

 A survey of Pliocene to Mid-Quaternary leaf cuticle from the North Island, New Zealand
A survey of Pliocene to Mid-Quaternary leaf cuticle from the North Island, New Zealand
Article number: 22.1.12
https://doi.org/10.26879/862
Copyright Paleontological Society, March 2019
Author biography
Plain-language and multi-lingual abstracts
PDF version
Submission: 27 February 2018. Acceptance: 7 February 2019
{flike id=2437}
ABSTRACT
This study reports on a survey of dispersed leaf cuticle from a range of late Pliocene to mid-Pleistocene (mostly coastal) lignites in the North Island of New Zealand. With few exceptions, the samples contain plant taxa common in New Zealand today, for example, Beilschmedia (Lauraceae) and numerous conifers. However, some of them are outside of their current latitudinal-altitudinal range. The common presence of Halocarpus (Podocarpaceae) is noteworthy, as almost all records from the North Island today are from upland regions. There is no clear evidence of Agathis (Araucariaceae) having existed further south than its present limit. However, a single find of an araucarian in the Rangitawa Fossil Beds in the southern North Island is notable. Its identity is not clear, but it is possibly Araucaria (Araucariaceae), making it the youngest occurrence of this genus in New Zealand. If it is Agathis, then it is the most southern Pleistocene macro-record. Cryptocarya (Lauraceae) is recorded in an undated peat at Huriwai. It represents an apparent Pleistocene survival of a genus known from the Miocene in New Zealand, but now locally extinct. Carmichaelia (Fabaceae) cuticle was common, but unexpected, as it is very rare in the palynological record.
Mike Pole. Queensland Herbarium, Brisbane Botanic Gardens Mt Coot-tha, Toowong, Qld, Australia. murihiku@yahoo.com
Keywords: epidermis; Lauraceae; Fabaceae; Araucariaceae; Podocarpaceae
Pole, Mike. 2019. A survey of Pliocene to Mid-Quaternary leaf cuticle from the North Island, New Zealand. Palaeontologia Electronica 22.1.12A 1-32. https://doi.org/10.26879/862
palaeo-electronica.org/content/2019/2437-pliocene-quaternary-cuticle
Copyright: March 2019 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
Each global glacial-interglacial cycle is effectively a naturally repeating experiment. Cold or dry conditions, or even the direct physical effects of glacial activity, substantially alter the vegetation and soil conditions. In otherwise forested regions, the trees return after each interglacial warming, but due to glacial period extinction or simple chance - the make-up of the forests may differ between interglacials. Vegetation from past interglacial periods is therefore of interest as analogues for the Holocene and possible future. However, as atmospheric carbon dioxide levels are now higher than their peaks throughout the Pleistocene, their efficacy as analogues becomes more doubtful. In that respect, earlier analogues must be sought, before 2.1 Ma, or even before 2.7 Ma--the Pliocene (Hönisch et al., 2009).
The sequence of vegetation change which occurred in New Zealand from the Last Glacial Maximum through the Holocene to the present is now understood in reasonable amount of detail. This has mostly been due to a concerted effort by palynologists (e.g., McGlone, 1983, 1988; McGlone et al., 2003) and stratigraphers. Now there is an increasing focus to extend this understanding back through previous Glacial-Interglacial cycles (e.g., Bussell, 1986, 1992; Bussell and Pillans, 1992; Moar and Suggate, 1996; Newnham et al., 1999; Moar and McKellar, 2001). Over this time scale, not only have there been major rearrangements of vegetation communities with respect to relative dominance, but there have been both extinctions, and a few natural introductions to New Zealand as well (e.g., Bussell and Mildenhall, 1990; Kershaw et al., 2000). A detailed knowledge of the timing of these events becomes more important in this time range, as other methods of dating become harder.
There have been a few Quaternary macrofossil studies in New Zealand, which complement the palynological work, for instance McQueen (1954), Couper and McQueen (1954), Newnham and Lusk (1987), and Ogden et al. (1993). However, none of these macrofossil studies have involved studying leaf cuticle. Cuticle is useful because small fragments (i.e., 1mm sq.) have the potential for greater taxonomic precision than using pollen or spores. They may have a typical resolution to the genus or species level, whereas pollen and spores are more typically to the genus or family level. Cuticle may therefore potentially identify extinct species. Several of the Pleistocene sites in the North Island have had pollen of Acacia reported (Mildenhall, 1975a, b; Bussell, 1992), a genus that is not now indigenous to New Zealand. If leaf cuticle of Acacia was present, it would allow both confirmation of the palynological record and an indication of the extant taxonomic affinities of the fossil within the genus. Cuticle may also identify taxa, which are usually invisible in the palynological record, the ‘classic’ case being Lauraceae, where cuticle may not only identify the family, but the genus as well. Cuticle is also more likely to record local vegetation rather than the regional, wind-dispersed pollen that is thought to dominate many palynological samples. Dispersed cuticle may also be present when there are no obvious signs of macrofossils. A relatively small sample (c. 100-300 g) may be sufficient to include many species of plant. The drawback of cuticle is that it is preserved much more infrequently than pollen, and for practical reasons, sample size (at least for a reconnaissance-level project such as this) is larger and temporal resolution lower than for palynology.
Lignite (or somewhat indurated peat) deposits of Late Pliocene-Early Pleistocene age are preserved in various localities in the North Island of New Zealand, mostly around the coast. In the north west of the North Island many of these isolated occurrences have been collectively called the Kaihu Group (Chappell, 1970) and the palynology has been documented in one of the largest palynological studies in New Zealand (Mildenhall et al., 1992) which incorporated several earlier works (e.g (Pocknall and Waterhouse, 1984; Mildenhall, 1985). Further to the south, the sediments of the Wanganui Basin have achieved a global importance as a detailed record of global sea level change over 2.6 Ma (Fleming, 1953; Pillans, 1994; Naish and Kamp, 1995; Carter and Naish, 1998a; Kamp et al., 2004; Pillans, 2017). The sediments are largely marine but include several lignite/peat horizons (Fleming, 1953). Previous macrofossil investigation of these peats includes that of McQueen (1954), Couper and McQueen (1954), and Newnham and Lusk (1987). The former paper described an extinct species of Proteaceae fruit, Proteoides ohukaensis from the mouth of Ohuka Creek. The third paper compared macrofossils with the palynological spectra.
