibits low amplitude interdigitation centrally between the orbital margins (e.g., LDUCZ x1176, BMNH K, UCMZ2614; AUP 11883) (Figure 48,
Figure 49). Correspondingly, in medial view the mid ventral portion of the facet bears ridges oblique to the long axis of the bone (Figure 43.7) representing some degree of Type-B interdigitation (YPM11419). The interfrontal
suture fully closes in hatchlings between stages S and T when the skull is 9 14
mm long (Howes and Swinnerton 1901;
Schauinsland 1903;
Werner 1962;
Rieppel 1992;
Jones and Lappin 2009).
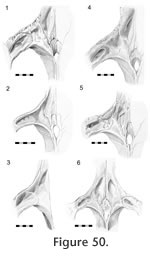 Frontal-postfrontal Frontal-postfrontal
In dorsal view the seam for this joint can vary dramatically between specimens (Figure 50).
 From the anterior border of the orbit, the seam travels posteromedially toward the junction with the parietal but may do so in a straight line (e.g., LDUCZ x036), a broad 'V'-shaped line (e.g., UMZC 2583, LDUCZ 1176), a gentle curve (e.g., UMZC 2614) a broad curve (e.g., BMB 101806, BMNH 19851212, LDUCZ x146, DGPC1, BMNH.K), a sigmoid curve (e.g., UMZC 2613, BMB 100225, NMNZ0382) or a very sigmoid curve (e.g., UMZC 2593, DGPC2). In a lateral view of the whole skull, the anterior segment of the seam travels anterodorsally from the orbital margin before turning posterodorsally at mid length. From the anterior border of the orbit, the seam travels posteromedially toward the junction with the parietal but may do so in a straight line (e.g., LDUCZ x036), a broad 'V'-shaped line (e.g., UMZC 2583, LDUCZ 1176), a gentle curve (e.g., UMZC 2614) a broad curve (e.g., BMB 101806, BMNH 19851212, LDUCZ x146, DGPC1, BMNH.K), a sigmoid curve (e.g., UMZC 2613, BMB 100225, NMNZ0382) or a very sigmoid curve (e.g., UMZC 2593, DGPC2). In a lateral view of the whole skull, the anterior segment of the seam travels anterodorsally from the orbital margin before turning posterodorsally at mid length.
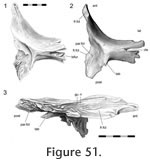
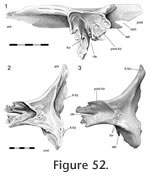 This joint primarily involves the medial surface of the postfrontal slotting into the lateral surface of the frontal (Figure 47,
Figure 48,
Figure 49,
Figure 50,
Figure 51,
Figure 52,
Figure 53,
Figure 54) but it can be divided into three sections. This joint primarily involves the medial surface of the postfrontal slotting into the lateral surface of the frontal (Figure 47,
Figure 48,
Figure 49,
Figure 50,
Figure 51,
Figure 52,
Figure 53,
Figure 54) but it can be divided into three sections.
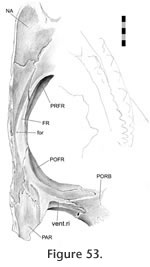 In the anterior section, the anterior process of the postfrontal (Figure 51,
Figure 52.2) inserts into a deep slot
in the frontal (Figure 43,
Figure 47,
Figure 48,
Figure 49,
Figure 54). This slot exhibits gutters and ridges running along its axis (Figure 43,
Figure 48). In the central and largest section the frontal sits in a deep concavity on the postfrontal (Figure 52.2), and some specimens possess a small shelf on the frontal that enters the postfrontal (BMNH K) (Figure 42). The posterior section is related to the anterior joints of the parietal. In LDUCZ x1176 this involves a thin flat process from the frontal slotting into the postfrontal (Figure 55). Alternatively, as in DGPC1, the postfrontal may overlap a small posterolateral sliver of the frontal (Figure 43.1,
Figure 44,
Figure 51.1) and posteriorly the posterior tip of the frontal overlaps a small triangular shelf extending medially from the postfrontal (Figure 43.3,
Figure 54). In the anterior section, the anterior process of the postfrontal (Figure 51,
Figure 52.2) inserts into a deep slot
in the frontal (Figure 43,
Figure 47,
Figure 48,
Figure 49,
Figure 54). This slot exhibits gutters and ridges running along its axis (Figure 43,
Figure 48). In the central and largest section the frontal sits in a deep concavity on the postfrontal (Figure 52.2), and some specimens possess a small shelf on the frontal that enters the postfrontal (BMNH K) (Figure 42). The posterior section is related to the anterior joints of the parietal. In LDUCZ x1176 this involves a thin flat process from the frontal slotting into the postfrontal (Figure 55). Alternatively, as in DGPC1, the postfrontal may overlap a small posterolateral sliver of the frontal (Figure 43.1,
Figure 44,
Figure 51.1) and posteriorly the posterior tip of the frontal overlaps a small triangular shelf extending medially from the postfrontal (Figure 43.3,
Figure 54).
Frontal-parietal
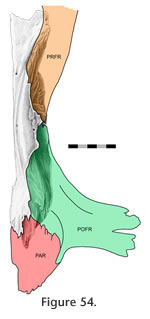 Elsewhere this cranial joint suture is sometimes referred to as the coronal suture, reflecting human terminology (e.g.,
Moss 1954,
1957;
Markens and Oudhof 1980;
Opperman 2000).The
joint is complex and is associated with the medial joints of the postfrontal. Primarily
this joint involves an alternating overlap; laterally the frontal overlaps the
parietal but medially the parietal overlaps the frontal Elsewhere this cranial joint suture is sometimes referred to as the coronal suture, reflecting human terminology (e.g.,
Moss 1954,
1957;
Markens and Oudhof 1980;
Opperman 2000).The
joint is complex and is associated with the medial joints of the postfrontal. Primarily
this joint involves an alternating overlap; laterally the frontal overlaps the
parietal but medially the parietal overlaps the frontal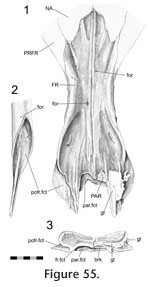 (Figure
42, Figure 43,
Figure 44,
Figure 48,
Figure 49,
Figure 50,
Figure 53,
Figure 54,
Figure 53,
Figure 54,
Figure 56,
Figure 57,
Figure 58,
Figure 59). (Figure
42, Figure 43,
Figure 44,
Figure 48,
Figure 49,
Figure 50,
Figure 53,
Figure 54,
Figure 53,
Figure 54,
Figure 56,
Figure 57,
Figure 58,
Figure 59).
