

 Symbiosis in corals and stromatoporoids from the Silurian of Baltica
Symbiosis in corals and stromatoporoids from the Silurian of Baltica
Article number: 25.2.a17
https://doi.org/10.26879/1206
Copyright Paleontological Society, June 2022
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Appendix
Submission: 8 January 2022. Acceptance: 17 May 2022.
ABSTRACT
The large collection of thin sections of stromatoporoids and corals from the Silurian of Ukraine, Moldova, Belarus, and Komi Republic (Russian Federation) revealed several incidences of skeletal intergrowth between stromatoporoids/corals and the other invertebrates. The stromatoporoids formed symbiotic associations with soft-bodied worms (Helicosalpinx and Chaetosalpinx), calcareous tentaculitoid tubeworms (microconchids, Cornulites, Conchicolites), and rugosans. Tabulate corals formed symbiotic associations with cornulitids. The studied stromatoporoid based associations are dominated by bioclaustrations of worms without mineral skeletons. Most likely non-mineralized invertebrates benefitted more from endobiotic life mode than invertebrates with mineralized skeletons as the latter already had protection on their own against predators. There was almost no difference in the number of symbiont taxa per host stromatoporoid species indicating that all studied stromatoporoids were rather similar in their tolerance towards different endobionts.
Tamara Borisenko. Geological Survey of Ukraine, Kyiv, Ukraine. tamaraborisenko2@gmail.com
Olev Vinn. University of Tartu, Institute of Ecology and Earth Sciences, Ravila 14A, 50411 Tartu, Estonia. olev.vinn@ut.ee
Volodymyr Grytsenko. National Natural History Museum NAS of Ukraine, Kyiv, Ukraine. favosites@ukr.net
Ion Francovschi. University of Bucharest, Faculty of Geology and Geophysics, Bucharest, Romania, and Institute of Geology and Seismology, Chișinău, Republic of Moldova. frankovski.ww@gmail.com
Yury Zaika. Unitary Enterprise “Geoservice”, Maura 53, 220015, Minsk, Belarus. yu_z@tut.by
Keywords: Calcareous sponges, tabulate corals, endobionts, bioclaustrations, eastern Europe, Silurian
Final citation: Borisenko, Tamara, Vinn, Olev, Grytsenko, Volodymyr, Francovschi, Ion, and Zaika, Yury. 2022. Symbiosis in corals and stromatoporoids from the Silurian of Baltica. Palaeontologia Electronica, 25(2):a17. https://doi.org/10.26879/1206
palaeo-electronica.org/content/2022/3618-symbiosis-in-the-silurian
Copyright: June 2022 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
There is little doubt that symbiosis has been an important driving force of biological evolution. The relationships between animal diversification and parasitism have been recently studied by De Baets et al. (2021b). However, many fundamental aspects of the evolution of symbiotic relationships have remained poorly understood, especially as compared to the evolutionary history of predation (Huntley and Kowalewski, 2007). Trace fossils often play a pivotal role in the study of parasite-host interactions in deep time (Huntley and De Baets, 2015). The study of symbiotic relationships in fossil animals involves various difficulties (De Baets et al., 2021a,b). It is mostly impossible to distinguish mutualism from commensalism or parasitism (Taylor, 2015; Robin, 2021). Epibionts often use skeletons of dead organisms as a hard substrate for their attachment. However, in the latter case, there is no evidence of intergrowth between two organisms. The intergrowth of two organisms provides us with the best evidence of symbiosis in the fossil record (Tapanila, 2008). We found multiple cases of intergrowth when reviewing our collection of Silurian stromatoporoids and tabulates. The majority of intergrown organisms in our collection are stromatoporoids as hosts but in some cases, it also involves tabulate corals. Frequently symbiotic invertebrates are embedded within the skeleton of a host organism, leaving only small apertures exposed for feeding and such embedment structures are called bioclaustrations (Palmer and Wilson, 1988). Ancient symbiotic relations are especially important for the study of coevolution. Symbiosis is here viewed as any type of close and long-term biological interaction between two different animals, whether it is parasitic, commensalistic, or mutualistic (Martin and Schwab, 2013).
The Silurian of Baltica has a rich record of symbiosis (Vinn and Wilson, 2016). There are records of possible polychaetes (Mõtus and Vinn, 2009; Vinn and Mõtus, 2014b), rugosans (Nestor, 1966; Kershaw, 1987; Vinn and Mõtus, 2014a), syringoporids (Nestor, 1966; Kershaw, 1987), and cornulitids (Vinn and Wilson, 2010a, b) in tabulate corals and stromatoporoids from the Silurian of Baltica.
The aims of this study are as follows: (1) to summarize the symbiotic associations found in the Silurian of Ukraine, Moldova, Belarus, and Komi Republic (all regions belonging to Baltica); (2) to discuss symbiosis in Silurian stromatoporoids; and (3) to discuss symbiosis in Silurian tabulates.
HISTORY OF GEOLOGICAL STUDIES
The study of the Silurian from Podillya commenced at the beginning of the nineteenth century by Staszic, Eichwald, and Malevsky. Their findings and observations are discussed in the monography of Kozlowsky (1929). The first stratigraphic schemes of the Upper Silurian and Lower Devonian deposits were created by Stur (1872). Alth (1874) established “the Skala beds” and described the fossils from the outcrops which he analysed. Venyukov (1899) identified and described new corals, brachiopods, bivalves, and other fossil species and established three coral horizons in the lower part of the Silurian which outcropped on the left bank of the Dniester River. A lot of details to the Silurian and Lower Devonian sequence from Podillya were added by Kozlowsky (1929) and Văscăuțanu (1931). Kozlowsky investigated the left bank of the Dniester River, specifically the upper course of the Zbruch River, while Văscăuțanu studied the right bank of the Dniester River. Kozlowsky identified the Skalian, Borschovian, and Chortkovian Regional Stages in the upper part of the sequence.
