

 Conodont biostratigraphy, paleoecology, and taphonomy of the Second Creek Bed and Wolcott Furnace Hematite (Clinton Group) in West Central New York State
Conodont biostratigraphy, paleoecology, and taphonomy of the Second Creek Bed and Wolcott Furnace Hematite (Clinton Group) in West Central New York State
Article number: 17.1.10A
https://doi.org/10.26879/388
Copyright Palaeontological Association, February 2014
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 22 March 2013. Acceptance: 5 February 2014
{flike id=679}
ABSTRACT
The Second Creek Bed is a thin, yet widely traceable, conglomeratic stratum at the base of the Williamson Formation in west-central New York and southern Ontario. Correlation of this unit is problematic, and earlier biostratigraphic studies of conodonts and graptolites have yielded contradictory ages. To address these issues, bulk whole-rock samples of the Second Creek Bed and underlying strata were collected from the type locality in west-central New York and processed for microfossil extraction.
A sparse conodont assemblage, tentatively assigned to the Pterospathodus eopennatus Superzone, was recovered from the Wolcott Furnace Hematite, which directly underlies the Second Creek Bed at this locality. The remains of a diverse early to middle Telychian conodont fauna was extracted from the Second Creek Bed. This assemblage is assigned to the Pterospathodus amorphognathoides angulatus Zone although many older, reworked conodont elements are present as well. The findings are consistent with previous chronostratigraphic studies conducted to the west.
Processed samples of the Second Creek Bed also yield small (500-1500 μm), pyritized internal molds of gastropods, bivalves, and tentaculitids in great abundance. We argue that these internal molds were produced during early diagenesis within anoxic mud substrates during the initial deposition of the bed and subsequently reworked and concentrated on the seafloor. However, the high concentration of pyrite and organic matter in this succession also suggests that anoxic, sulfidic conditions prevailed even as erosion was taking place. Therefore we propose that the Second Creek Bed was formed as a result of submarine erosion and reworking of remnant clasts in deep environments that were instigated by density currents, storm waves, and/or internal waves.
Nicholas B. Sullivan. Department of Geology, University of Cincinnati, Cincinnati, Ohio 45221, USA. sullivnb@mail.uc.edu
Mark A. Kleffner. Division of Geological Sciences, School of Earth Sciences, The Ohio State University at Lima, Lima, Ohio 45804, USA. kleffner.1@osu.edu
Carlton E. Brett. Department of Geology, University of Cincinnati, Cincinnati, Ohio 45221-0013, USA. brettce@ucmail.uc.edu
Keywords: Silurian; Conodont; Marine Authigenesis; Anoxia; Sequence Stratigraphy; Clinton Group
Final citation: Sullivan, Nicholas B., Kleffner, Mark A., and Brett, Carlton E. 2014. Conodont biostratigraphy, paleoecology, and taphonomy of the Second Creek Bed and Wolcott Furnace Hematite (Clinton Group) in West Central New York State. Palaeontologia Electronica Vol. 17, Issue 1;10A; 18p. https://doi.org/10.26879/388
palaeo-electronica.org/content/2014/679-conodonts-in-second-creek-bed
INTRODUCTION
Geologic Setting
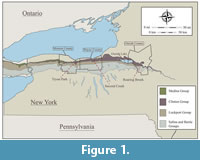 The Second Creek Bed is a thin conglomeratic stratum that can be traced along the Silurian outcrop belt between Oneida Lake and the Bruce Peninsula in Ontario (Brett et al., 1995). Although the bed was first recognized and described by Gillette (1947) it was formally named and defined by Lin and Brett (1988), who established the type section at Second Creek, a small tributary of Sodus Bay in Wayne County, New York (Figure 1). Here, the Second Creek Bed is 2-4 centimeters thick, and it contains abundant limestone rip-up clasts and rounded quartz and phosphate grains (Figure 2; Lin and Brett, 1988). Pyrite is ubiquitous, occurring frequently as coatings and sheaths around fossils or small, rounded phosphate or quartz grains (Figure 2.1-3). The unit also contains fairly abundant (often partially phosphatized) brachiopods, particularly the genus Eoplectodonta transversalis (Lin and Brett, 1988).
The Second Creek Bed is a thin conglomeratic stratum that can be traced along the Silurian outcrop belt between Oneida Lake and the Bruce Peninsula in Ontario (Brett et al., 1995). Although the bed was first recognized and described by Gillette (1947) it was formally named and defined by Lin and Brett (1988), who established the type section at Second Creek, a small tributary of Sodus Bay in Wayne County, New York (Figure 1). Here, the Second Creek Bed is 2-4 centimeters thick, and it contains abundant limestone rip-up clasts and rounded quartz and phosphate grains (Figure 2; Lin and Brett, 1988). Pyrite is ubiquitous, occurring frequently as coatings and sheaths around fossils or small, rounded phosphate or quartz grains (Figure 2.1-3). The unit also contains fairly abundant (often partially phosphatized) brachiopods, particularly the genus Eoplectodonta transversalis (Lin and Brett, 1988).
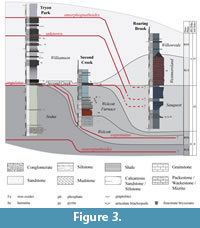
 The Second Creek Bed constitutes the basal portion of the Williamson Formation, the lower part of which is characterized by several centimeters of fissile, organic-rich black shales that yield abundant graptolites (Lin and Brett, 1988; Loydell et al., 2007). These beds overlie progressively older strata to the west; at its type locality, the Second Creek Bed overlies the Wolcott Furnace Hematite, at Rochester it overlies the Lower Sodus Shale (Figure 3; Brett et al., 1990, 1998).
The Second Creek Bed constitutes the basal portion of the Williamson Formation, the lower part of which is characterized by several centimeters of fissile, organic-rich black shales that yield abundant graptolites (Lin and Brett, 1988; Loydell et al., 2007). These beds overlie progressively older strata to the west; at its type locality, the Second Creek Bed overlies the Wolcott Furnace Hematite, at Rochester it overlies the Lower Sodus Shale (Figure 3; Brett et al., 1990, 1998).
These observations indicate that the bed overlies a regionally angular unconformity (Brett et al., 1998), and the co-occurrence of corroded limestone clasts and quartz pebbles (Figure 2) suggests that the bed incorporates the remnants of an erosional lag. However, the abundance of pyrite and phosphate clasts within the Second Creek Bed, as well as its close association with the black graptolitic shale facies of the lower Williamson, has led to the conclusion that it was deposited in a relatively deep (tens of meters), distal environment where dysoxic and highly sulfidic conditions prevailed during its final deposition (Eckert and Brett, 1989; Brett et al., 1990, 1998).