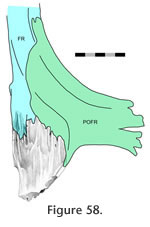
In dorsal view the external seam is short (Figure 50;
Günther 1867;
Arnold 1998;
Evans 2008;
Jones and Lappin 2009;
Jones et al. 2009). From the midline it may travel posterolaterally around the anterior margin of the parietal foramen before meeting the junction with the postfrontal (Figure 50.6; e.g., LDUCZ x036, UMZC 2613, UMZC 2611). Alternatively it may be more sigmoid, at first travelling laterally or anterolaterally before turning posteriorly and finally curving posterolaterally (Figure 50.2; e.g., LDUCZ x1176, UMZC 2582, UMZC 2610, NMNZ RE0382). The frontal has two posterior processes (Figure
43), a lateral process that overlaps the
 parietal (Figure 43.3,
Figure 54) and a medial process that is overlapped by the parietal (Figure 43.1,
Figure 44,
Figure 53,
Figure 55,
Figure 58,
Figure 59). The two processes are separated by an oblique slot (Figure 43.5) that may be smaller in juvenile frontals (e.g., LDUCZ x1176). Occasionally the posteriormost parietal (Figure 43.3,
Figure 54) and a medial process that is overlapped by the parietal (Figure 43.1,
Figure 44,
Figure 53,
Figure 55,
Figure 58,
Figure 59). The two processes are separated by an oblique slot (Figure 43.5) that may be smaller in juvenile frontals (e.g., LDUCZ x1176). Occasionally the posteriormost
 medial edge of the frontal is exposed dorsally (e.g., NMNZ RE0382) but otherwise the overlapping facets of both bones are scarfed, and therefore this resembles the 'birds-mouth' joint found in wood joinery (Figure 2.15;
Graubner 1992). On its own, without soft tissue, this joint would resist all movements except perhaps anterior movement of the frontal and posterior movement of the parietal. medial edge of the frontal is exposed dorsally (e.g., NMNZ RE0382) but otherwise the overlapping facets of both bones are scarfed, and therefore this resembles the 'birds-mouth' joint found in wood joinery (Figure 2.15;
Graubner 1992). On its own, without soft tissue, this joint would resist all movements except perhaps anterior movement of the frontal and posterior movement of the parietal.
In hatchlings the adjoining medial portions of the frontals and parietals have not fully ossified, resulting in a fronto-parietal fontanelle (Howes and Swinnerton 1901;
Rieppel 1992). It is closed in specimens with skull lengths approaching 15 mm (Howes and Swinnerton 1901;
Jones and Lappin 2009).
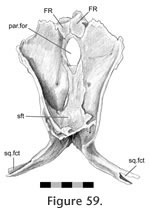
 Postfrontal-parietal Postfrontal-parietal
This is the largest joint of the parietal and overall involves the posterior end of the postfrontal overlapping the anterolateral part of the parietal (Figure 42,
Figure 44,
Figure 47,
Figure 48,
Figure 49,
Figure 50,
Figure 51,
Figure 52,
Figure 53,
Figure 56,
Figure 57,
Figure 58,
Figure 59). In dorsal view the seam runs posteromedially from the junction with the frontal for a short distance, occasionally reaching the anterior edge of the parietal crest at a point level with the posterior end of the parietal foramen (Figure
50). Here, it curves around, and the seam continues anterolaterally.
 It may continue this course (e.g., LDUCZ x1176, LDUCZ x723 right side) or it may fold back posteriorly (e.g., LDUCZ x036; LDUCZ x343 left side, DGPC1). Variation in the posterior extent of the postfrontals can occur even within the same individual (Figure 50; e.g., DGPC2, LDUCZ x036). It may continue this course (e.g., LDUCZ x1176, LDUCZ x723 right side) or it may fold back posteriorly (e.g., LDUCZ x036; LDUCZ x343 left side, DGPC1). Variation in the posterior extent of the postfrontals can occur even within the same individual (Figure 50; e.g., DGPC2, LDUCZ x036).
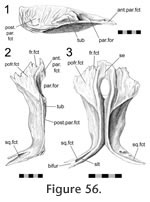
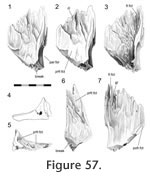
Disarticulation of the skull bones demonstrates that the postfrontal overlaps the broad anterolateral expansion of the parietal (Figure 56,
Figure 57). The expansion is concave, and aligned along the contours are several gutters (Figure
57), ridges and tubercles that slot into complementary features on the postfrontal. Therefore the structure corresponds to a scarf joint (Figure 2.5) with some Type-A interdigitation (Figure 2.2). Often a splint or tab of bone from the posterolateral corner of the postfrontal (Figure 51,
Figure 52.1) underlaps the anterolateral edge of the parietal (e.g., DGPC1, LDUCZ x036) (Figure
58). This joint would resist any significant dorsal movement of the parietal and ventral movement of the postfrontal. It would also inhibit anterolateral movements of the parietal and posteromedial movements of the postfrontal.