One of the first stratigraphic schemes that generalized previous discoveries was established by Lungershausen and Nikifiriova (1942). They identified seven horizons: Kitaigorod, Mouksha, and Ustia belonging to the Llandovery epoch; Malynivtsy - Wenlock epoch; Skala - lower Ludlow and Aimestri, Borschov - upper Ludlow; Chortkov - Přidoli epoch. Nikiforova continued to study the brachiopods from the Silurian of Podillya after the Second World War (Nikiforova, 1954). The famous Russian geologist Sokolov published a monograph related to tabulate corals from this region (Sokolov, 1955). Dikenstein (1957) divided the Kitaigorod horizon into the Restevo and Grushevtsy beds and the Skalian horizon into the Isakivtsy, Prigorodok, Rashkiv, and Volkivtsy beds. Boucot and Pankivsky (1962) considered that the boundary between the Silurian and the Devonian is at the base of the Skalian horizon, which they considered to belong to the Devonian system. Sytova (1966) divided the Malinivtsy horizon into the Konivka, Sokil, and Grinchuk beds. Drygant (1967, 1968) identified exclusive species of conodonts which allowed the marking of the exact boundary between the Silurian and Devonian in the Podillyan sequence. The same results were obtained by Tsegelnjuk (1968), Koren (1968), and others.
Drygant and Tsegelnjuk (1968) proved that the previously considered sterile Llandovery deposits from the Silurian of Podillya are fossiliferous by finding conodonts and graptolites in the outcrops located near the Kitaigorod village. Corals were studied by Bondarenko (1982), Grytsenko (1977a, b, 1987). The ecostratigraphical approach toward the Silurian sequence and fossils was used by Grytsenko et al. (1999).
The materials were gathered from hundreds of drilled boreholes, outcrops, and a significant number of fossils from collections were reconsidered for analysis by a group of researchers from Kyiv, Lviv (Ukraine), St. Petersburg, Moscow, and Yekaterinburg (Russia). They summarized previously published data as well as new information regarding the Silurian system and released the guide for geological excursions of the International Subcommission on Silurian Stratigraphy of the International Union of Geological Sciences (Tsegelnjuk et al., 1983). The guide contained an improved stratigraphic scheme. All fossil findings were linked to the new detailed scheme. The authors presented a new approach toward correlating the available scheme with the international stratigraphical scheme based on the geological development of the basin, which is attached to the western ridge of Baltica. The sedimentary cover of the basin did not suffer any significant metamorphic changes and tectonic activities. The different degrees of cyclic movements are reflected in the lithological features and allowed to demarcate the local geological groups: Bolotine (Llandovery), Jaruga (Wenlock), Malynivtsy (Ludlow), Skala (Přidoli) and stages: Wenlock, Ludlow, Ulichian (after Ludlow and before Přidoli), and Skalian. Also, a new Graptolitic zonal subdivision was proposed. Additionally, the guide contained 65 tables with the distribution of all studied fossils: brachiopods, trilobites, Tabulata and rugosan corals, Heliolitida, Stromatoporata, conodonts, Chitinozoa, algae, cephalopods, ostracodes, and graptolites.
The International Subcommission on Silurian Stratigraphy (ISSS) decided to choose the stratotype in Czechia. Among other applicants, the Silurian from Podillya was chosen to be the parastratotype for the boundary between the Silurian and Devonian. Nevertheless, the Silurian section has some advantages. Firstly, the entire sequence is without any significant interruptions, metamorphic changes, or tectonic activities and is rich in different kinds of fossils. Several outstanding outcrops and thousands of drilled boreholes showed the cyclic development of the Silurian Sea basin from the Llandovery to the Gedinnian (Tsegelnjuk et al., 1983). The final version of the Silurian scheme of Ukraine was published in the last work of Konstantinenko and Tsegelnjuk (2013).
GEOLOGICAL SETTING
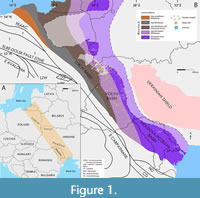 The Silurian deposits are located along the southwestern and western margins of Baltica and have different areas of extent which depend on the local tectonic structure (Figure 1).
The Silurian deposits are located along the southwestern and western margins of Baltica and have different areas of extent which depend on the local tectonic structure (Figure 1).
The Silurian from the eastern part of the Moldova-Podillya basin (southwestern Baltica) has minimal facies variations and is mostly consistent throughout the entire region (Bukatchuk et al., 1988). It is divided into the lower (Llandovery and Wenlock) and upper (Ludlow and Pridoli) sequences. The Llandovery sediments are represented by marine limestones in the lower part and by claystones in the upper part of the sequence (Tsegelnjuk and Bukatchuk, 1974). The Wenlock deposits begin with marine limestones, which are covered by lagoonal limestones and dolostones (Tsegelnjuk, 1980a). The Ludlow and Přidoli sediments are defined by various limestones and dolostones, which were deposited in lagoonal, inner to outer shelf, and shallow-water depositional environments (Tsegelnjuk, 1980a, b).