 This apparently contradictory set of observations is not unprecedented. In their studies of Middle and Upper Devonian successions elsewhere in New York State, Baird and Brett (1986, 1991) proposed a genetic model explaining the origin of pyritic lag beds in successions that comprise predominantly unfossiliferous (and presumably deep-water) black and dark gray shales. They argued that, during times of low sedimentation, lag deposits were formed by episodic erosion and scouring by deep storm waves, density currents, and internal waves produced at the interface between water masses of different densities (Baird and Brett, 1986, 1991). As evidence of this, Baird and Brett (1986, 1991) cite horizons of concentrated, reworked pyrite and phosphate debris overlain by (and interbedded with) black shales, as well as outcrop-scale scour features in similar facies. Given that pyrite is highly unstable in oxidizing conditions, they conclude that anoxic to dysoxic conditions prevailed throughout deposition, leading to preferential preservation of pyritic and phosphatic sediment at the expense of carbonate, which would have been susceptible to dissolution in these reducing conditions (Baird and Brett, 1991; Boggs, 2001).
This apparently contradictory set of observations is not unprecedented. In their studies of Middle and Upper Devonian successions elsewhere in New York State, Baird and Brett (1986, 1991) proposed a genetic model explaining the origin of pyritic lag beds in successions that comprise predominantly unfossiliferous (and presumably deep-water) black and dark gray shales. They argued that, during times of low sedimentation, lag deposits were formed by episodic erosion and scouring by deep storm waves, density currents, and internal waves produced at the interface between water masses of different densities (Baird and Brett, 1986, 1991). As evidence of this, Baird and Brett (1986, 1991) cite horizons of concentrated, reworked pyrite and phosphate debris overlain by (and interbedded with) black shales, as well as outcrop-scale scour features in similar facies. Given that pyrite is highly unstable in oxidizing conditions, they conclude that anoxic to dysoxic conditions prevailed throughout deposition, leading to preferential preservation of pyritic and phosphatic sediment at the expense of carbonate, which would have been susceptible to dissolution in these reducing conditions (Baird and Brett, 1991; Boggs, 2001).
Age and Correlation
Correlation of the Second Creek Bed is problematic, particularly to the east of Oneida Lake (see Figure 1). Previous authors have argued for its equivalence with the Westmoreland Hematite, a sparsely fossiliferous oolitic ironstone that occurs between the underlying Sauquoit and overlying Willowvale Formations in Oneida County (Figure 1, Figure 3; Lin and Brett, 1988; Brett et al., 1990, 1998). This correlation is implied by Gillette (1947) on the basis of macrofossil assemblages, an ostracode based biostratigraphy, and phosphatic-pyritic beds with hematitic stringers overlying the Sauquoit Formation in cores drilled from bedrock between Second Creek and Oneida Lake (these cores are described in detail by Gillette (1947) but, unfortunately, they are no longer accounted for). Lin and Brett (1988) suggested that these pyritic, hematitic beds overlying the Sauquoit Formation may represent transitional facies between the Second Creek Bed and Westmoreland Hematite.
This presumed equivalence of these units has been an important keystone for many paleoecological and stratigraphic studies (e.g., Eckert and Brett, 1989; Brett et al., 1990, 1998; McLaughlin et al., 2008a). Brett et al. (1990) interpreted these units as the basal transgressive lag of S-IV, the fourth 3rd-order sequence of the classic Niagaran Series in the northern Appalachian Basin and surrounding depositional provinces (Brett et al., 1998; Brett and Ray, 2005; McLaughlin et al., 2008b; Cramer, 2009).
However, chronostratigraphic data on these problematic units is limited. Rexroad and Rickard (published as Rexroad and Richard, 1965, p. 1219) make a passing reference to conodonts from the late Telychian Pterospathodus amorphognathoides Zone recovered in the underlying Sauquoit Formation, although the reasoning behind this claim was not discussed. Conodonts of this Zone were also recovered by Kleffner (reported via Brett et al., 1998) in the Westmoreland Hematite and overlying Willowvale Shale, with possible admixture of older Pterospathodus celloni Zone conodonts.
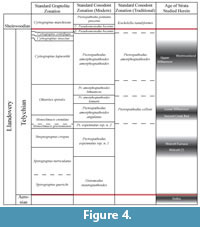 No conodonts from the Second Creek Bed were reported or figured in publication until a 2007 report by Loydell and others. By this time, the Telychian conodont zonation had undergone several revisions and refinements (Figure 4; Jeppsson, 1997; Männik, 1998, 2007a). The Pterospathodus amorphognathoides Zone of Walliser (1964) was split by Jeppsson (1997) into three Zones; the lowest of these retained the original name, while the upper two were reassigned to the Lower and Upper Pseudooneotodus bicornis Zones (Figure 4). However, a decade later Männik (2007a), employing a refined taxonomy for the conodont genus Pterospathodus (Männik, 1998), redefined the Pterospathodus amorphognathoides Zone using the zonally significant subspecies Pterospathodus amorphognathoides amorphognathoides.
No conodonts from the Second Creek Bed were reported or figured in publication until a 2007 report by Loydell and others. By this time, the Telychian conodont zonation had undergone several revisions and refinements (Figure 4; Jeppsson, 1997; Männik, 1998, 2007a). The Pterospathodus amorphognathoides Zone of Walliser (1964) was split by Jeppsson (1997) into three Zones; the lowest of these retained the original name, while the upper two were reassigned to the Lower and Upper Pseudooneotodus bicornis Zones (Figure 4). However, a decade later Männik (2007a), employing a refined taxonomy for the conodont genus Pterospathodus (Männik, 1998), redefined the Pterospathodus amorphognathoides Zone using the zonally significant subspecies Pterospathodus amorphognathoides amorphognathoides.
The original Pterospathodus celloni Zone of Walliser (1964) was elevated by Männik (2007a) to the rank of Superzone, and split into the Pterospathodus amorphognathoides angulatus, Pterospathodus amorphognathoides lennarti, and Pterospathodus amorphognathoides lithuanicus Zones, in ascending order (Figure 4). Two additional Zones, the Pterospathodus eopennatus ssp. n. 1 and ssp. n. 2 Zones (of Männik, 1998) constitute the early Telychian, below the Pterospathodus celloni Superzone (Figure 4; Männik, 2007a). Straddling the base of the Telychian is the Distomodus staurognathoides Zone, which extends downward into the Aeronian Stage (Männik, 2007b)
Loydell et al. (2007) reported a diverse conodont assemblage from the Second Creek Bed at Tryon Park (Rochester, New York; Figure 1), which was assigned to the Pterospathodus amorphognathoides angulatus Zone (Figure 3). This diagnosis, which corresponds to the early part of the Pterospathodus celloni Zone of previous terminology, suggests that the Second Creek Bed and the lower part of the Williamson Shale are appreciably older than the Westmoreland Hematite and the Lower Willowvale Shale, which were reassigned to the Pterospathodus amorphognathoides amorphognathoides Zone in that same study (Figure 3, Figure 4; Loydell et al., 2007). Conodonts of the Pterospathodus amorphognathoides amorphognathoides Zone were found in the Williamson Shale, but only in strata 1.5-4.5 meters above its base (Figure 3; note that half a meter has been added to the original measurements of Loydell et al. (2007) due to a misplacement of the base of the Williamson; see Verniers et al., 2012). The younger age of the Westmoreland Hematite has also been corroborated by regional studies of stable carbon isotope (δ13Ccarb) analyses which show steadily rising values in the upper Sauquoit-Westmoreland-Willowvale succession, which are interpreted as the rising limb of the upper Llandovery-lower Wenlock "Ireviken" δ13Ccarb Excursion (McLaughlin et al., 2012).