Postorbital-postfrontal
The postorbital and postfrontal of Sphenodon are both fairly robust and together form the upper part of the postorbital bar (Günther 1867;
Evans 2008;
Jones et al. 2009). The bar has a dorsal surface, an anteroventral surface and a posteroventral surface, with the latter two separated by the ventral ridge (Figure
53). The articulation is a complex, and tightly fitting, slot joint between the medial process of the postorbital and the lateral process of the postfrontal (Figure 42,
Figure 51,
Figure 52,
Figure 53,
Figure 60).
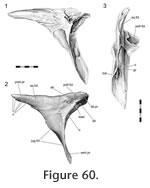 The seam demonstrates intraspecific variation but overall it travels around the central part of the upper postorbital bar in a zig-zag fashion. On the dorsal surface the seam arcs medially from the edges of the upper postorbital bar. The shape of this arc is subject to variation (Figure 50); it may be broadly 'U'-shaped (Figure 50.1, 50.2, 50.3;
Jones et al. 2009, figure 2.2; e.g., LDUCZ x146, LDUCZ x721, LDUCZ x723, LDUCZ x1176, NMNZ RE0382; NMNZ RE2509) or more 'V'-shaped (Figure 50.4, 50.5, 50.6; e.g., DGPC 1, LDUCZ x036, LDUCZ x343, BMNH1985.1212, YPM9194), demonstrating varying degrees of symmetry. On the posteroventral surface the seam travels ventrolaterally before folding back ventromedially in a 'V' shape (e.g., LDUCZ x1176), or more irregularly (e.g., DGPC1). The seam continues ventromedially until a point just The seam demonstrates intraspecific variation but overall it travels around the central part of the upper postorbital bar in a zig-zag fashion. On the dorsal surface the seam arcs medially from the edges of the upper postorbital bar. The shape of this arc is subject to variation (Figure 50); it may be broadly 'U'-shaped (Figure 50.1, 50.2, 50.3;
Jones et al. 2009, figure 2.2; e.g., LDUCZ x146, LDUCZ x721, LDUCZ x723, LDUCZ x1176, NMNZ RE0382; NMNZ RE2509) or more 'V'-shaped (Figure 50.4, 50.5, 50.6; e.g., DGPC 1, LDUCZ x036, LDUCZ x343, BMNH1985.1212, YPM9194), demonstrating varying degrees of symmetry. On the posteroventral surface the seam travels ventrolaterally before folding back ventromedially in a 'V' shape (e.g., LDUCZ x1176), or more irregularly (e.g., DGPC1). The seam continues ventromedially until a point just
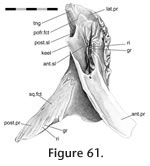 before the ventral ridge where it turns ventrolaterally. On the anteroventral surface the seam continues ventrolaterally before redirecting dorsomedially. This redirection can occur almost immediately (LDUCZ x036, LDUCZ x146, DGPC2, YPM9194), in a simple 'V' shape (e.g., LDUCZ x1176), or in a lobed curve (DGPC1, LDUCZ x723). before the ventral ridge where it turns ventrolaterally. On the anteroventral surface the seam continues ventrolaterally before redirecting dorsomedially. This redirection can occur almost immediately (LDUCZ x036, LDUCZ x146, DGPC2, YPM9194), in a simple 'V' shape (e.g., LDUCZ x1176), or in a lobed curve (DGPC1, LDUCZ x723).
The internal part of the joint can be divided into a dorsal part and a ventral part. The dorsal part consists of a tongue-in-groove joint where the tongue-shaped medial process of the postorbital (Figure 60,
Figure 61) sits in a concavity on the postfrontal (Figure 2.13,
Figure 42,
Figure 52,
Figure 62). The ventral part of this joint consists of a triple vertical slot joint, with two slots in the postorbital (Figure 2.11,
Figure 60,
Figure 61) and one in the postfrontal (Figure 51,
Figure 52,
Figure 53). The latter slot is here referred to as the postfrontal cleft. Its exact position can vary, and it may be absent altogether (e.g., Figure 42, BMNH.K, AIM LH 833) but it is usually found at the base of the postfrontal concavity with a long axis parallel to that of the upper postorbital bar. This cleft accepts a narrow keel of bone that projects ventrally from the postorbital (Figure 60,
Figure 61). When present the postorbital cleft divides the lateral end of the postfrontal into anterolateral and posterolateral processes as suggested by the postorbital-postfrontal seam (Figure
52). It is these two processes that insert into the two slots on the postorbital. The postfrontal anterolateral process is slightly larger than the dorsolateral process and slots into an anterior cavity against the postorbital keel. The ceiling of the cavity bears a long ridge (Figure 61) complimentary to a groove on the anterolateral process of the postfrontal (Figure 52.2). Smaller tubercles and ridges are also present. In lateral view a fairly large foramen can be seen at the proximal end of one of the gutters (Figure 52.1). The tip of the postfrontal posterolateral process also bifurcates (Figure 51.1) around a small process inside the posteromedial slot of the postorbital (Figure 60.1).
Even without soft tissue, the only movements permitted by this joint are medial movement of the postfrontal and lateral movement of the postorbital. The dorsal tongue-in-groove and ventral vertical slot joint would resist lateral movement of the postfrontal, medial movement of the postorbital and anteroposterior movements of either bone. Dorsoventral movements are restricted by the shape of the slots in the postorbital. In addition, the tongue-in-groove would obstruct dorsal movements of the postfrontal and ventral movements of the postorbital. The keel, ridges and slots would all increase the potential surface area for soft tissue attachment.
In a large hatchling Sphenodon (skull length = 12.3 mm) the postorbital overlaps the postfrontal to a similar extent to that found in adults (Rieppel 1992) but in smaller skulls the overlap is less extensive (Howes and Swinnerton 1901, skull length = 9 mm).