The western part of the Moldova-Podillya basin is less complete than the eastern part of the platform (Pătruț, 1981). It is divided into two sectors, the Bătrânești - Todireni - Iași sector and the Rădăuți sector (Iordan, 1975; Pătruț, 1981). Both of them start with a thin sequence of Llandovery siliciclastic sediments, followed by a thick sequence represented by predominantly calcareous Wenlock - Lower Ludlow deposits. In the Bătrânești - Todireni - Iași sector the limestones from the lower part of the upper sequence are covered by calcareous claystones with ostracods. However, in the Rădăuți sector, these layers are interposed by a thick sequence of Ludlovian graptolitic claystones. Pătruț (1981) considers that the graptolitic claystones are older than the Lower Ludlow upper limestones with brachiopods from the Bătrânești - Todireni - Iași sector, nonetheless they could also represent a lateral facies variation of these limestones or, at least, of their terminal part.
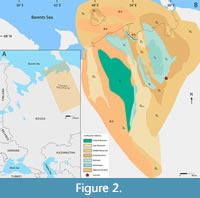 The Volyn basin has a structure similar to a fragmented plate due to the presence of several faults. The territory corresponding to modern-day Belarus is defined by the deep Podlasie-Brest Depression. The Peribaltic basin developed in shallow water conditions and the sedimentary sequence is defined by the presence of several hiatuses, which is especially visible in the outcropped nearshore sections. The section in the north of Russia (Timan-Pechora basin) is located in a depression (Figure 2). In this region, outcrops are rare and can be found in the incised valleys of some major rivers (e.g., Kozhym River). The most recent stratigraphical scheme of the Silurian deposits is very complicated and reflects the complex features of Baltica. Facies regions that are described in the scheme have different lithological compositions which depend on the depositional depth as well as the lithological characteristics. The slope of the Silurian Sea basin shows that there is a gradual facies change of the deposits, starting with a lagoon, to a nearshore environment, to a shallow-water facies with corals (Venyukov, 1899; Grytsenko, 1977a, b; Grytsenko, 2007) and finally transitioning to a deep water one with graptolites.
The Volyn basin has a structure similar to a fragmented plate due to the presence of several faults. The territory corresponding to modern-day Belarus is defined by the deep Podlasie-Brest Depression. The Peribaltic basin developed in shallow water conditions and the sedimentary sequence is defined by the presence of several hiatuses, which is especially visible in the outcropped nearshore sections. The section in the north of Russia (Timan-Pechora basin) is located in a depression (Figure 2). In this region, outcrops are rare and can be found in the incised valleys of some major rivers (e.g., Kozhym River). The most recent stratigraphical scheme of the Silurian deposits is very complicated and reflects the complex features of Baltica. Facies regions that are described in the scheme have different lithological compositions which depend on the depositional depth as well as the lithological characteristics. The slope of the Silurian Sea basin shows that there is a gradual facies change of the deposits, starting with a lagoon, to a nearshore environment, to a shallow-water facies with corals (Venyukov, 1899; Grytsenko, 1977a, b; Grytsenko, 2007) and finally transitioning to a deep water one with graptolites.
Stromatoporoids appeared in the sequences of Baltica later than in North America, North China, or Australia where the earliest indubitable stromatoporoids have been already recorded from the Darriwilian strata. It has been explained by the location of the Baltica paleocontinent in the southern temperate climate zone until the Katian time when it finally migrated into the equatorial belt (Nestor, 1997). In the Llandovery, clathrodictyids were diverse and abundant, and Clathrodictyon and Ecclimadictyon became dominating genera. They could constitute more than 80% of stromatoporoid specimens. Labechiids (Pachystylostroma, Forolinia, Rosenella, and Labechia) were also abundant but slightly less frequent than clathrodictyids. During the Wenlock, the abundance of stromatoporids increased and their diversification continued (Nestor, 1997). During the Ludlow, the diversity of the stromatoporoid fauna reached its peak in Baltica. In different taxonomical branches new taxa appeared, e.g., Lophiostroma among labechiids, Plexodictyon among clathrodictyids, Pseudolabechia in Actinostromatida, Syringostromella among stromatoporids, and Parallelostroma - the first representative of the Order Syringostromatida in the sequences of Baltica (Nestor, 1997). Baltic stromatoporoids were highly facies-dependent organisms with a relatively narrow ecological niche (Nestor, 1997). The most abundant and diverse stromatoporoid association occurred in the high-energy shoal facies belt, represented in the fossil record by coral-stromatoporoid boundstones, skeletal and coquinite grain- and rudstones (Nestor, 1997). Stromatoporoids were also rather numerous in the moderate- to low-energy open-shelf facies belt where biomicritic deposits were accumulated (Nestor, 1997).
The earliest Silurian was a time of rapid diversification of Mesofavosites and Paleofavosites (Mõtus, 1997). In the middle of Llandovery, Multisolenia, and Parastriapora appeared and favositids became the dominant group of tabulates in the Silurian of Baltica (Mõtus, 1997). The Wenlock was characterized by major changes in the tabulate fauna. Syringolites, Thecia, and Mastopora appeared whereas Mesofavosites and Catenipora disappeared in the early Wenlock. During the middle Wenlock, the diversity of tabulates increased again (Mõtus, 1997). In the Ludlow, tabulates had a low generic diversity in Baltica and Favosites was the most important component of the tabulate fauna (Mõtus, 1997). A few representatives of Thecia, Romingerella, Laceripora, and Syringopora have also been found from the Ludlow (Mõtus, 1997).