The aim of this work is to investigate some of these lignites and associated muddy sediments for plant macrofossils, which have cuticle preservation. The initial goal, which stimulated this project, was to try and locate cuticle belonging to the plant that produced the Acacia pollen reported by Mildenhall (1975a, b) and Bussell (1992). The paper offers a photographic record of the cuticle of a range of extant New Zealand taxa that will be a useful reference for further work.
MATERIAL AND METHODS
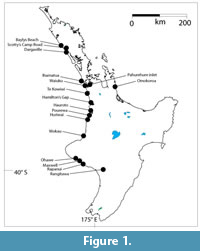 The study focussed on sites known to be Pliocene or earlier Pleistocene in age (Figure 1). Dating in most cases is broad and based on a mixture of palynology, inferred sea-level, and direct dating of volcanigenic sediment or carbonaceous material.
The study focussed on sites known to be Pliocene or earlier Pleistocene in age (Figure 1). Dating in most cases is broad and based on a mixture of palynology, inferred sea-level, and direct dating of volcanigenic sediment or carbonaceous material.
Division of Pliocene-Pleistocene time in New Zealand (Figure 2) follows Raine et al. (2015), which largely follows Beu (2001). This does not recognise several terms that occur frequently in the earlier palynological literature and therefore to help the reader, some of the system of Carter and Naish (1998b) is also included.
The localities were visited over multiple trips from the mid-1990s. Sample size was about 400 g, and where there was no obvious change in lithology, they were taken across the full thickness of a carbonaceous bed (up to around 1 m in some cases). Grid references are given in New Zealand Transverse Mercator (NZTM). Appropriate New Zealand Geological Society Fossil Record Numbers given in Mildenhall et al. (1992) are also cited (e.g., ‘P7 f9583’).
 Disaggregation of samples was usually achieved by immersing the sample in hot water and adding 45 % hydrogen peroxide. If this worked the remaining sludge was then sieved through an approximately 1 mm mesh and the fines discarded. Adhering silica was removed using 70 % hydrofluoric acid. Lignite that did not disaggregate in peroxide was placed directly into c. 10 % chromium trioxide or concentrated hydrogen peroxide and warmed gently. This process reduced any leafy plant material to translucent cuticle, which could then be stained with safranin. Cuticle fragments were then mounted on either microscope slides using thymol glycerine jelly or on scanning electron microscope stubs with double-sided tape and coated with platinum. Microscope slides are catalogued with the prefix ‘SL’ or ‘LX’ and electron microscope stubs with ‘S-’. Slides were viewed under Transmitted Light Microscopy (TLM) or with Scanning Electron Microscopy (SEM). Comparative herbarium material is prefixed with OTA (Otago University Botany Department, CHR (the Allan Herbarium), and OPH (material collected by the author).
Disaggregation of samples was usually achieved by immersing the sample in hot water and adding 45 % hydrogen peroxide. If this worked the remaining sludge was then sieved through an approximately 1 mm mesh and the fines discarded. Adhering silica was removed using 70 % hydrofluoric acid. Lignite that did not disaggregate in peroxide was placed directly into c. 10 % chromium trioxide or concentrated hydrogen peroxide and warmed gently. This process reduced any leafy plant material to translucent cuticle, which could then be stained with safranin. Cuticle fragments were then mounted on either microscope slides using thymol glycerine jelly or on scanning electron microscope stubs with double-sided tape and coated with platinum. Microscope slides are catalogued with the prefix ‘SL’ or ‘LX’ and electron microscope stubs with ‘S-’. Slides were viewed under Transmitted Light Microscopy (TLM) or with Scanning Electron Microscopy (SEM). Comparative herbarium material is prefixed with OTA (Otago University Botany Department, CHR (the Allan Herbarium), and OPH (material collected by the author).
Results are discussed by location stratigraphically below, and taxonomically in the Systematic Palaeontology section.
Material is lodged in the Queensland Herbarium, Brisbane.
RESULTS
A. Kaihu Group
i. Formation undifferentiated
Scottys Camp Road (P7/f9579)
Location. Roadside cutting 0.75 km SE of intersection of State Highway 12 and Scottys Camp Road (grid reference: NZTM 1674159, 6024386).
Age. Hautawan (late Pliocene) or possibly earliest Marahauan (earliest Pleistocene) (Mildenhall, 1985).
Palynology. The pollen assemblage is dominated by Myrtaceae (incl. Eucalyptus ) and Acacia pollen is present (Mildenhall, 1985).
Macroflora (present study). Includes Dacrycarpus, Dacrydium, Beilschmiedia, Carmichaelia.
Dargaville (P9/f9582)
Location. Roadside cutting 1.5 km NW of Dargaville (grid reference: NZTM 1676256, 6025190.
Age. Hautawan (late Pliocene) or possibly earliest Marahauan (earliest Pleistocene) (Mildenhall, 1985).
Palynology. The pollen assemblage is dominated by Myrtaceae (incl. Eucalyptus ) and Nothofagus s.g. Fuscospora. Acacia pollen is abundant (Mildenhall, 1985; Mildenhall et al., 1992).
Macroflora (previous work). Beilschmiedia taraire leaves were reported by J. Braggins (personal commun., to Mildenhall, 1985).
Macroflora (present study). The dominant plant to be an unidentified monocot. Remains of other taxa are uncommon, but include Dacrycarpus dacrydioides, Beilschmiedia, and Carmichaelia.
Baylys Beach (P7 f9583)
Location. Coastal cliffs at the end of Baylys Coast road (grid reference: NZTM 1667272, 6020671.
Age. Possibly early Pleistocene (Richardson, 1985).
Palynology. A palynological sample 6.4 km to the NW (P7/f9581) is dominated by Nothofagus “fusca” group and Dacrydium cupressinum. No extinct pollen taxa were found, and it was regarded as Castlecliffiian or younger (Mildenhall, 1985).