Interparietal
This cranial joint is sometimes referred to as the sagittal suture of human terminology (e.g.,
Moss 1954,
1957;
Opperman 2000,
Byron et al. 2004). In Sphenodon, the parietals contact each other along the midline with a straight seam either side of the parietal foramen, and form a raised crest (Figure 14). They are not necessarily fused (contra
Evans 1980, p. 240) although they can often be very firmly apposed (Figure 14.1; e.g., DGPC2, LDUCZ x036, LDUCZ x723). In other individuals, however, the two halves of the crest are separated dorsally or bifurcated (e.g., LDUCZ x146, LDUCZ x721; LDUCZ x343). As this bifurcation can be found in small skulls (e.g., LDUCZ x1176) it is unlikely that it relates to presence or absence of the large scaly "nuchal crest" found in adult males (e.g.,
Gillingham et al. 1995;
Parkinson 2002). In posterior view the external seam can appear sigmoid (LDUCZ x036).
Examination of isolated specimens demonstrates that this joint is essentially a large butt joint. However, the medial surface of each parietal is dorsoventrally expanded so that the contact surface is two or three times deeper than that of the interfrontal or internasal (Figure 56.1). Also, the facet surfaces are not entirely smooth. For example, the left parietal of YPM 11419 has a large anterior bony tubercle anteriorly (Figure 56.1) that inserts into a corresponding depression on the right parietal. Posteroventrally directed striations are also visible above the tubercle and posterior to the parietal foramen. These may serve to resist some anteroposterior movements. The sigmoid shape of the seam, often visible posteriorly, also shows that a small process from one parietal inserts into a small depression in the posterior edge of the other.
In hatchlings the parietals make contact along the midline after stage S and before stage T when the skull is about 10 mm long and most of it is ossified (Howes and Swinnerton 1901;
Rieppel 1992). Before their contact, the parietals are separated by a large central space (Howes and Swinnerton 1901;
Schauinsland 1903;
Werner 1962).
Parietal-squamosal
The medial process of the squamosal sits in the bifurcating end of the parietal bone (Figure 56,
Figure 59). The latter is asymmetrically 'U' shaped in cross-section, with the posterior process being taller than the anterior process by about half. This joint resists anteroposterior movements of both bones, medial and ventral movements of the squamosal, lateral and dorsal movements of the parietal. Some very minor mediolateral movement may have been permitted to relieve local tension caused by contraction of the adductor musculature.
Parietal-supratemporal
The supratemporal is unknown in adult Sphenodon although a tiny sliver representing it was tentatively identified in a hatchling specimen (Rieppel 1992).
Temporal Joints
The temporal joints surround the lower temporal fenestra. The temporal unit is linked to the palatal unit by the jugal and pterygoid, the roofing unit by the postorbital and squamosal, and the metakinetic unit by the pterygoid and quadrate.
Jugal-postorbital
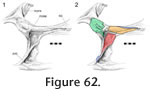 The postorbital sits on top of and against the dorsal process of the jugal. In lateral view the seam is diagonal, sloping backwards with a slight sigmoid shape (Figure 62.1;
Günther 1867;
Evans 2008;
Jones 2008;
Jones et al. 2009). The joint is composed of a single but twisted articulating surface that can be separated into upper and lower parts. In the lower parts the facet on the jugal faces anteromedially (Figure 31.4,
Figure 32.2,
Figure 34.3,
Figure 62) and provides a shallow depression that accommodates the postorbital. In the upper portion of the joint the jugal expands dorsally alongside the medial surface of the postorbital (Figure 62,
Figure 63). In the juvenile LDUCZ x1176, the tip of the jugal dorsal process tapers rather than being "squared-off" as in the adult Sphenodon examined (Figure 32.2,
Figure 34). The postorbital sits on top of and against the dorsal process of the jugal. In lateral view the seam is diagonal, sloping backwards with a slight sigmoid shape (Figure 62.1;
Günther 1867;
Evans 2008;
Jones 2008;
Jones et al. 2009). The joint is composed of a single but twisted articulating surface that can be separated into upper and lower parts. In the lower parts the facet on the jugal faces anteromedially (Figure 31.4,
Figure 32.2,
Figure 34.3,
Figure 62) and provides a shallow depression that accommodates the postorbital. In the upper portion of the joint the jugal expands dorsally alongside the medial surface of the postorbital (Figure 62,
Figure 63). In the juvenile LDUCZ x1176, the tip of the jugal dorsal process tapers rather than being "squared-off" as in the adult Sphenodon examined (Figure 32.2,
Figure 34).
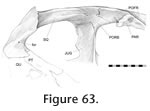 In DGPC1, the articulation surface bears subtle striations aligned with its long axis (Figure 31). However, the only substantial texture is located on the postorbital in the mid-part of the joint. It consists of a tubercle surrounded by gutters and ridges (Figure 60.3). Overall, the joint has an extensive surface area for soft tissue with overlap occurring along the full height of the postorbital bar. Primarily, it would restrict dorsoanteromedial movement of the jugal and ventroposterolateral movement of the postorbital. The medial seam of the joint crosses the medial ridge of the postorbital and jugal which itself is a continuation of the ventral ridge found on the postfrontal. In DGPC1, the articulation surface bears subtle striations aligned with its long axis (Figure 31). However, the only substantial texture is located on the postorbital in the mid-part of the joint. It consists of a tubercle surrounded by gutters and ridges (Figure 60.3). Overall, the joint has an extensive surface area for soft tissue with overlap occurring along the full height of the postorbital bar. Primarily, it would restrict dorsoanteromedial movement of the jugal and ventroposterolateral movement of the postorbital. The medial seam of the joint crosses the medial ridge of the postorbital and jugal which itself is a continuation of the ventral ridge found on the postfrontal.
Available images of hatchling skulls suggest there is overlap at this joint but it is not as extensive as in adult skulls (Figure 4;
Rieppel 1992;
Werner 1962).