MATERIAL AND METHODS
 The collection (8000 thin sections, including about 100 with examples of symbiosis containing approximately 30% stromatoporoid and 70% coral specimens) was collected for more than 50 years by the scientists of the National Natural History Museum NAS of Ukraine. The fieldwork started with the investigation of a couple of boreholes drilled in Podillya in 1966. Later on, similar fieldwork took place every year in Ukraine (Podillya) (Appendix 1). In addition, a few field trips visited Moldova, Belarus, Estonia and Komi Republic. Most of the samples were collected from outcrops and boreholes drilled during geological prospecting in western Ukraine, Belarus, and Moldova (Figure 3 and Figure 4). All studied thin sections were prepared in the laboratories in Ukraine, Moldova, and Belarus. The endobionts in thin sections were discovered using a light microscope MBS-9 LOMO. All endobionts were photographed using a digital camera (USB digital microscope, made in China and SONY DSC-H7).
The collection (8000 thin sections, including about 100 with examples of symbiosis containing approximately 30% stromatoporoid and 70% coral specimens) was collected for more than 50 years by the scientists of the National Natural History Museum NAS of Ukraine. The fieldwork started with the investigation of a couple of boreholes drilled in Podillya in 1966. Later on, similar fieldwork took place every year in Ukraine (Podillya) (Appendix 1). In addition, a few field trips visited Moldova, Belarus, Estonia and Komi Republic. Most of the samples were collected from outcrops and boreholes drilled during geological prospecting in western Ukraine, Belarus, and Moldova (Figure 3 and Figure 4). All studied thin sections were prepared in the laboratories in Ukraine, Moldova, and Belarus. The endobionts in thin sections were discovered using a light microscope MBS-9 LOMO. All endobionts were photographed using a digital camera (USB digital microscope, made in China and SONY DSC-H7).
DESCRIPTIONS OF FOSSILS AND SYMBIOTIC ASSOCIATIONS
Stromatoporoid Based Associations
 Silurian stromatoporoids from Baltica host a rich fauna of endobionts. In the studied collection, 22 stromatoporoids contain Helicosalpinx bioclaustrations, 18 stromatoporoids revealed Chaetosalpinx bioclaustrations, 13 stromatoporoids contain syrigoporiids intergrown with stromatoporoid, six stromatoporoids contain embedded Palaeoconchus microconchids, six stromatoporoids host the cornulitids Cornulites and Conchicolites, two stromatoporoids contain intergrown rugosans.
Silurian stromatoporoids from Baltica host a rich fauna of endobionts. In the studied collection, 22 stromatoporoids contain Helicosalpinx bioclaustrations, 18 stromatoporoids revealed Chaetosalpinx bioclaustrations, 13 stromatoporoids contain syrigoporiids intergrown with stromatoporoid, six stromatoporoids contain embedded Palaeoconchus microconchids, six stromatoporoids host the cornulitids Cornulites and Conchicolites, two stromatoporoids contain intergrown rugosans.
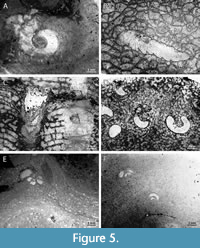 Helicosalpinx concoenatus (Figure 5A, D; Figure 6B) occurs in Densastroma sp., Clathrodictyon mukshiensis, Clathrodictyon sp., Vikingia sp., Plexodictyon sp., Plectostroma podolicum, and Perplexostroma savaliensis. Tubes of H. concoenatus lack their own wall; they are helical and oriented more or less perpendicular to the host stromatoporoid’s growth lamellae (Figure 5A, D; Figure 6B).
Helicosalpinx concoenatus (Figure 5A, D; Figure 6B) occurs in Densastroma sp., Clathrodictyon mukshiensis, Clathrodictyon sp., Vikingia sp., Plexodictyon sp., Plectostroma podolicum, and Perplexostroma savaliensis. Tubes of H. concoenatus lack their own wall; they are helical and oriented more or less perpendicular to the host stromatoporoid’s growth lamellae (Figure 5A, D; Figure 6B).
Chaetosalpinx bioclaustrations (Figure 6B) occur in Clathrodictyon sp., Clathrodictyon microstriatellum, Vikingia sp., Densastroma sp., Perplexostroma savaliensis, and Parallelostroma pulchra. Tubes of Chaetosalpinx are straight to irregularly curved (Figure 6B). They lack their own wall and are oriented more or less perpendicular to the growth lamellae of host stromatoporoid (Figure 6B).
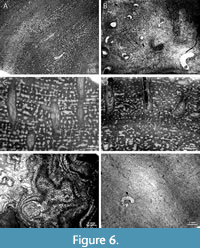 Syringoporiid endobionts (Figure 6C, D) occur in Parallelostroma communis, Parallelostromapora pulchra, Parallelostroma sp., and Clathrodictyon sp. Syringoporiids are oriented perpendicular to the lamellae of host stromatoporoid (Figure 6C, D).
Syringoporiid endobionts (Figure 6C, D) occur in Parallelostroma communis, Parallelostromapora pulchra, Parallelostroma sp., and Clathrodictyon sp. Syringoporiids are oriented perpendicular to the lamellae of host stromatoporoid (Figure 6C, D).
Microconchid Palaeoconchus sp. (Figure 5E, F) occurs in Parallelostroma sp., Clathrodictyon mukschiense, Clathrodictyon planum, and Plectostroma podolicum. Tubes of Palaeoconchus are planispiral and oriented more or less parallel to the growth lamellae of host stromatoporoid (Figure 5E, F).