Macroflora (previous work). C.J. Burrows (personal commun., to Richardson, 1985, p. 118) reported that P7/f9583 “Contains seeds of an apparently extinct plant.” C.J. Burrows and J. Braggins (personal commun., to Mildenhall, 1985) reported that P7/f9581 contained leaves of Dacrydium cupressinum, Podocarpus totara, Agathis, Cyathodes, and Nothofagus truncata, along with wood of Dacrydium colensoi, Agathis and Myrtaceae, and seeds of Elaeocarpus dentatus, Eleocharis sphacelate, and D. cupressinum.
Macroflora (present study). Overwhelmingly dominated by Agathis australis leaves and cone scales, with Dacrydium cupressinum also present.
ii. Ohuka Formation
Pourewa (R15/f73, 74)
Location. Exposed at the edge of a beach at the high tide level, Pourewa Point, Aotea (grid reference: NZTM 1760484, 5791809).
Age. Hautawan-Marahauan (Late Pliocene - Early Pleistocene). Chappell (1970) referred to this as Pourewa Lignite, a term that Mildenhall et al. (1992) recommended be given member status of the Ohuka Formation.
Palynology. Pocknall and Waterhouse (1984) noted a dominance of Nothofagus s.g. fuscospora pollen and that there were no extinct species and correlated the Pourewa Lignite with the Te Kowiwi Lignite. Mildenhall et al. (1992) recorded similar palynological assemblages but also noted the extinct spore Polypodiisporites radiatus Pocknall and Mildenhall (1984).
Macroflora (present study). No stomatiferous leaf cuticle was found.
Te Kowiwi (R15/f83, 84)
Location. Shore exposure, west side of Kowiwi Bay, directly below power lines (grid reference: NZTM 1760185, 5790908.
Age. Mildenhall et al. (1992) regard this assemblage as Hautawan (late Pliocene). Pocknall and Waterhouse (1984) referred to this as Te Kowiwi Lignite, a term that Mildenhall et al. (1992) recommended be given member status of the Ohuka Formation.
Palynology. Pocknall and Waterhouse (1984) recorded the dominant pollen taxa as Nothofagus s.g. Fuscospora, Dacrydium, Phyllocladus, Prumnopitys, and Libocedrus. The extinct taxon Polypodiisporites radiatus Pocknall and Mildenhall was also recorded. This has a known upper limit of basal Nukumaruan (The base of the Nukumaruan is about 2.5 Ma, Pliocene; Beu, 2001). Mildenhall et al. (1992) recorded similar assemblages with one including a large fraction of Dacrycarpus.
Macroflora (present study). This study found the macroflora to be overwhelmingly dominated by small and imbricate-leaved conifer shoots, predominantly of Halocarpus, and also Dacrycarpus and Libocedrus. Only two angiosperm taxa are present, Phormium and Pseudowintera. The macroflora suggests the original vegetation was a conifer-dominated swamp. However, the conifer leaves and shoots were mostly very twisted and distorted, perhaps indicating some disturbance before final burial.
Waiuku (R12/f45, 46)
Location. An exposure at water level in an inlet at the southern end of Manukau Harbour at the end of Racecourse Road, Waiuku (grid reference: NZTM 1753646, 5877698.
Age. Marahauan-lower Haweran; Late Pliocene-late Pleistocene (Mildenhall et al., 1992; See also Byrami et al., 2005).
Palynology. Mildenhall et al. (1992) placed this exposure in the Ohuka Formation, noting a dominance of Podocarpaceae pollen, but including no extinct species.
Macroflora (present study). The cuticle assemblage consists entirely of Beilschmiedia.
Ihumatao
Location. Peaty material with abundant wood and stumps is exposed by wave action on a coastal platform, (grid reference: NZTM 1755940, 5903759).
Age. A range of dates have been obtained by several researchers, including from radiocarbon from c. 29,000 to c. 43,600 years BP (Ferguson and Rafter, 1959; Grant-Taylor and Rafter, 1963; Polach et al., 1969), K-Ar dates on basaltic material of c. 74,000 (McDougall et al., 1969) and 38,000 years BP using thermoluminescence (Wood, 1991). Following this uncertainty, a focussed study including Optically Stimulated Luminescence dating by Marra et al. (2006) regarded the forest as much older‒being deposited “during late MIS 7 (probably substage 7.1)” (c. 190,000-210,000 BP). Ricketts (1977) described this as the “Ihumatao Fossil Forest” and concluded it had been buried by volcanic tuff (See also Hayward,1977). Hayward and Hayward (1995) recognised two forests, one growing within peat and a further one on the peat. They mapped the location of in situ stumps and larger fallen logs. The forest was subsequently investigated for insects and palynology (Marra et al., 2006).
Macroflora (previous work). Ricketts (1977) noted Agathis and cf. Dacrydium cupressinum. Hayward and Hayward (1995) listed Dacrydium cupressinum, Elaeocarpus dentatus, Prumnopitys ferrugineus, Agathis australis, Phyllocladus trichomanoides, and Dianella. Marra et al. (2006) illustrated in situ Dacrydium.
Macroflora (present study). Found the peat around the lower forest to be dominated by Agathis australis, but also with Halocarpus, Beilschmiedia, Pseudowintera, and Toronia.
Hamiltons Gap
Location. Beds of highly compressed leaves and other carbonaceous matter occur in high cliffs to the south of Hamiltons Gap, (grid reference: from NZTM 1739935, 5888430 to 1739958, 5888394).
Age. The Hamiltons Gap material appears to be similar to that reported by Newnham and Lusk (1987) from about 15 km to the south and 1.5 km north of Karioitahi Beach, (grid reference> R12 564 357).
Macroflora (previous work). Newnham and Lusk’s (1987) macrofossil material included Laurelia novae-zelandiae, Alectryon excelsus, Mysine salicina, Beilschmiedia taraire, B. tawa, Macropiper excelsum, Elaeocarpus sp., Cyathea sp., Griselinia sp., Podocarpus sp. (identified in the field only), and indeterminate monocot leaves. They noted the abundance of Podocarpaceae and Myrtaceae pollen, although corresponding macrofossils were, consecutively, rare and absent.
Macroflora (present study). The material is distinctly lithified and does not disaggregate well with peroxide treatment. However, chromium trioxide treatment showed fragmented cuticle was abundant and well-preserved, and includes Grisselinia, Olearia, and Astelia.