Jugal-quadratojugal
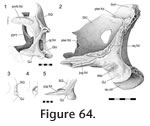 In Sphenodon the posterior process of the jugal generally overlaps the anterior process of the quadratojugal but the exact nature of this overlap varies intraspecifically (Figure 64). In lateral view, the seam travels posterodorsally to the junction with the squamosal (Jones et al. 2009). In ventral view the seam is generally posterolaterally transverse. However, it is variable, and may differ in individuals between left and right sides. It may be straight (e.g., LDUCZ x723 right side, LDUCZ x1176, LDUCZ x036, LDUCZ x146), sigmoid (e.g., DGPC1, DGPC2) or 'V' shaped (e.g., LDUCZ x723 left side). In DGPC1 most of the quadratojugal lateral facet is 'L' shaped in coronal section, and the posterior process of the jugal sits on the sill of the 'L'. As a result the posterior end of the jugal overlaps the quadratojugal for a length two or three times the height of the lower temporal bar. In ventral view the quadratojugal generally overlaps the jugal but occasionally along the medial edge of the lower temporal bar, the jugal may overlap a portion of the quadratojugal (DGPC1). The joint has additional features in some larger individuals and so may change with ontogeny. The features include interlocking pegs anteroventrally (e.g., DGPC1) so that the joint as a whole can resemble an asymmetrical 'birds mouth joint' (Figure 2.15;
Graubner 1998). Furthermore in DGPC1, at least, the facet on the quadratojugal bears a slight ridge that fits into a groove on the jugal. It should be noted that the shape of the jugal posterior process (and thus also the lower temporal bar) is variable; both in terms of its relative dorsoventral dimension (Jones 2008) but also its mediolateral dimension. In coronal section, the process can be twice as tall as wide (Figure 64.3; e.g., LDUCZ x146) or it may be nearly circular with equal width and height (Figure 64.4; e.g., DGPC1). Where the lower temporal bar is very circular in cross-section the posterior process of the jugal is acuminate, and the corresponding joints with the quadratojugal and squamosal are more slot-like. When the lower temporal bar is tall in cross-section (YPM 11419), the suture is more likely to be a simple scarfed overlap (Figure 2.5). In Sphenodon the posterior process of the jugal generally overlaps the anterior process of the quadratojugal but the exact nature of this overlap varies intraspecifically (Figure 64). In lateral view, the seam travels posterodorsally to the junction with the squamosal (Jones et al. 2009). In ventral view the seam is generally posterolaterally transverse. However, it is variable, and may differ in individuals between left and right sides. It may be straight (e.g., LDUCZ x723 right side, LDUCZ x1176, LDUCZ x036, LDUCZ x146), sigmoid (e.g., DGPC1, DGPC2) or 'V' shaped (e.g., LDUCZ x723 left side). In DGPC1 most of the quadratojugal lateral facet is 'L' shaped in coronal section, and the posterior process of the jugal sits on the sill of the 'L'. As a result the posterior end of the jugal overlaps the quadratojugal for a length two or three times the height of the lower temporal bar. In ventral view the quadratojugal generally overlaps the jugal but occasionally along the medial edge of the lower temporal bar, the jugal may overlap a portion of the quadratojugal (DGPC1). The joint has additional features in some larger individuals and so may change with ontogeny. The features include interlocking pegs anteroventrally (e.g., DGPC1) so that the joint as a whole can resemble an asymmetrical 'birds mouth joint' (Figure 2.15;
Graubner 1998). Furthermore in DGPC1, at least, the facet on the quadratojugal bears a slight ridge that fits into a groove on the jugal. It should be noted that the shape of the jugal posterior process (and thus also the lower temporal bar) is variable; both in terms of its relative dorsoventral dimension (Jones 2008) but also its mediolateral dimension. In coronal section, the process can be twice as tall as wide (Figure 64.3; e.g., LDUCZ x146) or it may be nearly circular with equal width and height (Figure 64.4; e.g., DGPC1). Where the lower temporal bar is very circular in cross-section the posterior process of the jugal is acuminate, and the corresponding joints with the quadratojugal and squamosal are more slot-like. When the lower temporal bar is tall in cross-section (YPM 11419), the suture is more likely to be a simple scarfed overlap (Figure 2.5).
This joint would have resisted lateral and dorsal movement of the quadrate and medial and ventral movements of the jugal. The ventral pegs would also resist anterior movement of the quadratojugal and posterior movement of the jugal.
As known for over a hundred years, the lower temporal bar is incomplete early in ontogeny (e.g.,
Howes and Swinnerton 1901;
Schauinsland 1903;
De Beer 1937;
Evans 1980;
Bellairs and Kamal 1981;
Whiteside 1986;
Rieppel 1994;
Müller 2003). In a relatively large hatchling (e.g., FMNH 65905, skull length = 12.3 mm) the jugal extends posteriorly and overlaps the quadratojugal but the contact is loose (Rieppel 1992). In smaller hatchlings (Werner 1962 skull length ~ 6 mm) and more clearly in the embryos, the jugal posterior process is only partly developed and does not reach the quadratojugal (Howes and Swinnerton 1901;
Schauinsland 1903, skull lengths < 6 mm).
Jugal-squamosal
In the lower temporal bar of adult Sphenodon, the posterior process of the jugal overlaps the ventral process of the squamosal above the larger jugal-quadratojugal joint. In a lateral view the seam generally runs anterodorsally from the junction with the quadratojugal; in medial view the seam runs anteroventrally from the dorsal crest of the lower temporal bar to the junction with the quadratojugal. As with the jugal-quadratojugal seam there is some individual variation. In lateral view the seam may be sigmoid (e.g., LDUCZ x146) or it may run posterodorsally before redirecting anterodorsally because the posterior tip of the jugal lies against the squamosal rather than at the junction with the quadratojugal. For example on the right side of LDUCZ x036 and left side in OMNH 908 (as shown in Jones [2008] but modified in
Jones et al. [2009]). In addition, the shape of the ventral process of the squamosal and thus of the seam, can be affected by the cross-section of the lower temporal bar. The joint essentially involves the posterior process of the squamosal expanding medially around the dorsal edge of the jugal but not necessarily to the extent that it maintains contact with the quadratojugal at its anteriormost point (Figure 64.1). This arrangement resists dorsal and medial movements of the jugal. In DGPC1 the medial surface of the facet bears anteroventrally directed grooves while the dorsal surface is rugose. In small hatchling skulls (< 9 mm long), the jugal does not meet the squamosal (Howes and Swinnerton 1901;
Werner 1962).