Cornulites is intergrown with Stromatopora sp. and Conchicolites sp. occurs in Clathrodictyon microstriatellum and Pseudolabechia sp. Tubes of cornultids are crossing growth lamellae of host stromatoporoid at various angles (Figure 6A, E).
Clathrodictyon planum, Plexodictyon sp. and Stromatopora sp. host single species of endobionts. Clathrodictyon microstriatellum, Clathrodictyon mukshiensis, Densastroma sp., Parallelostroma sp., Parallelostroma communis, Parallelostromapora pulchra, Perplexostroma savaliensis, Plectostroma podolicum, and Vikingia sp. host two species of endobionts. Clatrodictyon sp. hosts three species of endobionts.
Tabulate Based Associations
Studied tabulates host only cornulitids (Figure 5B). Cornulites sp. is intergrown with Heliolites sp. and Mesosolenia sp. Cornulitids are oriented at various angles regarding the growth lamellae of host tabulates. There are no obvious changes in the morphology of host corals near the cornulitid endobionts (Figure 5B). It should be noted that in the northeastern part of Baltica, the role of tabulate corals (primarily favositids) as hosts of endobionts increases significantly in the Lochkovian and possibly in the uppermost Přidolian. Numerous favositid specimens hosting endobionts (Phragmosalpinx australiensis) occur in the Lochkovian Ovinparma Formation in the western slope of North Urals and Tchernychev Uplift. At the same stratigraphic level, similar endobionts are also often found in Favositids from other Arctic regions: Vaigach Island, Novaya Zemlya, Taimyr (Zaika, 2007).
DISCUSSION
Endobionts
The studied stromatoporoid based associations are dominated by bioclaustrations of worms without mineral skeleton, Helicosalpinx concoenatus being the most abundant. Among skeletonized endobionts syringoporiids were most abundant making up more than half of all skeletonized endobionts in stromatoporoids. Syringoporiids are also one of the major groups of endobiont in stromatoporoids from the Ludlow of Estonia and Gotland (both Baltica) (Kershaw, 1987; Kershaw and Mõtus, 2016), and they indicate a similar situation in our sections. Tentaculitoid tubeworms (i.e., microconchids and cornulitids) are more common in the studied Silurian stromatoporoids than rugosans. A strikingly small number of endobiotic rugosans in the studied thin sections likely represents a sampling bias as such associations are very diverse and numerous in the Silurian of Estonia and Gotland (Baltica) (Kershaw, 1987; Vinn and Wilson, 2016). Among tentaculitoid tubeworms, cornulitids are known to form symbiotic associations with stromatoporoids in the Sheinwoodian of Estonia (Vinn and Wilson, 2010a) but embedded microconchids have not been previously reported from stromatoporoids. It remains unresolved whether microconchid-stromatoporoid associations represent intergrowth of two organisms or an overgrowth. Based on the similarity with endobiotic microconchids in the bryozoans from the Přidoli of Saaremaa, Estonia it is possible to hypothesize that some microconchids may have had endobiotic life mode within stromatoporoids in the Silurian of Baltica. Alternatively, microconchids encrusted some dead spots on the living stromatoporoid and were thereafter overgrown and killed by the host. Abundant worm biocluastrations in stromatoporoids are known from the Ludlow of Saaremaa, Estonia but our data suggest that worm biocluastrations may have been even more common in the stromatoporoids from the Silurian of Baltica. Worm bioclaustration without mineral skeleton forms a major part of endobiont fauna in the Silurian tabulates (Tapanila, 2005; Mõtus and Vinn, 2009), and it is likely that they also formed a major part of endobiont fauna of the Silurian stromatoporoids. One could ask why non-mineralized worms dominated both tabulate and stromatoporoid based symbiotic associations. The most likely answer is that non-mineralized invertebrates benefited more from endobiotic life mode than invertebrates with mineralized skeletons as the latter already had protection on their own against the predators. Another likely answer is that, among the Silurian faunas, some worms lacking a mineralized skeleton developed adaptive strategies that led to symbiotic intergrowth, possibly by means of the ability to settle on a living substrate that was not suitable for other potential epibionts. Examples of specialized, host-specific symbiotic taxa occur among recent faunas with well-documented cases such as the hydroid familiy Zancleidae, with species that are known to be symbionts of corals and bryozoans. Spirally coiled and non-spiral worm bioclaustrations are about equally common in the studied stromatoporoid collection indicating that both of these morphologies were equally successful within a stromatoporoid host. Endobiotic cornulitid occur in the early Silurian tabulates from Canada (Dixon, 2010). However, such an association has not been previously described from the Silurian of Baltica (Vinn and Wilson, 2016). Thus, it is possible that cornulitid-tabulate symbiosis may have had wider geographical or even global distribution in the Silurian. The majority of the studied stromatoporoid thin sections and all tabulate thin sections contain just single species of endobionts. Only Helicosalpinx and Chaetosalpinx occur together in the same thin section. This occurrence may indicate that stromatoporoids and tabulates were seldom infested by several species of endobionts but stromatoporoid infested by multiple endobiont species were common in the Ludlow of Saaremaa and Gotland. Moreover, thin sections do cover only a small section of the host, and thus it is more likely that most endobionts specimens simply avoided close vicinity of the specimens of another endobiont species. Such distance between endobionts could be due to feeding competition as most of the studied endobionts were suspension feeders with the exception of micropredatory rugosans. Endobionts selectively infested host stromatoporoids. Some endobionts such as Helicoslapinx and Chaetosalpinx mostly occur in the same taxa of stromatoporoids whereas syringoporiids infested a completely different set of stromatoporoid taxa. The similarity in host taxa of worm bioclaustration could indicate that Helicosalpinx and Chaetosalpinx were ecologically similar. On the other hand, syringoporiids and worms responsible for Helicosalpinx and Chaetosalpinx bioclaustrations likely had different ecologies. This is not surprising as worms were likely suspension feeders and syringoporiids as all corals were micropredators.