Huriwai
Location. Peat in coastal cliff (grid reference: NZTM 1751200, 5854900).
Age. Unknown.
Macroflora (present study). The cuticle assemblage consists entirely of Pseudowintera, Beilschmiedia, and Cryptocarya.
Hauroto Bay
Location. Coastal exposure (grid reference NZTM 1771840, 5814030).
Age. Hautawan (Mildenhall, 1992).
Macroflora (present study). The cuticle assemblage consists mostly of Beilschmiedia.
Pahurehure
Location. Peaty material is exposed in the low cliff bordering Pahurehure Inlet a few meters west of the Auckland Motorway. (grid reference NZTM 1770980, 5896670).
Age. Unknown.
Macroflora (present study). Contains Dacrycarpus, Dacrydium, Libocedrus, Beilschmiedia, and the two extant New Zealand Proteaceae; Toronia toru and Knightia excelsa.
Ohawe
Location. Bussell (1990) recognised two broad localities: ‘Ohawe Waterfall’ samples came from within 24 below the Stratford Lahar and fell into his OH4 zone, and ‘Ohawe East’ samples came from just above the Stratford Lahar and fell into his OH5 zone. The present study found one productive sample from the ‘Ohawe Waterfall’ locality (my sample Ohawe-4, from lignite directly below the Stratford Lahar, grid reference: NZTM 1704360, 5616740) and two samples from ‘Ohawe East’ (my sample Ohawe-5, from the lower main lignite, and, separated by a c. 30 cm overlying tuff, sample Ohawe-6, from the upper main lignite, grid reference NZTM 1704700, 5616710).
Wards (1996) studied a section c. 1640 m northwest of Bussel’s (1990) ‘Ohawe Waterfall’ locality. Fallen blocks from here were sampled and come from approximately 35-36 m on Wards’ (1996, Ohawe Waterfall section (my sample Ohawe-2, grid reference: NZTM 1702870, 5617400).
Age. Bussell (1990) dated the waterfall section as Oxygen Isotope Stage 6 (c. 130,000-190,000 years BP, full glacial) and Ohawe east as OI5e (c. 110,000-130,000 years BP, full interglacial), reported by Bussell and Mildenhall (1990).
Palynology. Bussell (1990) interpreted the Water Fall samples as representing a rush-fern shrubland with sedges and common Phormium, but with patches of shrubland including Halocarpus. In contrast, he considered that Ohawe East samples probably represented forest with prominent Dacrycarpus and Syzygium with wetland taxa in patches.
Macroflora (present study). Ohawe-4 contains Phormium and Carmichaelia, Ohawe-5 contains Cyperaceae and Ohawe-6 contains Halocarpus.
Rapanui
Location. Exposure in coastal cliff (grid reference: NZTM 1764460, 5581563).
Age. Bussel (1992), with reference to work of Pillans (1983, 1988, 1990) estimated the age of the Rapanui Lignite as c. 110,000 years BP.
Palynology. Bussel (1992) interpreted as “low forest/shrubland with prominent Myrtaceae, and including a now-extinct Acacia -type.”
Macroflora (present study). Dominated by Halocarpus and Dacrydium with some Dacrycarpus and Prumnopitys.
Omokoroa
Location. (grid reference NZTM 1869491, 5831249).
Age. Briggs et al. (2005). Note the Te Puna Ignimbrite (0.93 Ma) overlies lignites at Omokoroa.
Macroflora (present study). Contains Halocarpus and Phyllocladus.
B. Wanganui Basin
i. Maxwell Group
The basic stratigraphy of the Wanganui Basin was documented by Fleming (1953), who described many marine units, but amongst these were three formations, which he described as containing ‘lignite’ - the Upper Maxwell, Middle Maxwell, and Lower Maxwell Formations. These crop out in the coastal cliffs east and west of the mouth of Ototaka Ck, south of Maxwell. The palynology of these three units was documented by W.F. Harris and R.A. Couper (in Fleming, 1953). Some major quantitative differences were noted between samples, but no extinct taxa were recorded.
Age. The Maxwell Formations were deposited from just before the Plio-Pleistocene boundary (1.81 Ma) and approximately 1.6 Ma (Marahauan, early Pleistocene; Carter and Naish, 1998b).
Palynological samples were documented by W.F. Harris and R.A. Couper (personal commun., in Fleming, 1953).
Upper Maxwell Formation
Location. From 2100 m southeast of the Ototoka Stream mouth (sample Maxwell-13, grid reference: NZTM 1758686, 5584805).
Palynology. Harris and Couper’s (in Fleming, 1953) palynological analysis of two ‘lignite’ samples within the Formation suggested Nothofagus - Weinmannia forest, and podocarp-dicotyledonous forest. No extinct taxa were noted.
Macroflora (present study). No good cuticle.
Middle Maxwell Formation
Location. From 360-660 m southeast of the Ototoka Stream mouth (sample Maxwell-07 grid reference NZTM 1757000, 5585360, and Maxwell-09 grid reference: NZTM 1757279, 5585261).
Palynology. Harris and Couper’s (in Fleming, 1953) palynological analysis of a ‘lignite’ bed suggested a Weinmannia - Nothofagus forest. No extinct taxa were noted.
Macroflora (present study). No good cuticle.
Lower Maxwell Formation
Location. From 830 m west of the Ototoka Stream mouth (sample Maxwell-17, grid reference: NZTM 1755840, 5585629).
Palynology. Harris and Couper’s palynological analysis of three ‘lignite’ beds included within the Lower Maxwell Formation noted three distinct assemblages which they identified with Nothofagus - Weinmannia forest, Podocarp-dicotyledonous forest, and Metrosideros -podocarp forest. No extinct taxa were noted.
Macroflora (present study). Contains sparse Dacrydium, Dacrycarpus and Cyperaceae.
Rangitawa Fossil Beds
Location. The Rangitawa Fossil Beds (RFB) was first described by Te Punga (1952, 1962) and more detailed work followed by McQueen (1953), Bussell (1986), and Kohn et al. (1992). For the present study unambiguous RFB was sampled (samples Rangitawa-7, and Rangitawa-11) from outcrop close to normal flow level of the Rangitawa Stream (thus low in the local section which has a very gentle-dip). The peaty material also contains molluscs, as per original descriptions of the RFB, but may be 100 m downstream of where Bussell (1986) indicated samples on his map, (grid reference: NZTM 1809596, 5554665).