Postorbital-squamosal
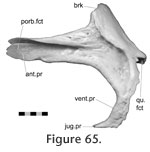 In general, the posterior process of the postorbital overlaps the wide anterior process of the squamosal in a joint that spans almost the full length of upper temporal bar (Figure 60,
Figure 63,
Figure 64.1,
Figure 65,
Figure 66). In medial view the anterior process of the squamosal can be seen to reach the postorbital bar. The tip of the thin squamosal anterior process ends in two to four triangular points (two in LDUCZ x723, YPM 11419, LDUCZ x343; three in In general, the posterior process of the postorbital overlaps the wide anterior process of the squamosal in a joint that spans almost the full length of upper temporal bar (Figure 60,
Figure 63,
Figure 64.1,
Figure 65,
Figure 66). In medial view the anterior process of the squamosal can be seen to reach the postorbital bar. The tip of the thin squamosal anterior process ends in two to four triangular points (two in LDUCZ x723, YPM 11419, LDUCZ x343; three in
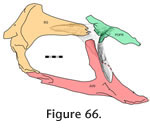 DGPC1, LDUCZ x036; four in LDUCZ x1176) which lie in a depression on the postorbital (Figure
63). Posteriorly, the tip of the postorbital sits against a recess in the squamosal bone (Figure 2.13). The extent of the recess varies; being very shallow in some specimens, e.g., LDUCZ x1176 (Figure 64.1); or deep with edges that envelop the posteriorly tapering postorbital, e.g., DGPC1 (Figure 65). Primarily this joint would substantially restrict lateral movement of the squamosal and medial movement of the postorbital. However, when the facet of the squamosal is deeper it would also restrict anterior or dorsoventral movement of the squamosal and posterior or dorsoventral movement of the postorbital. DGPC1, LDUCZ x036; four in LDUCZ x1176) which lie in a depression on the postorbital (Figure
63). Posteriorly, the tip of the postorbital sits against a recess in the squamosal bone (Figure 2.13). The extent of the recess varies; being very shallow in some specimens, e.g., LDUCZ x1176 (Figure 64.1); or deep with edges that envelop the posteriorly tapering postorbital, e.g., DGPC1 (Figure 65). Primarily this joint would substantially restrict lateral movement of the squamosal and medial movement of the postorbital. However, when the facet of the squamosal is deeper it would also restrict anterior or dorsoventral movement of the squamosal and posterior or dorsoventral movement of the postorbital.
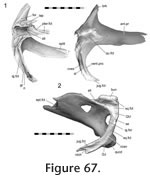 Squamosal-quadrate/quadratojugal Squamosal-quadrate/quadratojugal
In posterolateral view, this slightly convoluted seam extends posterodorsally from the junction with the jugal, with only the posterior portion curving over the dorsal tip of the quadrate. In posterior view the seam can be seen to continue anteromedially before disappearing at the junction with the pterygoid. In medial view, the seam travels posteriorly from the junction with the jugal, curves over the quadratojugal foramen and continues dorsally before turning anteromedially to the junction with the pterygoid.
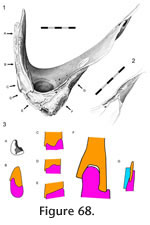 The squamosal articulates with the head of the quadrate in a seemingly firm slotted joint (Figure 64,
Figure 67,
Figure 68,
Figure 69, and
Figure 70) that can be divided into three parts: a lateral part, a small posterior part (above the quadratojugal foramen) and large posteromedial part. In the lateral portion the ventral process of the squamosal sits on the dorsal edge of the quadratojugal but the lateral part of the interface alters along its length. Anteriorly, where this joint is associated with the jugal, the quadratojugal is 'L' shaped in cross-section with a dorsally expanded medial edge ('A' in
Figure 68.3). This medial edge is enveloped by the squamosal which bears a number of anteroventrally directed ridges ('B' in
Figure 68.3). The opposing facet on the quadratojugal is rough but matching grooves are not obvious (Figure 64). More posteriorly, near the posterior part of the quadratojugal-jugal joint, the lateral edge of the quadratojugal expands dorsally (Figure 64.2). The squamosal articulates with the head of the quadrate in a seemingly firm slotted joint (Figure 64,
Figure 67,
Figure 68,
Figure 69, and
Figure 70) that can be divided into three parts: a lateral part, a small posterior part (above the quadratojugal foramen) and large posteromedial part. In the lateral portion the ventral process of the squamosal sits on the dorsal edge of the quadratojugal but the lateral part of the interface alters along its length. Anteriorly, where this joint is associated with the jugal, the quadratojugal is 'L' shaped in cross-section with a dorsally expanded medial edge ('A' in
Figure 68.3). This medial edge is enveloped by the squamosal which bears a number of anteroventrally directed ridges ('B' in
Figure 68.3). The opposing facet on the quadratojugal is rough but matching grooves are not obvious (Figure 64). More posteriorly, near the posterior part of the quadratojugal-jugal joint, the lateral edge of the quadratojugal expands dorsally (Figure 64.2).