Hosts
The number of stromatoporoids with endobionts exceeds several times the number of endobionts in tabulates which at least partially can be explained by a collecting bias. However, Vinn and Wilson (2016) suggested that stromatoporoids were the most common hosts to various endobionts in the Silurian of Baltica and our data here support this view. Baltic tabulates usually contain worm biocluastrations (i.e., Chaetosalpinx) in large numbers (Mõtus and Vinn, 2009; Klaamann, 1958), and the lack of such specimens in the studied collection likely represents a collecting bias. All studied Silurian stromatoporoids and tabulate species hosted only a small number of endobiont taxa. There was only little or no difference at all in the number of symbiont taxa per host stromatoporoid species. This demonstrates that all of the studied stromatoporoids were rather similar in their tolerance toward different endobionts. It also indicates that all host stromatoporoids created a specific enough environment for their symbionts that could possibly fit only with the requirements of a few endobiont taxa.
CONCLUSIONS
The data from the studied collection confirm that stromatoporoids were the most common hosts to various endobionts in the Silurian of Baltica. Bioclaustrations of non-mineralized worms likely formed a major part of the endobiont fauna of the Silurian stromatoporoids and in this respect, stromatoporoids were similar to the Silurian tabulates. Microconchids formed symbiotic associations with the stromatoporoids in the Silurian of Baltica. The cornulitid-tabulate symbiosis may have had wide geographic or even global distribution in the Silurian. Most endobiont specimens avoided close vicinity of the specimens of another endobiont species due to feeding competition. Endobionts infested selectively the host stromatoporoids and all of the studied Silurian stromatoporoids and tabulate species hosted only a small number of endobiont taxa.
ACKNOWLEDGEMENTS
This research was funded by the Estonian Research Council grant number PRG836, a Research Grant of the Institute of Ecology and Earth Sciences, University of Tartu, and the Paleontological Society Sepkoski Grant 2021. The work was prepared in accordance with the project “Geological diversity”. Participation by IF was supported by the national project GeoSeism 86-PS. We are very grateful to our assistants A. Koba and N. Okonishnikova from the Geological Department of the Kyiv National University “Taras Shevchenko” who have prepared thin sections during the last year and other unknown for us specialists from Moldova and Belarus. Many thanks to G. Vinogradov from the Geological Survey of Ukraine for assistance during fieldwork. We thank the administration of the Kyiv National University and National Natural History Museum for the financial support of fieldwork (Project NAS of Ukraine No 0117U000261 “Geological diversity of Volyn-Podillya plate and development of recommendations deal with conservation of Geoheritage”). This study was supported by the Project PN-III-P1-1.1-PD-2021-0400, No. 60/2022 of the Romanian Executive Agency for Higher Education, Research, Development and Innovation, UEFISCDI to I.F.
REFERENCES
Alth, A. 1874. Über die Paleozoischen Gebilde Podoliens und deren Versteinerungen. Abhandlungen der Kaiserlich-Königlichen Geologischen Reichsanstalt (Österreich), 1:1-78.
Bondarenko, O.B. 1982. Pridolian Heliolitida: variability, morphogenesis, bios-tratigraphy. Izvestija Akademii Nauk SSSR, serija geologicheskaja, 5:46-58.
Boucot, A. and Pankiwskyi, K. 1962. Llandoverian to Devonian stratigraphy of Podolia and adjacent Moldavia. Symposium Band Bonn-Bruxelles, 2:1-11.
Bukatchuk, P.D., Bliuk, I.B., and Pokatilov, V.P. 1988. Geological map of the Moldavian Soviet Socialist Republic, scale 1:200 000. Explanatory notes. Moldovageology, Chișinău. (In Russian)
De Baets, K.D., Huntley, J.W., Klompmaker, A.A., Schiffbauer, J.D., and Muscente, A.D. 2021a. The fossil record of parasitism: Its extent and taphonomic constraints, p. 1-50. In De Baets, K.D. and Huntely, J.W. (eds.), The Evolution and Fossil Record of Parasitism. Springer, Cham. https://doi.org/10.1007/978-3-030-52233-9_1
De Baets, K., Huntley, J.W., Scarponi, D., Klompmaker, A.A., and Skawina, A. 2021b. Phanerozoic parasitism and marine metazoan diversity: dilution versus amplification. Philosophical Transactions of the Royal Society B, 376:20200366. https://doi.org/10.1007/978-3-030-52233-9_1
Dikenstein, G. Kh. 1957. The paleozoic deposits of southwestern Russian platform. Gostoptekhizdat, Moscow. (In Russian)
Dixon, O.A. 2010. Endobiotic cornulitids in Upper Ordovician tabulate corals and stromatoporoids from Anticosti Island, Quebec. Journal of Paleontology, 84:518-528.