Age. Kohn et al. (1992) suggest the Rangitawa Fossil Beds (RFB) date to a little after the start of OIS 11, an interglacial, making them around 420, 000 yrs BP. They date the overlying Rangitawa Pumice to about 350,000 yrs BP, which correlates with OIS 10, a glacial period.
Palynology. Bussell (1986) described the palynology of the RFB, and interpreted a “warm-temperate paleoclimate” under “lowland podocarp-hardwood-beech forest.”
Macroflora (previous work). Leaf and seed fossils were described from the Rangitawa Stream by McQueen (1953, 1954) and indicate forested conditions. Two extinct seed species were described; Podocarpites tolleyi (Podocarpaceae) and Rumex pachyperianthus (Polygonaceae).
Macroflora (present study). Contains Carmichaelia and Beilschmiedia.
Rangitawa Pumice
Location. Roots, logs and plant macrofossils were reported by Bussell (1986) in mudstone associated with the Rangitawa Pumice about 17 m above the RFB and about 70 m downstream of his RFB samples. In the present study, what is regarded as the same horizon was located, exposed in a cliff, about 80 m upstream of Bussell’s RFB. The horizon is inaccessible, and therefore fossil material was collected from fallen blocks directly below the cliff (samples Rangitawa-8, -9, -10, Grid reference: NZTM 1809761, 5554759).
Age. Kohn et al. (1992) date the Rangitawa Pumice to about 350,000 yrs BP, which correlates with OIS 10, a glacial period.
Palynology. The palynology of the sediments associated with the Rangitawa Pumice was documented by Bussell (1986) and later by Kohn et al. (1992, as their ‘Pole Site 2’). These results indicated open, non-forested conditions of grassland-shrubland, similar to a “Present day subalpine environment” (Bussell, 1986).
Macroflora (present study). Contains Beilschmiedia and Carmichaelia.
Rangitawa ‘Upstream’
Location. In situ peaty material was collected near water level in the left bank of the Rangitawa River (sample Rangitawa-6, grid reference NZTM 1809990, 5554740). This is 300‒400 m upstream and along strike of known RFB outcrop and is a likely correlate.
Macroflora (present study). Contains the conifers Halocarpus, Libocedrus, Dacrydium, Phyllocladus, Prumnopitys, and the angiosperms Carmichaelia and Pseudowintera.
Rangitawa ‘Float’
Location. Two blocks of ‘float’ fossiliferous mudstone were collected from the Rangitawa stream bed downstream of Rangitawa Pumice outcrop and of probable RFB outcrop. It is uncertain what horizon they derive from (samples Rangitawa-4 and Rangitawa-5, grid reference NZTM 1809750, 5554700).
Macroflora (present study). Contains the conifers Araucaria or Agathis, Libocedrus, Phyllocladus, and angiosperms Carmichaelia Beilschmiedia, and Pseudowintera.
C. Uncertain unit
Mokau
Location. In 1996 a collection was made from a c, 2 m thick carbonaceous mud horizon exposed in a fresh road-cutting of State Highway c. 3 km south of the bridge over the Mokau River. A return visit to clarify the stratigraphy in 2005 could not relocate it, as it appears to have been covered by a small slump, (grid reference: NZTM 1740577, 5712664).
Stratigraphy and age. Henderson and Ongley (1923, p.54) mentioned Early Pleistocene “soft sandy silt containing plant-remains”... “on the coast half a mile south of the Mokau River”. It is possible that this refers to similar material, although the locality collected here is inland.
Macroflora (present study). Contains Dacrycarpus, Halocarpus and an indet. conifer, and the angiosperms Carmichaelia, Griselinia, Litsea. Pseudowintera, and Phormium.
DISCUSSION
This survey demonstrates the existence of well-preserved leaf cuticle in some of the Plio-Pleistocene peat and associated sediments in the North Island (Figure 3). Both angiosperm and conifer cuticle is often abundant. Due to the sometimes higher taxonomic resolution of cuticle compared with pollen, this type of evidence makes it a useful contribution to the field.
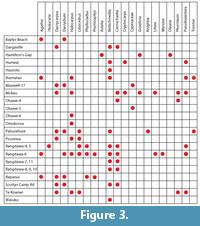 The presence of a now-extinct taxon of Lauraceae, Cryptocarya, and perhaps Araucaria, in the Quaternary, is notable. However, these extinct taxa are rare finds. Overall the taxa recovered as cuticle indicate interglacial vegetation that was little different from the swamp-forest (with common conifers) and associated lower vegetation (Phormium-dominated) that grew around the coast of the North Island in Holocene times. The cuticle evidence allows the identification of Beilschmedia, and sometimes Litsea, both locally common components of North Island forest today (with Beilschmedia extending to the north-east of the South Island; (Wardle, 1991), but essentially absent from the palynological record.
The presence of a now-extinct taxon of Lauraceae, Cryptocarya, and perhaps Araucaria, in the Quaternary, is notable. However, these extinct taxa are rare finds. Overall the taxa recovered as cuticle indicate interglacial vegetation that was little different from the swamp-forest (with common conifers) and associated lower vegetation (Phormium-dominated) that grew around the coast of the North Island in Holocene times. The cuticle evidence allows the identification of Beilschmedia, and sometimes Litsea, both locally common components of North Island forest today (with Beilschmedia extending to the north-east of the South Island; (Wardle, 1991), but essentially absent from the palynological record.
The repeated occurrence of the conifers Halocarpus and Libocedrus confirms palynological evidence for these taxa. The occurrences are significant as they are outside of their current geographical range. Both of these taxa have restricted, upland distributions in the North Island today, although they occur at sea level much further to the south, in the South Island. Libocedrus and Halocarpus are, for example, important conifers today on Mount Hauhungatahi above 1000-1150 m (Atkinson, 1981; Druitt et al., 1990), but Halocarpus especially, is almost never below this level in the central North Island. The occurrence of both taxa at present sea level in the North Island suggests cooler conditions.