Loydell et al. (2007) also identified the graptolite Stimulograptus clintonensis occurring throughout the lower 2.5 meters of the Williamson, ranging upward into beds that had been assigned to the Pterospathodus amorphognathoides amorphognathoides conodont Zone. This was considered problematic, since this biostratigraphically important graptolite had never been recorded above the Monograptus griestoniensis Zone (Loydell et al., 2007). This previously established upper limit of the range of Stimulograptus clintonensis corresponds to the upper part of the Pterospathodus eopennatus ssp. n. 2 conodont Zone, indicating a significantly older age for these beds than what is suggested by conodont data (Figure 4; Loydell et al., 2007; Männik, 2007a).
This "enigma" was addressed in a more recent study of chitinozoans and conodonts collected from the succession at Tryon Park made by Verniers et al. (2012). Although they note that the Second Creek Phosphate Bed is mislabeled on figure 2 of Loydell et al. (2007; it is actually 0.5 meters lower in the section than they show), they corroborate a Pterospathodus amorphognathoides angulatus assignment for the bed with additional chitinozoan biostratigraphy (Verniers et al., 2012).
Statement of the Problem
Nearly all the published biostratigraphic data on the Second Creek Bed come from Tryon Park in Rochester New York (e.g., Loydell et al., 2007; Verniers et al., 2012). However, different biostratigraphic indicators collected from this locality suggest contradictory ages (Loydell et al., 2007). These ages in turn, are significantly older than the accepted age of the bed's presumed lateral equivalent: the Westmoreland Hematite (Lin and Brett, 1988; Brett et al., 1998; Loydell et al., 2007). Furthermore, the sedimentological characteristics also display an unusual combination of features: the bed has many aspects of a transgressive lag bed overlying an erosive contact, yet the abundant pyrite and phosphate clasts and the association with graptolitic black shales suggests that anoxic (and presumably deep water) conditions prevailed during its deposition (Lin and Brett, 1988).
These unresolved issues have implications for stratigraphic frameworks at local, regional, and global scales. The chronostratigraphic control provided here will necessitate a series of refinements to the sequence stratigraphic framework for the well-studied sections of New York (Brett et al., 1990). This in turn will have a significant impact on sequence stratigraphic interpretations throughout eastern North America, particularly those that employ the New York successions as a reference standard (i.e., Brett et al., 1998; Chowns, 2006; McLaughlin et al., 2008b, 2012; Cramer, 2009). A refined depositional history may also lead to a better understanding of the depositional processes behind the genesis of certain unusual and economically valuable rock types that are common throughout these intervals. Finally, the data presented here may elucidate the local manifestation of a globally recognized biotic crisis which occurred just prior to the deposition of the Second Creek Bed. This is the Valgu Event, which is recorded within the lower Telychian Pterospathodus eopennatus Superzone (Männik, 2005, 2007a; Munnecke and Männik, 2009).
MATERIALS AND METHODS
Two localities were measured and sampled over the course of this study. The first is an interval of 1.3 meters of strata exposed in the bed and banks of Second Creek (i.e., Section 14 of Gillette, 1947, p. 142-146). These outcrops are in Alton (Wayne County), New York, approximately 0.40 kilometers from the intersection between Red Mill Road, Shaker Tract Road, and Shaker Road, along a heading 116.9° west of north (precise coordinates: N 43.233322°; E -76.976819°). Several kilograms of sample were collected; stratigraphic height and orientation were noted. Approximately 4 kilograms of the Second Creek Bed were collected; an additional 1.3 kilograms of a limestone bed was also collected from the underlying Wolcott Furnace Hematite.
The second locality is a small, south-flowing tributary of Oriskany Creek that serves as the type locality of the Westmoreland Hematite (Gillette, 1947, p. 92-93; see also Brett and Goodman, 1996, Stop 13, p. 221-223). Locally referred to as "Roaring Brook," this outcrop is located just south of New York State Route 5 (Seneca Turnpike), approximately 0.81 kilometers due east of the intersection of County Road 15A and New York State Route 5 in Lairdsville, NY (precise coordinates: N 43.080083°; E -75.421331°). Sampled beds included a thin, rippled conglomeratic bed near the top of the Sauquoit, a dolomitic sandstone horizon immediately below the Westmoreland, and samples of the ironstone itself (Figure 3).
Whole-rock samples were then processed for conodont microfossil extraction using a buffered formic acid solution following a procedure modified from the recommendations of Jeppsson and Anehus (1995). Every 650 grams of sample was dissolved using a solution containing 8 liters of water, 150 grams of powdered CaCO3, 10 grams of powdered CaPO4, and 750 milliliters of 85% formic acid (mixed together in that order).
The dissolved residue was washed and sorted by grain size using sieves (U.S. Standard Sieve Mesh # 10, 40, 140, and 230). The residues collected on Mesh #140 were examined under a standard light microscope and microfossils were collected and many were identified. The Wolcott Furnace sample produced abundant residue, but few conodont elements. To expedite the process of conodont collection, this residue was subjected to heavy liquid separation, using LST (a solution of lithium polytungstate and water).
Several specimens that held biostratigraphic or paleoecological significance were sputtered with a Gadolinium-Platinum alloy and imaged on a scanning electron microscope maintained by the College of Engineering at the University of Cincinnati. All collected microfossil specimens and residues are now in storage at the School of Earth Sciences at the Ohio State University Campus at Lima in the collections of Mark A. Kleffner. Slabs of the larger samples were cut, polished, and scanned at high resolution for general sedimentological analyses. These are currently held in the storage at the Wisconsin Geological and Natural History Survey.