Drygant 1967. Nekotorye dannye o conodontovykh zonakh i vozraste perekhodnykh sloev ot Silura k Devonu v Podolii. Paleontological review, 4(2):56-59. (In Russian)
Drygant, D.M. 1968. Nekotorye vidy conodontov Silura Podolii. Paleontological review, 5(1):46-52. (In Russian)
Drygant, D.M. and Tsegelnjuk, P.D. 1968. Regarding the age of the Resteve and Kitaigorod horizons of the Silurian of Podillya. Reports of the Academy of Sciences UkrSSR. Series B., 12:1066-1069. (In Ukrainian)
Grytsenko, V.P. 1977a. The coral reefs of the Silurian of Podolia. Collection of scientific works of the Kyiv University. Kyiv, Vyscha School, 13:5-11. (In Russian)
Grytsenko, V.P. 1977b. The shallow water facies and corals of the Silurian of Volyn-Podillya. Kyiv, Preprint of the Institute of Geological Sciences, Academy of Sciences UkrSSR, 1:87-21. (In Russian)
Grytsenko, V. 1987. K paleoecologii corallov Silura Volyno-Podolii. Naukova dumka, 1:23-26. (In Russian)
Grytsenko, V. 2007. Distribution of Corals from the Silurian of the Podolian Shelf / Fossil Coral and Sponges. Schriftenreihe der Erdwissenschaftlichen Kommissionen, 17:185-198. https://doi.org/10.1553/0x001597b4
Grytsenko, V., Istchenko, A., Konstantinenko, L., and Tsegelnjuk, P. 1999. Animal and plant communities of Podolia, p. 462-487. In Boucot, A.J. and Lawson, J.D. (eds.), Paleocommunities: A case study from the Silurian and Lower Devonian. New York, Cambridge University Press.
Huntley, J.W. and Kowalewski, M. 2007. Strong coupling of predation intensity and diversity in the Phanerozoic fossil record. Proceedings of the Natural Academy of Sciences, 38:15006-15010. https://doi.org/10.1073/pnas.0704960104
Huntley, J.W. and De Baets, K. 2015. Trace fossil evidence of trematode-bivalve parasite-host interactions in deep time: Advances in Parasitology, 90:201-231. https://doi.org/10.1016/bs.apar.2015.05.004
Iordan, M. 1975. Studiul biostratigrafic al Paleozoicului din forajul Bătrânești (Platforma Moldovenească). Dări de seamă ale Institutului de Geologie și Geofizică, 4:27-50. (In Romanian)
Kershaw, S. 1987. Stromatoporoid-coral intergrowths in a Silurian biostrome. Lethaia, 20:371-380. https://doi.org/10.4202/app.00094.2014
Kershaw, S. and Mõtus, M-A. 2016. Palaeoecology of corals and stromatoporoids in a late Silurian biostrome in Estonia. Acta Palaeontologica Polonica, 61:33-50. https://doi.org/10.4202/app.00094.2014
Klaamann, E. 1958. Uue fossiilse ussi leiust eesti aluspõhjas. Eesti Loodus, 5:306-307.
Konstantinenko, L.I. and Tsegelnjuk, P.D. 2013. Section 5, p. 176-195. In Gozhik, P.F. (ed.), Silurian System. Stratigraphy of Upper Proterozoic and Phanerozoic of Ukraine. V.1. Stratigraphy of Upper Proterozoic, Paleozoic and Mesozoic of Ukraine. Academy of Sciences UkrSSR, Kyiv. (In Ukrainian)
Koren, T.N. 1968. Pervye nakhodki rannedevonskikh monogtaptid v borschovskom horizonte Podolii. Doklady AN USSR, 182(4):938-949. (In Russian)
Kozlowsky, R. 1929. Les brachiopods gothlandiens de la Podolie Polonaise. Palaeontologica Polonica, 1:1-254.
Lungershausen, L.F. and Nikifiriova, O.I. 1942. O stratigraficheskom otnoshenii siluriiskikh sloev Podolii k analogichnym slojam nekotorykh drugikh mest Zapadnoi Europy. Doklady Akademii Nauk USSR, 34(2):69-74. (In Russian)
Martin, B.D. and Schwab, E. 2013. Current usage of symbiosis and associated terminology. International Journal of Biology, 5:32-45. https://doi.org/10.5539/ijb.v5n1p32
Modlinski, Z. and Podhalanska, T. 2010. Outline of the lithology and depositional features of the Lower Paleozoic strata in the Polish part of the Baltic region. Geological Quarterly, 54:109-121.
Mõtus, M.-A. 1997. Tabulate corals, p. 219-223. In Raukas, A. and Teedumäe, A. (eds.), Geology and Mineral Resources of Estonia. Tallinn, Estonian Academy Publishers.
Mõtus, M.-A. and Vinn, O. 2009. The worm endosymbionts in tabulate corals from the Silurian of Podolia, Ukraine. Estonian Journal of Earth Sciences, 58:185-192. https://doi.org/10.3176/earth.2009.3.03
Nestor, H. 1966. Stromatoporoidei Venloka i Ludlova Estonii, Valgus, Tallinn, 1-87. (In Russian)
Nestor, H. 1997. Stromatoporoids, p. 215-219. In Raukas, A. and Teedumäe, A. (eds.), Geology and Mineral Resources of Estonia. Estonian Academy Publishers, Tallinn.