With the single exception of a possible fragment from Rangitawa, Agathis cannot be confirmed in any of the Plio-Pleistocene samples south of this range (i.e., southern North Island samples). If the Rangitawa Araucariaceae is Araucaria, then it is the youngest known occurrence in New Zealand. If it is Agathis, then it is far south of its current southern limit of 38.11° S (Ecroyd, 1982) or its earlier Holocene range (Ogden et al., 1992). The general absence of Agathis macrofossils south of its current range suggests temperatures may not have been warmer than the present day for those periods. Agathis has been reported further south in some interglacials (e.g., Mildenhall, 1986) but based on pollen, which is difficult or impossible to distinguish from Araucaria. In this context the possible record of Araucaria cuticle from Rangitawa is significant.
The coexistence of both of New Zealand’s extant Proteaceae species, Knightia excelsa and Toronia toru, at Pahurehure is unusual, but this combination does occur in various regions of the North Island today. K. excelsa is a very widespread forest tree of the North Island (and extends a little into the South Island) while T. toru is more restricted. Its southern limit is the Bay of Plenty, but it occurs from lowland to mountain forest (Dawson and Lucas, 2000). It is common in Agathis australis forest at Waipoua, Northland (McGregor, 1948, p.22), where it is described as a “distinctively northern tree”. Grove and Brandon (2006) reported both Proteaceae species in the foothills of the Coromandel Range, near Tairua. They specifically reported T. toru only from the upper slopes and ridge top of the generally secondary vegetation.
The common presence of Libocedrus and Winteraceae amongst the plant remains reinforces the findings of Pole (2007a) in Pliocene localities in the South Island. However, other taxa are noticeable by their general absence or rarity in this survey. Among the conifers for instance, Phyllocladus might be expected, given its importance today in relatively extreme environmental situations, but it was found only in two localities (Rangitawa and Rapanui). There are few records of Phyllocladus growing near sea level in the Auckland region today, although it is common in the uplands. However, all the extant sea-level records appear to be Phyllocladus trichomanoides, while it is unclear to which species the fossils belong to.
The widespread presence of Carmichaelia in these samples was surprising. McGlone et al. (1984) recorded a trace of Carmichaelia pollen in the late Pleistocene of the Inaha Terrace (near the Ohawe site), but in general the genus appears to be severely under-represented in the palynological record of these peats. However, the genus is known from swamps, even tidal swamps (Heads, 2017), and one species, Carmichaelia arborea, is known as ‘Swamp broom’. The cuticle comes from cladodes (it was initially suspected as the source plant of the Acacia pollen, another cladode-bearing plant), but despite not being shed like normal leaves, the remains clearly find their way into the fossil record with some ease. In this respect, the absence of Carmichaelia from any of the diverse early Miocene cuticle assemblages is notable (personal observ.).
The macrofossils found by Ogden et al. (1993) make an interesting comparison to those reported here. For example, the Lauraceae that are virtually ubiquitous in the present samples, were absent from theirs. Conversely, their material included Phyllocladus, although it is uncommon in samples studied here.
There is a clear discrepancy between Bussell’s (1986) palynological results of the Rangitawa Pumice, which indicated grassland-shrubland and the “logs, and plant macrofossils” indicated on his figure 2. The Beilschmiedia cuticle recovered in this work suggests forest, consistent with the logs. This is difficult to reconcile, but might, for example, result from long-distance river transport of forest debris from an isolated patch of forest in sheltered conditions upstream. However, for most of the cuticle in this study, its presence in peat, or at least very carbonaceous muds, suggests little fluvial transport, which would dilute the organic fraction with mud or sand. On that basis, the cuticle assemblages are inferred to represent local vegetation (Ferguson, 1985; Greenwood, 1991; Spicer, 1991; Gastaldo and Staub, 1999).
The broad composition of these samples - composed mainly of extant taxa plus a few, rare taxa destined to become extinct sometime within the Pleistocene, supports the conclusions of palynology (Bussell and Mildenhall, 1990).
As fossil cuticle samples the present material is of relatively low diversity. The most diverse sample was Rangitawa-6, with 11 taxa. Typically there are fewer taxa. In contrast, early Miocene dispersed cuticle assemblages from New Zealand typically contain more, and sometimes over 20 species (e.g., Pole, 2008a). Furthermore, Plio-Pleistocene samples typically have 1-2 species of Lauraceae, comparing well with present-day forests in the northern half of New Zealand, but significantly less than early Miocene where samples, which often have more than 10 Lauraceae species (Pole, 2007b). Both these observations confirm a relatively low diversity of the Plio-Pleistocene source vegetation compared with early Miocene and is consistent with some aspect of temperature being generally cooler than then.
SYSTEMATIC PALAEONTOLOGY
Illustrations of selected specimens of all taxa found in this survey are provided here. For most of these taxa a range of illustrations have been published previously and therefore in these cases only basic TLM are given here.
Conifers
Araucariaceae
Agathis australis
Figure 4.1-2
Specimens. SL4401, Baylys Beach; SL5882, Ihamatao.
Identification. Agathis litter (as leaves and cone scales) is common at Baylys Beach and Ihamatao. Under TLM the stomatal complexes show the typical ‘spoked’ structure of the genus (Pole, 2008b).
?Araucaria
Figure 4.3-4
Specimens. SL4641, Rangitawa-5.
Identification. A single cuticle fragment from Rangitawa shows araucarian stomatal structure with oblique orientation. However, the ‘spoked’ structure of the stomatal complexes under TLM is not apparent, and it is possible that this is an Araucaria rather than an Agathis.
Cupressaceae
Libocedrus sp.
Figure 5
Specimens. SL4395, Te Kowiwi; SL4503, Pahurehure; SL4648, Rangitawa-6.
Identification. Monocylic stomates and scattered papillae and scale-like leaves indicate Libocedrus. However, the two extant New Zealand species, L. plumosa and L. bidwilli are difficult to distinguish without whole shoot data. Florin and Boutelje (1954) noted that L. plumosa had papillae on normal epidermal cells, away from the stomatal bands at the base of the leaves. The present fossils display a range of papillae prominence, some appear to lack the fused papillate ring around the stomatal aperture, and one specimen is at least partially dicyclic.