RESULTS
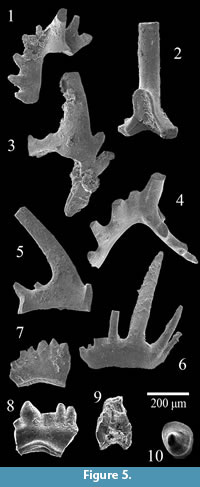 A single sample of the Wolcott Furnace Hematite collected from Second Creek, weighing 650 grams, was dissolved for microfossil extraction. This bed was a thin (2-3 cm) bluish grey limestone collected from 35 centimeters above the base of the Wolcott Furnace, 38 centimeters below the Second Creek Bed. The limestone horizon is a skeletal packstone composed primarily of small fenestrate bryozoans, though it contains ribbed strophomenid brachiopods as well. The sample yielded several dozen conodont elements, which were light brown in color (CAI 2). Although many of the elements are fragmented, several are sufficiently intact for tentative identification. The conodont assemblage recovered was predominantly composed of coniform elements such as Panderodus but several ramiform elements and a single pectiniform element were recovered as well (Figure 5, Figure 6). However, most of these were too fragmented to make definitive identifications. Specimens 1 and 2 on Figure 5 have been tentatively recognized as Aspelundia expansa. Specimens 3 and 4 also show affinity toward this taxon, though the latter is also comparable to the Sc element of Oulodus sigmoideus. Specimen 5 can also be assigned to the genus Aspelundia,
A single sample of the Wolcott Furnace Hematite collected from Second Creek, weighing 650 grams, was dissolved for microfossil extraction. This bed was a thin (2-3 cm) bluish grey limestone collected from 35 centimeters above the base of the Wolcott Furnace, 38 centimeters below the Second Creek Bed. The limestone horizon is a skeletal packstone composed primarily of small fenestrate bryozoans, though it contains ribbed strophomenid brachiopods as well. The sample yielded several dozen conodont elements, which were light brown in color (CAI 2). Although many of the elements are fragmented, several are sufficiently intact for tentative identification. The conodont assemblage recovered was predominantly composed of coniform elements such as Panderodus but several ramiform elements and a single pectiniform element were recovered as well (Figure 5, Figure 6). However, most of these were too fragmented to make definitive identifications. Specimens 1 and 2 on Figure 5 have been tentatively recognized as Aspelundia expansa. Specimens 3 and 4 also show affinity toward this taxon, though the latter is also comparable to the Sc element of Oulodus sigmoideus. Specimen 5 can also be assigned to the genus Aspelundia,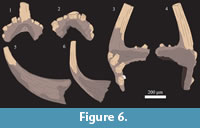 though it is too fragmented to confirm species identification. Specimen 6 could not be confidently identified, though it shows affinities with the Sb element of a species of Oulodus or Aspelundia. Specimens 7 and 8 on Figure 5, though poorly preserved can be assigned to the genus Ozarkodina polinclinata estonica; the Sb element of this taxon is illustrated in Figure 6, as are the Sc element of an Oulodus or Aspelundia species, and two Panderodus unicostatus elements. A small, lyriform element of Apsidognathus tuberculatus ssp. n. 1 of Männik, 2007a was also recovered from the Wolcott Furnace (Figure 5.9).
though it is too fragmented to confirm species identification. Specimen 6 could not be confidently identified, though it shows affinities with the Sb element of a species of Oulodus or Aspelundia. Specimens 7 and 8 on Figure 5, though poorly preserved can be assigned to the genus Ozarkodina polinclinata estonica; the Sb element of this taxon is illustrated in Figure 6, as are the Sc element of an Oulodus or Aspelundia species, and two Panderodus unicostatus elements. A small, lyriform element of Apsidognathus tuberculatus ssp. n. 1 of Männik, 2007a was also recovered from the Wolcott Furnace (Figure 5.9).
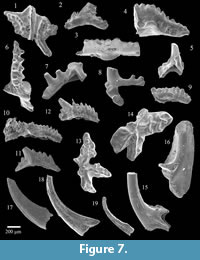 Insoluble residues of the Second Creek Bed yielded several hundred conodont elements, which were also light brown (CAI 2) in color. Several representative taxa of this assemblage have been illustrated in Figure 7 and Figure 8. Many zonally significant taxa can be recognized in this sample, notably Aulacognathus kuehni (Figure 7.1), Ozarkodina polinclinata estonica (Figure 7.2), Ozarkodina gulletensis (Figure 7.3), Ozarkodina sp. n. of Männik, 2007a
Insoluble residues of the Second Creek Bed yielded several hundred conodont elements, which were also light brown (CAI 2) in color. Several representative taxa of this assemblage have been illustrated in Figure 7 and Figure 8. Many zonally significant taxa can be recognized in this sample, notably Aulacognathus kuehni (Figure 7.1), Ozarkodina polinclinata estonica (Figure 7.2), Ozarkodina gulletensis (Figure 7.3), Ozarkodina sp. n. of Männik, 2007a (Figure 7.4, Figure 8.1), and Apsidognathus tuberculatus ssp. n. 2 of Männik, 2007a (Figure 8.2). Several elements of the zonal taxon Pterospathodus amorphognathoides angulatus are also present (Figure 7.10-12, Figure 8.5-6). Although it shares many anatomical features in common with the slightly older Pterospathodus eopennatus ssp. n. 2, Pterospathodus amorphognathoides angulatus can be distinguished by a greater number of denticles and more elongate Pa and Pb elements (see Männik, 1998). The insoluble residues also contained numerous pyritized internal molds of small mollusks (Figure 9). None of these specimens could be assigned definitely to genus or species level. However, bivalves, gastropods, and tentaculitids are all represented.
(Figure 7.4, Figure 8.1), and Apsidognathus tuberculatus ssp. n. 2 of Männik, 2007a (Figure 8.2). Several elements of the zonal taxon Pterospathodus amorphognathoides angulatus are also present (Figure 7.10-12, Figure 8.5-6). Although it shares many anatomical features in common with the slightly older Pterospathodus eopennatus ssp. n. 2, Pterospathodus amorphognathoides angulatus can be distinguished by a greater number of denticles and more elongate Pa and Pb elements (see Männik, 1998). The insoluble residues also contained numerous pyritized internal molds of small mollusks (Figure 9). None of these specimens could be assigned definitely to genus or species level. However, bivalves, gastropods, and tentaculitids are all represented.
 Two samples of the Second Creek Bed were cut, polished, and scanned at high resolution (Figure 2). These images show large rip-up clasts of fine grained carbonate set within a coarse matrix of sand sized black (presumably phosphatic) and iron sulfide minerals. The edges of these rip-up clasts show many pits and cavities that have been filled with grains of phosphate and iron sulfides (Figure 2.1). Large (up to half centimeter) rounded clasts of quartz and phosphate are also present in great abundance, as are shells which can be viewed in cross section (Figure 2.2). Many of the smaller (sand-sized) grains of phosphate and quartz are coated in a thin pyritic rind, which can be seen under higher magnification (Figure 2.2), though in some places, sediment grains composed entirely of iron sulfides are concentrated along surfaces (Figure 2.3).