Nikiforova, O.I. 1954. The Stratigraphy and brachiopods of the Silurian deposits from Podolia. Gostoptekhizdat, Moscow, 1-178. (In Russian)
Nikonov, N.I., 2000. Timan-Pechora sedimentary basin. Atlas of Geological Maps. Regionalinyi Dom Pechati. (In Russian)
Palmer, T.J. and Wilson, M.A., 1988. Parasitism of Ordovician bryozoans and the origin of pseudoborings. Palaeontology, 31:939-949.
Pătruț, I. 1981. Le Silurien de la Plate-forme Moldave et sa position dans le cadre geologique regional. Lucrările Sesiunii Științifice Grigore Cobălcescu 2:181-190.
Robin, N. 2021. Importance of data on fossil symbioses for parasite-host evolution, p. 51-73. In De Baets, K.D. and Huntely, J.W. (eds.), The Evolution and Fossil Record of Parasitism. Springer, Cham. https://doi.org/10.1007/978-3-030-52233-9_2
Sokolov, B.S. 1955. Tabulata of Paleozoic European part of USSR. Trudy VNIGRI, Novaya seria, vyp, 85:3-527. (In Russian)
Sytova, V.A. 1966. The bioherms of the Malinuvtsy horizon (Silurian) of Podolia. Bulletin of Moscow Society of Explorers of Nature, Department of Geology, 41:130-131. (In Russian)
Stur, D. 1872. Der westliche Theil der Aufnahmgebietes am Dniester in Galizien und Bukowina in der Umgebungen von Zaleszcyki. Abhandlungen der Kaiserlich-Königlichen Geologischen Reichsanstalt (Österreich), 3:1-124.
Tapanila, L. 2005. Palaeoecology and diversity of endosymbionts in Palaeozoic marine invertebrates: trace fossil evidence. Lethaia, 38:89-99. https://doi.org/10.1080/00241160510013123
Tapanila, L. 2008. Direct evidence of ancient symbiosis using trace fossils, p. 271-287. In Kelley, P.H. and Bambach, R.K. (eds.), From Evolution to Geobiology: Research Questions Driving Paleontology at the Start of a New Century: Paleontological Society Short Course, October 4, 2008. Paleontological Society, Washington. https://doi.org/10.1017/S1089332600001728
Taylor, P.D. 2015. Differentiating parasitism and other interactions in fossilized colonial organisms. Advances in Parasitology, 90:329-347. https://doi.org/10.1016/bs.apar.2015.05.002
Tsegelnjuk, P.D. 1969. Stratigraphy and age of Silurian and Lower Devonian deposits Pridniestrovia. Geological journal, 29(4):59-66. (In Russian)
Tsegelnjuk, P.D. 1980a. The Jarug and Malinovits Series (Lower - Upper Silurian) of Podillya and Volyn. Preprint 80-2. Institute of Geological Sciences USSR, Kiev, 1-54. (In Russian)
Tsegelnjuk, P.D. 1980b. The Rukshkin and Tzygan Series (Upper Silurian - Lower Devonian) of Podillya and Volyn. Preprint 80-11. Institute of Geological Sciences USSR, Kiev, 3:1-55. (In Russian)
Tsegelnjuk, P.D. and Bukatchuk, P.D. 1974. The southwestern slope of the Ukrainian Shield. Stratigraphy of the Ukrainian SSR, Silurian. Naukova Dumka, 4:156-173. (In Ukrainian)
Tsegelnyuk, P.D., Grytsenko, V.P., Konstantinenko, L.I., Ishchenko, A.A., Abushik, A.F., Bogoyavlenskaya, O.V., Drygant, D.M., Zaika-Novatsky, V.S, Kadlets, N.M., Kiselev, G.N. and Sytova, V.A. 1983. The Silurian of Podolia. The guide to excursion. Kiev, Naukova dumka. (In English and Russian)
Văscăuţanu, T. 1931. Les formations siluriennes de la rive Roumaine du Dnister. Extras din Anuarul Institutului Geologic al României, l:425-546.
Venyukov, P.N. 1899. The fauna of Silurian deposits of Podillyan province. Russian Academy of Sciences, St. Petersburg. (In Russian)
Vinn, O. and Mõtus, M.-A. 2014a. Endobiotic rugosan symbionts in stromatoporoids from the Sheinwoodian (Silurian) of Baltica. PLoS ONE, 9:e90197. https://doi.org/10.1371/journal.pone.0090197
Vinn, O. and Mõtus, M.-A. 2014b. Symbiotic worms in biostromal stromatoporoids from the Ludfordian (Late Silurian) of Saaremaa, Estonia. GFF, 136:503-506. https://doi.org/10.1080/11035897.2014.896412
Vinn, O. and Wilson, M.A. 2010a. Endosymbiotic Cornulites in the Sheinwoodian (Early Silurian) stromatoporoids of Saaremaa, Estonia. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 257:13-22. https://doi.org/10.1127/0077-7749/2010/0048
Vinn, O. and Wilson, M.A. 2010b. Microconchid-dominated hardground association from the late Pridoli (Silurian) of Saaremaa, Estonia. Palaeontologia Electronica, 13(2):9A. https://palaeo-electronica.org/2010_2/212/index.html
Vinn, O. and Wilson, M.A. 2016. Symbiotic interactions in the Silurian of Baltica. Lethaia, 49:413-420. https://doi.org/10.1111/let.12158
Zaika, Yu, 2007. Ordovician and Silurian Tabulatomorphic corals of the Russian Arctic. Thesis for: Candidate of Geological and Mineralalogical Sciences, St. Petersburg, VSEGEI, 1-21. (In Russian)