Podocarpaceae
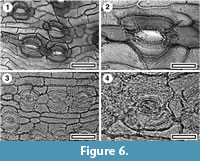
Dacrycarpus dacrydioides
Figure 6.1-2
Specimens. SL0202, Dargaville; LX2735, Maxwell-13; LX2837, Maxwell-17; SL2345, Mokau-1; SL4501, Pahurehure; SL4667, Rapanui; SL0225, Scottys Camp; SL4392, Te Kowiwi.
Identification. The very smooth and relatively thin-walled, unbuttressed epidermal cells of this genus are distinctive.
Dacrydium cuppresinum
Figure 6.3-4
Specimens. SL5972, Baylys; SL4509, Pahurehure; SL4669, Rapanui; SL5879, Ihamatao; SL2370, Max-07; SL0221, Scottys Camp; LX2836, Maxwell-17.
Identification. The cuticle morphology of Dacrydium is broadly like that of Dacrycarpus, except that the epidermal cells can be buttressed (Florin, 1931). However, the buttressing of the New Zealand Dacrydium cuppresinum can be subtle and much less than other species in this genus (see Wells and Hill, 1989). Within New Zealand, D. cuppresinum can be consistently distinguished from Dacrycarpus dacrydioides by its thicker and rougher epidermal cell walls and generally less elongate stomatal complexes.
 |
 |
 |
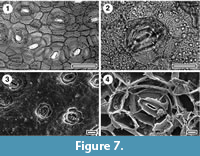
Halocarpus bidwilli
Figure 7
Specimens. SL5974, Baylys; SL5878, Ihamatao; SL2360, Mokau; SL5965, Omokoroa; LX2293, Ohawe-6; SL4636, Rangitawa-4; SL4388, Te Kowiwi.
Identification. New Zealand currently has three genera of podocarps that may display small, spirally oriented, scale-like leaves. These include Halocarpus, Manoao, and Lepidothamnus. The stomatal construction of all specimens in this study is consistent with Halocarpus, almost certainly H. bidwill i, These features include a prominent ring of subsidiary cells, which usually have distinctly thicker periclinal walls than normal epidermal cells, and this thickening is consistent from the outer margin to almost the edge of the guard cells, which are surrounded by a ring of thin cuticle (see figures in Florin, 1931; Wells and Hill, 1989; Jordan et al., 2011).
Phyllocladus sp.
Figure 8
Specimens. SL5968, Omokoroa; SL4980, Rangitawa-6.
Identification. Cuticle sheets with multiple rows of well-spaced, paratetracyctic stomatal complexes with lappets and well-developed Florin ring (Cookson and Pike, 1954; Hill, 1989). There are three extant species of Phyllocladus in New Zealand: P. alpinus, P. toatoa, and P. trichomanoides, but based on cuticle alone, these species are probably not distinct.
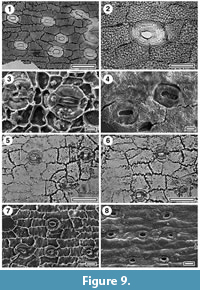
Prumnopitys taxifolia
Figure 9.1-4
Specimens. SL4981, Rangitawa-6; SL4675, Rapanui.
Identification. The specimens have the typical Prumnopitys stomatal distribution whereby stomates are widely-spaced along closely-spaced rows, making the nearest neighbour of a stomate usually in an adjacent row. They also have the typical stomates of this genus which have an “inflated” appearance. Of the two extant New Zealand species, they match P. taxifolia (compare figures in Greenwood et al., 2013)
Conifer gen indet.
Figure 9.5-8
Specimen. SL2355, Mokau.
Identification. A single specimen has epidermal cells with sinuous anticlinal cell walls. stomatal complexes longitudinally-aligned, widely-spaced (no clear row system), with a basically paratetracytic construction but inconsistent. There are similarities to Prumnopitys, but also to what was described as ‘Cycad gen. et sp. indet’ in Pole (2007c).
 |
 |
 |
Angiosperms
Monocots
CUT-Mo-FGD
Phormium tenax
Figure 10
Specimens. SL205, Dargaville; LX2731, Maxwell-13; SL2353, Mokau; LX2253, Ohawe-4; SL4649, Rangitawa-6; SL2325, Te Kowiwi.
Identification. The epidermis has alternating stomatal and non-stomatal zones. The stomatal complexes are aligned and tend to be arranged in an alternating fashion, the complexes are tightly-packed, and sometimes networked, so that some polar subsidiary cells are the lateral subsidiary cell of an adjacent complex. Subsidiary cells have several papillae. Some specimens are markedly less papillate, much less so than the Phormium tenax figured in Pole (2007d). It is not clear whether this is natural variation within the species, or is a different taxon (see also Conran et al., 2015, figure 6).
Astelia
Figure 11.1-4
Specimens. SL5978, Dargavillle; SL5913 Hauroto; SL5436, Hamiltons Gap.
Identification. Astelia has widely-spaced stomata, hidden under pairs of adjacent trichome bases (see Pole, 2007d; Maciunas et al., 2011; Conran et al., 2015).
Cyperaceae
Gen et sp. Indet.
Figure 11.5-6
Specimen. SL2376, Maxwell-07; LX2838, Maxwell-17; LX2274, Ohawe-5.
Identification. A single fragment of monocot cuticle from Maxwell shows stomatal complexes with pairs of distinct elongate and rounded subsidiary cells, and sinuous epidermal cells. This is regarded as Cyperaceae, genus unknown (cf. Conran et al., 2015, figure 8).
Arecaceae
Gen et sp. Indet.
Figure 11.7-8
Specimens. SL4671, Rapanui.
Identification. A single specimen shows broad palm morphology. However, it appears distinct from Rhopalostylus sapida, the single extant New Zealand palm (see Pole, 2007d).
Dicots
Winteraceae
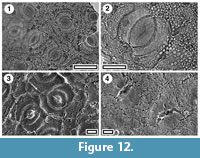
Pseudowintera colorata
Figure 12
Specimens. SL5457, Huriwai; SL2382, Maxwell-07; SL2361, Mokau; SL5452, Rangitawa-6.