Two samples of the Second Creek Bed were cut, polished, and scanned at high resolution (Figure 2). These images show large rip-up clasts of fine grained carbonate set within a coarse matrix of sand sized black (presumably phosphatic) and iron sulfide minerals. The edges of these rip-up clasts show many pits and cavities that have been filled with grains of phosphate and iron sulfides (Figure 2.1). Large (up to half centimeter) rounded clasts of quartz and phosphate are also present in great abundance, as are shells which can be viewed in cross section (Figure 2.2). Many of the smaller (sand-sized) grains of phosphate and quartz are coated in a thin pyritic rind, which can be seen under higher magnification (Figure 2.2), though in some places, sediment grains composed entirely of iron sulfides are concentrated along surfaces (Figure 2.3).
No conodonts were recovered from the Westmoreland Hematite at Roaring Brook. A sample of the rippled conglomeratic horizon underlying the Westmoreland Hematite, scanned at high resolution, shows numerous rounded to sub-rounded quartz and phosphate pebbles reaching up to a centimeter in diameter (Figure 10.1-2). The bed contains ubiquitous, sand-sized grains which may display concentric laminae when viewed in cross section (Figure 10.1-3). The mineralogy of these laminae is unknown, but they may vary significantly in color, ranging from light gray, to pink, to dark red. Iron sulfides are also common, frequently occurring as thin rinds on the margins of larger clasts (Figure 10.2).
DISCUSSION
Biostratigraphy
 The conodont assemblage recovered from the Wolcott Furnace Hematite, while of limited biostratigraphic value on its own, can offer some insight into the age of this bed when used in conjunction with other data. The Lower Sodus Shale, which is stratigraphically below the Wolcott and Wolcott Furnace, has been assigned to the Distomodus staurognathoides Zone and an assemblage of chitinozoans collected from the same unit, suggest an age no earlier than the latest Aeronian (Figure 4; Verniers et al., 2012). The only biostratigraphically significant taxa that can be recognized in the Wolcott Furnace assemblages with a reasonable degree of confidence are Ozarkodina polinclinata estonica and Apsidognathus tuberculatus ssp. n. 1 of Männik, 2007a, the co-occurrence of which suggest assignment to the Pterospathodus eopennatus Superzone (Figure 11). Although Aspelundia expansa has not been reported above the top of the Distomodus staurognathoides Zone (Männik, 2007b), the biostratigraphic range of this taxa is comparatively uncertain, and none of the elements can be assigned to this species with a high degree of confidence.
The conodont assemblage recovered from the Wolcott Furnace Hematite, while of limited biostratigraphic value on its own, can offer some insight into the age of this bed when used in conjunction with other data. The Lower Sodus Shale, which is stratigraphically below the Wolcott and Wolcott Furnace, has been assigned to the Distomodus staurognathoides Zone and an assemblage of chitinozoans collected from the same unit, suggest an age no earlier than the latest Aeronian (Figure 4; Verniers et al., 2012). The only biostratigraphically significant taxa that can be recognized in the Wolcott Furnace assemblages with a reasonable degree of confidence are Ozarkodina polinclinata estonica and Apsidognathus tuberculatus ssp. n. 1 of Männik, 2007a, the co-occurrence of which suggest assignment to the Pterospathodus eopennatus Superzone (Figure 11). Although Aspelundia expansa has not been reported above the top of the Distomodus staurognathoides Zone (Männik, 2007b), the biostratigraphic range of this taxa is comparatively uncertain, and none of the elements can be assigned to this species with a high degree of confidence.
 The presence of (presumably reworked) Aulacognathus kuehni and Apsidognathus tuberculatus ssp. n. 2 of Männik (2007a) in the Second Creek Bed suggests it incorporates remnants of strata referable to the Pterospathodus eopennatus Superzone (Männik, 2007a). Although the extensive lateral transport of material cannot be ruled out entirely, it is probable (though not certain) that beds of this interval used to overlie the Wolcott Furnace before their removal during an erosive event that preceded deposition of the Second Creek Bed.
The presence of (presumably reworked) Aulacognathus kuehni and Apsidognathus tuberculatus ssp. n. 2 of Männik (2007a) in the Second Creek Bed suggests it incorporates remnants of strata referable to the Pterospathodus eopennatus Superzone (Männik, 2007a). Although the extensive lateral transport of material cannot be ruled out entirely, it is probable (though not certain) that beds of this interval used to overlie the Wolcott Furnace before their removal during an erosive event that preceded deposition of the Second Creek Bed.
These conclusions are also consistent with circumstantial evidence for a Pterospathodus eopennatus Superzone assignment for the Wolcott Furnace. Previous workers have correlated the underlying Wolcott Limestone to the Waco Limestone of eastern Kentucky (Hunter, 1960, 1970), primarily on the basis of traditional stratigraphic methods employing macrofossil assemblages and physical stratigraphy. This correlation is notable because the Waco Limestone contains conodonts from the Pterospathodus eopennatus Superzone (Kleffner, in McLaughlin et al., 2008a), and a significant positive δ13C excursion which has been interpreted as the early Telychian Valgu Excursion, also restricted to the Pterospathodus eopennatus Superzone (Munnecke and Männik, 2009; McLaughlin et al., 2012).
The conodonts recovered from the Second Creek Bed allow for a more definitive statement on the timing of its deposition. The co-occurrence of Ozarkodina polinclinata estonica, Ozarkodina sp. n. of Männik (2007a), Ozarkodina gulletensis and Pterospathodus amorphognathoides angulatus suggests that this assemblage belongs to the Pterospathodus amorphognathoides angulatus Zone (Figure 11; Männik, 2007a). Although Männik (2007a), working primarily in eastern Europe, placed the last appearance datum of Aulacognathus kuehni and Apsidognathus tuberculatus ssp. n. 2 in the Pterospathodus eopennatus ssp. n. 2 Zone (well below the first appearance datum of Pterospathodus amorphognathoides angulatus) the co-occurrence of these taxa in this sample is perhaps unsurprising. In light of the numerous rip-up clasts present in this bed and its position over a regionally angular unconformity, it seems entirely probable that this bed would incorporate a significant portion of reworked older conodont elements.
The conodont assemblage recovered from the Second Creek Bed at its type locality is qualitatively very similar to the assemblage reported basal phosphatic horizon of the Williamson Formation at Tryon Park, in Rochester, New York, which is located approximately 46 kilometers to the west (Figure 1; Loydell et al., 2007). This association corroborates the original correlation of these beds as proposed by Lin and Brett (1988) and demonstrates the relative isochroneity of this stratigraphic contact on a regional scale. Abundant graptolites were observed in the black shales immediately overlying the Second Creek Bed at its type locality, though none are identified here. Therefore, these findings cannot resolve the "biostratigraphical enigma" discussed by Loydell et al. (2007), beyond demonstrating that the basal phosphate horizon of the Williamson at Tryon Park is in fact the Second Creek Bed, and that its assignment to Pterospathodus amorphognathoides angulatus Biozone is a regional characteristic of this unit.