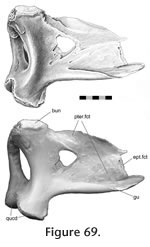 At this point the dorsal surface of the quadratojugal is concave (Figure
67). Correspondingly, the squamosal surface exhibits a wide convex ridge and two grooves to accept the surface (Figure 67, 'C' in
Figure 68.3). In the posterior part of the squamosal-quadratojugal joint both bones widen, and the facets are rugose but the interface between the two bones is more planar and abutting ('D' and 'E' in
Figure 68.3). At this point the dorsal surface of the quadratojugal is concave (Figure
67). Correspondingly, the squamosal surface exhibits a wide convex ridge and two grooves to accept the surface (Figure 67, 'C' in
Figure 68.3). In the posterior part of the squamosal-quadratojugal joint both bones widen, and the facets are rugose but the interface between the two bones is more planar and abutting ('D' and 'E' in
Figure 68.3).
The posteromedial part of the joint is associated with the pterygoid-squamosal joint, and consists of two slot joints ('F' in
Figure 68.3). The posteriormost and largest of these slot joints involves the dorsal tip of the quadrate which is an expanded bun-like process with an anteromedially directed long axis (Figure 64,
Figure 68,
Figure 69). This 'bun' is held in a large concavity in the squamosal
(Figure 68) between two ventral extensions (Howes and Swinnerton 1901): a robust posteromedial process (or lappet) which extends ventrally from the dorsal process and a longer thinner process which extends anteromedially from the squamosal ventral process and is associated with the squamosa-pterygoid joint (Figure 69.2). The second slot joint involves the thinner process and extends anteriorly into a slightly sigmoid groove on the crest of the quadrate wing (Figure 68.2). The groove itself is contiguous with the rough posterior portion of the quadrate-squamosal joint. Posteriorly, the medial border is strongest (consisting of the quadrate dorsal process) but anteriorly the lateral border is strongest. Anteriorly the base of this groove is perforated by foramina. In both the lateral and posterior parts a short curtain of bone, extending ventrally from the medial edge of the squamosal ventral process, overlaps the dorsomedial edge of the quadrate/quadratojugal. This arrangement increases the contact between the bones. The ventral extension is continuous with the anteromedial squamosal process that slots into the quadrate posteromedially. The joint would prevent significant movement between the two bones in all directions except either dorsal movement of the squamosal or ventral movement of the quadrate.
The expanded bun-like process of the quadrate is spongy and "unfinished" surface suggesting a cartilage component to the joint or small synovial cavity ('F' in
Figure 68.3) (Jones 2006).
Holliday and Witmer (2008) considered a synovial joint to be present here in Sphenodon but did not fully explain the basis for this inference.
The ventral process of the squamosal in hatchling Sphenodon is comparable to that of adults, bearing medial and lateral extensions to hold the quadratojugal. There also seems to be a pocket for the head of the quadrate (Rieppel 1992, figure 2). However, there is no apparent posteromedial lappet as found in juvenile specimens (LDUCZ x1176).
Pterygoid-squamosal
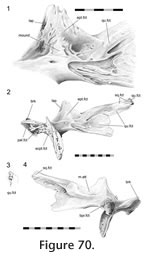 This joint is related to the squamosal-quadrate/quadratojugal joint and the posterior part of the pterygoid-quadrate joint. The posterodorsal processes of the pterygoid wing (Figure 70.3) slot into a pocket formed by the anterior ends of the squamosal posteromedial and anteromedial processes (Figure
67, Figure 68,
Figure 69). The tip of the posterodorsal process of the pterygoid wing is roughly rectangular in cross-section but bears longitudinal grooves and ridges along its surface. The facet on the squamosal also bears some texture. The pocket would resist mediolateral movements between the two bones. This joint is related to the squamosal-quadrate/quadratojugal joint and the posterior part of the pterygoid-quadrate joint. The posterodorsal processes of the pterygoid wing (Figure 70.3) slot into a pocket formed by the anterior ends of the squamosal posteromedial and anteromedial processes (Figure
67, Figure 68,
Figure 69). The tip of the posterodorsal process of the pterygoid wing is roughly rectangular in cross-section but bears longitudinal grooves and ridges along its surface. The facet on the squamosal also bears some texture. The pocket would resist mediolateral movements between the two bones.
Pterygoid-quadrate
As previously observed (e.g.,
Robinson 1973), the quadrate of Sphenodon is firmly joined to the pterygoid. Primarily, the joint involves the lateral surface of the pterygoid posterolateral process and the medial surface of the quadrate anteromedial process (Figure 69,
Figure 70,
Figure 71,
Figure 72).
 However, there are two features of note. Firstly, the ventral edge of the pterygoid
wing slots into a gutter that runs laterally along the base of the quadrate wing
(Figure 69,
Figure 72); secondly, the posterodorsal tip of the pterygoid wing abuts the main body of the quadrate and is related to the squamosal-pterygoid joint (Figure 67, 'G' in
Figure 68.3). In posterior view the seam has a long sigmoid shape (Jones et al. 2009). From the base of the quadrate-pterygoid wing it passes posterodorsally, turns anterodorsally and finally turns posterodorsally again before terminating at a junction with the squamosal. In anteromedial view the short seam runs anteriorly, dorsally and then anteriorly again, before reaching the base of the epipterygoid. However, there are two features of note. Firstly, the ventral edge of the pterygoid
wing slots into a gutter that runs laterally along the base of the quadrate wing
(Figure 69,
Figure 72); secondly, the posterodorsal tip of the pterygoid wing abuts the main body of the quadrate and is related to the squamosal-pterygoid joint (Figure 67, 'G' in
Figure 68.3). In posterior view the seam has a long sigmoid shape (Jones et al. 2009). From the base of the quadrate-pterygoid wing it passes posterodorsally, turns anterodorsally and finally turns posterodorsally again before terminating at a junction with the squamosal. In anteromedial view the short seam runs anteriorly, dorsally and then anteriorly again, before reaching the base of the epipterygoid.
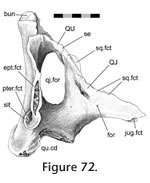 Quadrate-quadratojugal Quadrate-quadratojugal
In Sphenodon this connection is fused ventrally but a faint seam may be visible dorsolaterally above the quadratojugal foramen (e.g., DGPC1, LDUCZ x036) (Figure 72). In hatchlings the bones are not fused (Howes and Swinnerton 1901;
Rieppel, 1992).