However, the regional isochroneity of the base of the Williamson does cast further doubt on the correlation of the Second Creek Bed and the Westmoreland Hematite (e.g., Gillette, 1947; Lin and Brett, 1988; Eckert and Brett, 1989; Brett et al., 1990, 1998). The insoluble residues recovered from samples collected from Roaring Brook yielded no conodonts; therefore we cannot directly corroborate the Pterospathodus amorphognathoides amorphognathoides age assignment for the Westmoreland and Willowvale (Loydell et al., 2007). Nevertheless, the results from Wayne and Monroe Counties do suggest that the base of the Upper Clinton Group (i.e., the base of S-IV) at Second Creek and Tryon Park may not be the same stratigraphic contact as the one overlain by the Westmoreland Hematite in Oneida County, if we accept the previously reported age for the latter unit (Figure 3; Loydell et al., 2007). In light of this information, it is possible that the Second Creek Bed correlates to an as-of-yet unrecognized horizon within the lower Westmoreland or underlying Sauquoit Formation.
Yet there remains the observation that the Sauquoit Formation thins progressively to the west underneath a thin interval containing abundant pyrite and hematite (Gillette, 1947, p. 13, 181), coupled with the faunal and biostratigraphic demarcation between the Sauquoit and overlying strata (Gillette, 1947, p. 77). A possible solution to these concerns may be elucidated by a detailed microstratigraphic study of the Westmoreland, which can be quite lithologically heterogeneous, containing several discrete ferruginous zones separated by beds of fossiliferous shale that swell the overall thickness of the unit locally to about a meter (Gillette, 1947, p. 93; personal observation). Additionally, the frequent association of these beds with underlying dolomitic sandstones (e.g., Roaring Brook, shown on Figure 3, Figure 10) at some localities may suggest that the "Westmoreland" is actually a lithologically heterogeneous association of genetically linked beds. If this is the case, the Second Creek Bed may not necessarily be a correlative of the entire Westmoreland Iron Ore (sensu stricto), but rather a distal expression of one bed within this greater "Westmoreland" Interval. One plausible candidate for the eastern equivalent of the Second Creek Bed may be the conglomeratic unit underlying the Westmoreland Hematite at Roaring Brook (Figure 3, Figure 10). This is a particularly tempting possibility, given the abundant pyrite and large pebbles of quartz and phosphate found in both beds. However, due to the lack of biostratigraphic control on the Roaring Brook section, this must remain speculative for the time being.
Depositional Environment and Taphonomy
The abundance of rip-up clasts and authigenic minerals observed in the Second Creek Bed has been noted by previous workers who interpret this bed as a product of an episode of low sedimentation (associated with maximum flooding) with a significant fraction of the erosive remnants of the preceding lowstand (e.g., Lin and Brett, 1988; Brett et al., 1990, 1998). These authors have also noted that the abundance of pyrite coated grains and phosphatic debris found in the Second Creek Bed suggests that dysoxic, chemically reducing conditions prevailed as final deposition took place. In this context, the presence of pyritized internal molds of small mollusks (Figure 9) is particularly interesting, given that carbonate minerals would be chemically unstable under these same conditions, and the aragonitic shells of these organisms would have dissolved rapidly if left on the sea-floor for any extended period of time (Baird and Brett, 1991; Boggs, 2001). The limestone rip-up clasts contained in this bed are telling examples of this type of corrosive environment, as they show many pits and holes at their outer margins, a likely product of dissolution (Figure 2). Therefore it seems unlikely that these organisms lived concurrently with the deposition of this bed. In light of this, it seems more probable that these small mollusk shells were buried prior to the deposition of the Second Creek Bed, and that these pyritized internal molds are early diagenetic features which formed in the sediment and were exhumed, reworked, and concentrated on the sea floor at a later time. However, this also suggests that reducing conditions prevailed throughout as erosion was taking place; otherwise the pyrite would have been oxidized and remineralized. Therefore a scenario is required wherein dysoxic conditions could be maintained while erosion and reworking of sediments on the sea floor could occur.
One possibility is that the Second Creek Bed represents deposition below normal wave base in a shallow, poorly circulated, dysoxic water mass, such as a large lagoon or estuary. This scenario is appealing in some respects, as it could allow even moderate storm waves to occasionally scour the sea floor, while ensuring the prevalence of dysoxic conditions within the substrate. Although graptolite rich shales are commonly associated with deep-water water depositional environments, it should be noted that this may reflect a taphonomic bias (Kaljo, 1978). The high-abundance, low-diversity occurrence of graptolites at these localities may reflect a restrictive, ecologically stressed environment such as those found in nearshore, lagoonal settings (Berry and Boucot, 1972). However, this line of reasoning must be tempered by the relatively robust conodont assemblage, and the lack of comparable graptolite occurrences in Silurian intervals of eastern North America that are of unquestionably shallow origin. Furthermore, a lack of unambiguous shallow water sedimentary structures (e.g., desiccation features, hummocky cross stratification, flaser bedding, and tidal laminites) suggests an alternative scenario may be more parsimonious.
Eckert and Brett (1989) have argued for an offshore depositional environment for these strata on the basis of macrofossil assemblages containing Eoplectodonta, Dicoelosia, and Skenidioides, all traditionally held as representatives of deep water benthic assemblages (BA-4 to BA-5; see Ziegler, 1965; Boucot, 1992). The co-occurrence of these elements suggests a water depth on the order of about 50 meters (Brett et al., 1993). In light of this, and the shortcomings of a shallow water depositional model (discussed in the previous paragraph), we conclude that a deep-water depositional environment is a more cohesive interpretation. There are many parallels between the Second Creek Bed – Williamson Formation and pyritic lag beds in Devonian black shale successions in west-central New York State. A classic example of the latter is the upper Givetian (Middle Devonian) Leicester Pyrite Member, a lenticular pyritic-phosphatic bed overlain by the alternating dark gray and black shales of the Genesee Formation (Baird and Brett, 1986, 1991). To explain deposits such as these, Baird and Brett (1986, 1991) invoke a prolonged interval of sediment starvation concomitant with a rise in sea-level following lowstand, at which time episodes of submarine erosion could have been produced by deep storm waves, density currents, or internal waves (waves propagated at the interface of water masses of different densities). Any combinations of these mechanisms could potentially explain how pyrite that formed during early diagenesis within the substrate might be exhumed and concentrated at the sea-floor. In fairly deep, dysoxic environments below the chemocline, the reducing conditions required to maintain the chemical stability of pyrite need not have ever been disrupted. Furthermore, given the lack of clastic input, the authigenic minerals and the remains of planktic organisms (e.g., conodonts) living in the upper parts of the water column would necessarily constitute a large proportion of the rock as a whole.
CONCLUSIONS
At its type locality in Wayne County, New York, the Second Creek Bed yields a conodont fauna that can be assigned to the middle Telychian Pterospathodus amorphognathoides angulatus Zone. The presence of the Aulacognathus kuehni and Apsidognathus tuberculatus ssp. n. 2 Männik, 2007a in this conodont assemblage most likely represents a fraction of reworked, older conodont elements incorporated into the bed by erosional reworking. The underlying Wolcott Furnace Hematite, sampled at the same locality, is tentatively assigned to the upper part of the Pterospathodus eopennatus Superzone.
The conodonts recovered from the Second Creek Bed at its type section are similar to those reported in this unit at Tryon Park in Rochester, New York, corroborating arguments for the bed's continuity and isochroneity. This casts further doubt on an apparently much older age for the lower Williamson Formation that is indicated by the occurrence of the graptolites Stimulograptus clintonensis. These findings also make it seem unlikely that the Second Creek Bed is laterally time-equivalent to the Westmoreland Hematite sensu stricto, though it may be correlative to some other bed that is genetically linked to the iron ore.
The high concentrations of conodont elements and authigenic minerals corroborate the interpretations of other authors that the Second Creek Bed records a time of slow sedimentation in a dysoxic environment. The pyritized internal molds of mollusks recovered from the bed are interpreted to have formed diagenetically in the substrate prior to the deposition of the bed. These pyrite grains could then have been exhumed and concentrated on the dysoxic seafloor during episodes of submarine erosion driven by density currents, deep storm waves, or internal waves. Thus, we conclude that dysoxic and highly reducing conditions prevailed even as erosion, reworking, and final deposition of the bed took place. In light of these observations, it seems most parsimonious to conclude that the Second Creek Bed and the overlying Williamson represent deposition in fairly deep marine environments below storm wave base, and below the chemocline.
ACKNOWLEDGEMENTS
All rock dissolution and sieving done as part of this study was conducted in lab space generously provided by D.L. Meyer (University of Cincinnati). We also thank K. Hedrick and S. Tritschler (both at University of Cincinnati), who offered valuable time, lab space, and the materials necessary for heavy separation of conodont elements from lighter sediment residues. Helpful comments and feedback provided by P. Männik and an anonymous reviewer made this a stronger manuscript.
N.B. Sullivan acknowledges financial support from the University of Cincinnati, a Student Assistance Grant from the Society for Sedimentary Geology (SEPM), and a Graduate Student Research Grant from the Geological Society of America (GSA). C.E. Brett acknowledges the support of NSF Grant EAR-0518511. The research presented here is a component of the Master's Thesis of N. Sullivan; this paper is a contribution to the International Geoscience Programme (IGCP) 591 "The Early to Middle Paleozoic Revolution."
REFERENCES
Aldridge, R.J. 1972. Llandovery conodonts from the Welsh Borderland. Bulletin of the British Museum, Natural History, 22:127-231.
Armstrong, H.A. 1990. Conodonts from the Upper Ordovician-Lower Silurian carbonate platform of North Greenland. Bulletin Grønlands Geologiske Undersøgelse, 159:1-151.
Baird, G.C. and Brett, C.E. 1986. Erosion on an anaerobic seafloor: Significance of reworked pyrite deposits from the Devonian of New York State. Palaeogeography, Palaeoclimatology, Palaeoecology, 57:157-193.
Baird, G.C. and Brett, C.E. 1991. Submarine erosion on the anoxic sea floor: stratinomic, palaeoenvironmental, and temporal significance of reworked pyrite-bone deposits, p. 233-257. In Tyson, R.V. and Pearson, T.H. (eds.), Modern and Ancient Continental Shelf Anoxia. Geological Society Special Publication No. 58, London.
Berry, W.B.N. and Boucot, A.J. 1972. Silurian graptolite depth zonation. Proceedings of the 24th International Geological Conference, Section 7:59-65.
Bischoff, G.C.O. 1986. Early and middle Silurian conodonts from midwestern New South Wales. Courier Forschungsinstitut Senbeckenberg, 89:1-337.
Boggs, S. 2001. Principles of Sedimentology and Stratigraphy (Third Edition). Prentice Hall, Upper Saddle River, New Jersey.
Boucot, A.J. 1992. Benthic brachiopod community changes that reflect Silurian bioevents. Proceedings of the Estonian Academy of Sciences, 41(4):193-197.
Branson, E.B. and Mehl, M.G. 1933. Conodont Studies. University of Missouri Studies, 8:5-167.
Brett, C.E., Baarli, B.G., Chowns, T., Cotter, E., Driese, S.G., Goodman, W.M., and Johnson, M.E. 1998. Early Silurian condensed intervals, ironstones, and sequence stratigraphy in the Appalachian Foreland Basin, p. 89-143. In Landing, E. and Johnson, M.E. (eds.), Silurian Cycles: Linkages of Dynamic Stratigraphy with Atmospheric, Oceanic and Tectonic Changes. New York State Museum Bulletin. New York State Museum, Albany, NY.
Brett, C.E., Boucot, A.J., and Jones, B. 1993. Absolute depths of Silurian benthic assemblages. Lethaia, 26:25-40.
Brett, C.E. and Goodman, W.M. 1996. Silurian Stratigraphy of the Type Clinton Area of Central New York, p. 200-230. In Broadhead, T.W. (ed.), Sedimentary Environments of Silurian Taconia: Fieldtrips to the Appalachians and Southern Craton of Eastern North America. University of Tennessee Department of Geological Sciences Studies in Geology, No 26, Knoxville, Tennessee.
Brett, C.E., Goodman, W.M., and LoDuca, S.T. 1990. Sequences, cycles, and basin dynamics in the Silurian of the Appalachian Foreland Basin. Sedimentary Geology, 69:191-244.
Brett, C.E. and Ray, D.C. 2005. Sequence and Event Stratigraphy of the Silurian Strata of the Cincinnati Arch Region: Correlations with New York-Ontario Successions. Proceedings of the Royal Society of Victoria, 117(2):175-198.
Brett, C.E., Tepper, D.H., Goodman, W.M., LoDuca, S.T., and Eckert, B.-Y. 1995. Revised Stratigraphy and Correlations of the Niagaran Provincial Series (Medina, Clinton, and Lockport Groups) in the Type Area of Western New York. United States Geological Survey Bulletin, 2086:1-66.
Chowns, T.M. 2006. Sequence Stratigraphy of the Silurian Red Mountain Formation; Setting for the origin of the Birmingham ironstones. Alabama Geological Society, 43rd Annual Field Trip, 1:30.
Cramer, B.D. 2009. Application of Integrated High-Resolution Biochemostratigraphy to Paleozoic Chronostratigraphic Correlation: Recalibrating the Silurian System. Unpublished Doctoral Thesis, The Ohio State University, Columbus, Ohio.