Metakinetic joints
The metakinetic joints include those located between the back of the skull and the braincase. The metakinetic unit is linked to the roofing unit by the parietal and squamosal, and to the palatal, rostral and temporal units by the pterygoid. The usage of metakinetic to describe this unit should not be taken to imply that metakinetic movements occur in this taxon.
Epipterygoid-quadrate
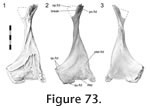 Robinson (1973) incorrectly thought the epipterygoid of Sphenodon overlapped the quadrate. In actuality, the posterior end of the epipterygoid base sits against the pterygoid abutting the anterior edge of the quadrate (Figure 70,
Figure 71,
Figure 72,
Figure 73;
Appendix 2). From the junction with the pterygoid base of the quadrate-pterygoid wing, in anterolateral view, the seam travels anterodorsally, posteriorly, dorsally and finally anterodorsally again. The two anterior processes from the quadrate (dorsal and ventral) are thin but rigid, and the edge of the quadrate between them is expanded and lacks a finished anterior surface. The corresponding posterior end of the epipterygoid also lacks a surface. The dorsally orientated part of the seam (between the unfinished ends of the epipterygoid and quadrate) is sometimes very wide and contains substantial space for soft tissue (e.g., DGPC 2, LDUCZ x1176). In AIM LH833 and the left side of LDUCZ x036 the seam is not easily discernable which suggests that the bones may have begun to fuse. This occurrence is perhaps not unexpected as both are endochondral bones and are derived from the same palatoquadrate cartilage (Howes and Swinnerton 1901;
Schauinsland 1903;
Werner 1962;
Rieppel 1992;
Evans 2008). Robinson (1973) incorrectly thought the epipterygoid of Sphenodon overlapped the quadrate. In actuality, the posterior end of the epipterygoid base sits against the pterygoid abutting the anterior edge of the quadrate (Figure 70,
Figure 71,
Figure 72,
Figure 73;
Appendix 2). From the junction with the pterygoid base of the quadrate-pterygoid wing, in anterolateral view, the seam travels anterodorsally, posteriorly, dorsally and finally anterodorsally again. The two anterior processes from the quadrate (dorsal and ventral) are thin but rigid, and the edge of the quadrate between them is expanded and lacks a finished anterior surface. The corresponding posterior end of the epipterygoid also lacks a surface. The dorsally orientated part of the seam (between the unfinished ends of the epipterygoid and quadrate) is sometimes very wide and contains substantial space for soft tissue (e.g., DGPC 2, LDUCZ x1176). In AIM LH833 and the left side of LDUCZ x036 the seam is not easily discernable which suggests that the bones may have begun to fuse. This occurrence is perhaps not unexpected as both are endochondral bones and are derived from the same palatoquadrate cartilage (Howes and Swinnerton 1901;
Schauinsland 1903;
Werner 1962;
Rieppel 1992;
Evans 2008).
Pterygoid-basisphenoid [sphenoid]
This joint may be referred to as the basipterygoid articulation (e.g.,
Evans 2003;
Johnston 2010) or basal joint (Holliday and Witmer 2008). In squamates this joint is synovial, permitting mobility associated with metakinesis (Evans 1980,
2003).
Günther (1867, p. 599) referred to this joint in Sphenodon as "immoveable" whereas
Romer (1956, p. 114) refers to it as "moveable". Neither author illustrated the articulation or described it further. More recently,
Arnold (1998, p. 325) considered this joint to be mobile in juveniles. Evidence for a synovial joint and interarticular cartilage has also been reported for Sphenodon based on examination of hatchling (Howes and Swinnerton
1901) and adult material (Holiday and Witmer 2008;
Johnston 2010).
Several dry specimens and CT data sets of YPM 9194 and LDUCZ x036 demonstrate that the ventral tips of the basisphenoid rest in pockets on the medial surface of the pterygoids. The pockets are enclosed ventrally, anteriorly and medially but not posteriorly or dorsally (Figure 38.3,
Figure 39.2,
Figure 70.4). In LDUCZ x1176 the pockets are comparably enclosed. However, in hatchlings the medial boundary is almost absent (CM 30660,
Maisano 2001).
Pterygoid-epipterygoid
In Sphenodon the epipterygoid sits against the lateral surface of the pterygoid
wing (Figure 70,
Figure 71). Dorsal expansion of the pterygoid increases the surface area between the two bones and, at least in DGPC1, the dorsal edge bears three triangular processes (Figure 70.1). The facet on the epipterygoid is inset (Figure 73) and within it there is a depression that corresponds to a mound on the pterygoid (Figure 70.1). The anterior edge of the pterygoid bears a subtle lappet which folds around the anterior edge of the epipterygoid (Figure 70.1). Both facets possess additional texture on their surface. The joint would have physically prevented medial movements of epipterygoid and resisted ventral, anterior and posterior movements.
Parietal-supraoccipital
In mammals, this joint is sometimes referred to as the lamboid suture (e.g.,
Opperman 2000). In Sphenodon (e.g., DGPC 2, LDUCZ x036) the lateral edges of the parietal extend ventrolaterally (Figure 59) overlapping the dorsal surface of the supraoccipital, but there is otherwise no distinct facet. The joint would have prevented the supraoccipital from moving dorsally and the parietal from moving ventrally. It would also reduce mediolateral movement between the paired bones.
Squamosal-opisthotic
At its tip, the anterolateral surface of the paroccipital process (opisthotic) rests against the posterior corner of the squamosal (e.g., DGPC2, LDUCZ x036). The joint appears to be fibrous.
Epipterygoid-prootic
The posteromedial surface of the dorsal process of the epipterygoid overlaps the anterior edge of the prootic. The joint appears to involve fibrous tissue (e.g., LDUCZ x036).
|