Cramer, B.D., Brett, C.E., Melchin, M.J., Männik, P., Kleffner, M.A., McLaughlin, P.I., Loydell, D.K., Munnecke, A., Jeppsson, L., Corradini, C., Brunton, F.R., and Saltzman, M.R. 2011. Revised correlation of Silurian Provincial Series of North America with global and regional chronostratigraphic units and δ13Ccarb chemostratigraphy. Lethaia, 44(2):185-202.
Eckert, B.-Y. and Brett, C.E. 1989. Bathymetry and paleoecology of Silurian benthic assemblages, late Llandoverian, New York State. Palaeogeography, Palaeoclimatology, Palaeoecology, 74:297-326.
Fisher, D.W., Isachsen, Y.W., and Rickard, L.V. 1970. Geologic Map of New York State, 1:250,000. New York State Museum Map and Chart Series No. 15, Albany, NY.
Gillette, T. 1947. The Clinton of Western and Central New York. New York State Museum Bulletin, 41:1-191.
Hunter, R.E. 1960. Iron Sedimentation in the Clinton Group of the Central Appalachian Basin. Unpublished Doctoral Thesis, Johns Hopkins University, Baltimore, Maryland.
Hunter, R.E. 1970. Facies of iron sedimentation in the Clinton Group, p. 101-121. In Fisher, G.W. (ed.), Studies of Appalachian Geology, Central and Southern. John Wiley and Sons, New York City, New York.
Jeppsson, L. 1997. A new latest Telychian, Sheinwoodian and Early Homerian (Early Silurian) Standard Conodont Zonation. Transactions of the Royal Society of Edinburgh: Earth Sciences, 88:91-114.
Jeppsson, L. and Anehus, R. 1995. A Buffered Formic Acid Technique for Conodont Extraction. Journal of Paleontology, 69(4):790-764.
Kaljo, D.L. 1978. On the Bathymetric Distribution of Graptolites. Acta Palaeontologica Polonica, 23(4):523-531.
Lin, B.-Y. and Brett, C.E. 1988. Stratigraphy and disconformable contacts of the Williamson-Willowvale interval: revised correlations of the late Llandoverian (Silurian) in New York State. Northeastern Geology, 10:241-253.
Loydell, D.K., Kleffner, M.A., Mullins, G.L., Butcher, A., Matteson, D.K., and Ebert, J.R. 2007. The lower Williamson Shale (Silurian) of New York: a biostratigraphical enigma. Geological Magazine, 144(2):225-234.
Männik, P. 1992. Taxonomy of the conodont species Ozarkodina polinclinata (Nicoll and Rexroad) in the Silurian of Estonia. Proceedings of the Estonian Academy of Sciences, 41:54-62.
Männik, P. 1998. Evolution and taxonomy of the Silurian conodont Pterospathodus. Palaeontology, 41:1001-1050.
Männik, P. 2005. Early Telychian Valgu Event - Some Preliminary Data from Estonia, p. 134-137. In Podobina, V.M. (ed.), Evolution of Life on the Earth: Proceedings of the III International Symposium, November 1-3, 2005. Tomsk State University, Tomsk, Russia.
Männik, P. 2007a. An updated Telychian (Late Llandovery, Silurian) conodont zonation based on Baltic faunas. Lethaia, 40(1):45-60.
Männik, P. 2007b. Recent developments in the Upper Ordovician and lower Silurian conodont biostratigraphy in Estonia. Estonian Journal of Earth Sciences, 56(1):35-46.
McLaughlin, P.I., Brett, C.E., and Wilson, M.A. 2008a. Hierarchy of Sedimentary Discontinuity Surfaces and Condensed Beds from the Middle Paleozoic of Eastern North America: Implications for Cratonic Sequence Stratigraphy, p. 175-200. In Pratt, B.R. and Holmden, C. (eds.), Geological Association of Canada Special Paper 48: Dynamics of Epeiric Seas. Geological Society of Canada, St. John's, Newfoundland and Labrador, Canada.
McLaughlin, P.I., Cramer, B.D., Brett, C.E., and Kleffner, M.A. 2008b. Silurian high-resolution stratigraphy of the Cincinnati Arch: Progress in recalibrating the layer-cake, p. 119-180. In Maria, A.H. and Counts, R.C. (eds.), From the Cincinnati Arch to the Illinois Basin. Geological Field Excursions along the Ohio River Valley: Geological Society of America Field Guide 12. The Geological Society of America Inc. Boulder, Colorado.
McLaughlin, P.I., Emsbo, P., and Brett, C.E. 2012. Beyond black shales: the sedimentary and stable isotope records of oceanic anoxic events in a dominantly oxic basin (Silurian; Appalachian Basin, USA). Palaeogeography, Palaeoclimatology, Palaeoecology, 367-368:153-177.
Melchin, M.J., Sadler, P.M., Cramer, B.D., Cooper, R.A., Gradstein, F.M., and Hammer, O. 2012. The Silurian Period, p. 525-558. In Gradstein, F.M., Ogg, J.G., Schmitz, M., and Ogg, G. (eds.), Geologic Timescale 2012. Elsevier B. V., Waltham, Massachusetts.
Mostler, H. 1967. Conodonten aus dem tieferen Silur der Kitzbühler Alpen (Tirol). Annalen des Naturhistorischen Museums in Wien, 71:295-303.
Munnecke, A. and Männik, P. 2009. New biostratigraphic and chemostratigraphic data from the Chicotte Formation (Llandovery, Anticosti Island, Laurentia) compared with the Viki core (Estonia, Baltica). Estonian Journal of Earth Sciences, 58(3):159.
Nicoll, R.S. and Rexroad, C.B. 1968. Stratigraphy and conodont paleontology of the Salamonie Dolomite and Lee Creek Member of the Brassfield Limestone (Silurian) in southeastern Indiana and adjacent Kentucky. Indiana Geological Survey Bulletin, 40:1-73.
Rexroad, C.B. and Richard, L.V. 1965. Zonal Conodonts from the Silurian Strata of the Niagara Gorge. Journal of Paleontology, 39(6):1217-1220.
Sadler, P.M., Cooper, R.A., and Melchin, M. 2009. High-resolution, early Paleozoic (Ordovician-Silurian) time scales. Geological Society of America Bulletin, 121(5-6):887-906.
Verniers, J., Van de Moortel, I., Steeman, T., Mortier, J., Vandenbroucke, T., Cramer, B.D., Brett, C.E., and Kleffner, M.A. 2012. The Tryon Park controversy: a combined chitinozoan and conodont restudy. In Cramer, B.D., and Brett, C.E. (eds.), IGCP 591 Annual Meeting. Cincinnati, Ohio.
Walliser, O.H. 1964. Conodenten des Silurs. Abhandlungen des Hessischen Landesamtes für Bodenforschung, 41:1-106.
Ziegler, A.M. 1965. Silurian Marine Communities and Their Environmental Significance. Nature, 270:270-272.