Identification. The paracytic stomates and very granular cuticle clearly place this cuticle in the Winteraceae where it can be compared directly with extant New Zealand Pseudowintera colorata (see Pole 2007a, figure 11).
 |
 |
 |
Lauraceae
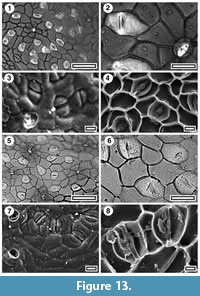
Beilschmiedia sp.
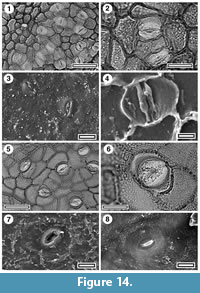
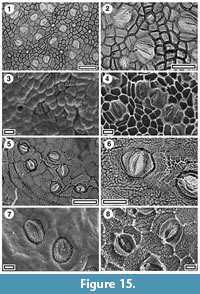
Figure 13, Figure 14, Figure 15.1-4
Specimens. SL0207, Dargaville; SL5959, Hauroto; SL4977, Huriwai; SL5883, Ihumatao; SL0211, Scottys Camp; SL4638, Rangitawa-4; SL2590, Waiuku; SL4495, Pahurehure.
Identification. Lauraceae cuticle is relatively easy to identify as such based on the presence of paracytic stomates overarched by subsidiary cells, and separated from them by cuticular “scales”, and, unless glabrous, by simple deciduous trichomes (Hill, 1986). Most of the Lauraceae cuticle found in this survey can mostly be compared directly with extant New Zealand Beilschmiedia. Depending on the authority followed, there are two or three Beilschmiedia species (Figure 13) in New Zealand (Wright, 1984) and also Litsea calicaris (Figure 14). Beilschmiedia tawa and B. tawaroa are probably not distinguishable on cuticular details. Both have somewhat angular stomatal complexes with truncated poles, thinner cuticle than normal epidermal cells, and narrow cuticular scales. On the external surface the outlines of the epidermal cells and subsidiary cells are clearly marked, and the stomatal pore is broad. B. tararoides is similar, except the external surface is much more subdued. The outlines of the epidermal cells are not obvious, and only the area of the subsidiary cells around the narrow stomatal pore is raised. In all Beilschmiedia the epidermal cell flanges are very clear in TLM. Thus, B. tawa/tawaroa and B. tararoides might be distinguishable using an SEM, though the difference is subtle.
Most of the fossil Lauraceae cuticle can be referred to Beilschmiedia, and most of that to B. tawa/tawaroa, particularly on the clearly defined cells on the outer surface. Material from Rangitawa is illustrated.
Litsea sp.
Figure 15.5-8
Specimens. SL2344, Mokau; SL4982, Rangitawa-6.
Identification. Litsea calicaris is distinct from New Zealand Beilschmiedia, having a much smoother and more defined stomatal outline, raised ledges around the outer stomatal pore, very subdued cuticular scales and a granularity to the inner and outer surface. The epidermal cell flanges can appear almost diffuse in TLM. Rangitawa has material comparable with Litsea calicaris, although at this stage it is prudent to refer to it as Litsea sp.
 |
 |
 |
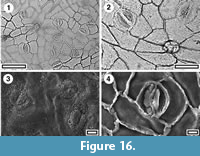
Cryptocarya sp.
Figure 16
Specimens. SL4977, Huriwai.
Identification. At Huriwai there is lauraceous cuticle that differs from extant New Zealand Beilschmiedia and Litsea in having well-developed, “butterfly-like”, cuticular scales, and a granular cover over the subsidiary cells and stomatal pore. The most likely identity of this is Cryptocarya (Christophel and Rowett, 1996), a genus now extinct in New Zealand.
Proteaceae
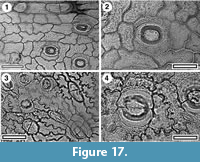
Toronia toru
Figure 17.1-2
Specimens. SL5880, Ihamatao; SL4498, Pahurehure.
Identification. Relatively large and aligned stomates and trichome bases associated with one or two modified epidermal cells identify it as Toronia toru (see figures in Pole, 1998; Carpenter et al., 2010).
Knightia excelsa
Figure 17.3-4
Specimens. SL4514, Pahurehure.
Identification. Randomly-oriented stomates and trichome attachment sites with over two to four modified epidermal cells and very sinuous epidermal cell flanges can be directly related to the extant Knightia excelsa (see figures in Pole, 1998).
Fabaceae
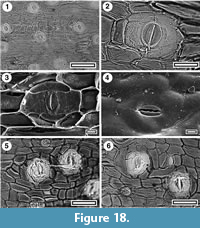
Carmichaelia sp.
Figure 18
Specimens. SL0204, Dargaville; SL2379, Maxwell-07; SL2351, Mokau; SL2365, Mokau; SL4515, Pahurehure; LX2251, Ohawe-4; LX2304, Rangitawa-4; SL4642, Rangitawa-5; SL5946, Rangitawa-6; SL0228, Scottys Camp.
Identification. This cuticle morphology is common and widespread. It is distinctive in having transversely-oriented stomates, organised in loose rows or narrow zones, and in elongate, flattened structures. Transversely-oriented stomates are such a restricted character that an unusual organ - i.e., a cladode, was immediately suggested. There was an initial suspicion that this may be the macro fossil corresponding to the Acaciapollenites pollen, which has been reported as from several Plio-Pleistocene localities in the North Island, including those covered here (Mildenhall, 1975a, b). However, Acacia was subsequently ruled out, and an extant New Zealand taxon, the cladodes of Carmichaelia, was recognised as the match. The species is not yet clear. Heenan (1998) recognised 23 species of Carmichaelia indigenous to New Zealand (and noted that they have transverse stomates). The cuticle fragments suggest one a species with larger cladodes, such as C. australis. This species would be consistent with the peaty facies (many other extant Carmichaelia are species of drylands), but the fossils have trichome-bases, which have not been observed on C. australis.
 |
 |
 |
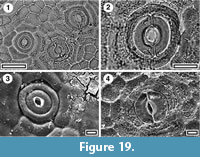
Grisseliniaceae
Grisellinia lucida
Figure 19
Specimen. SL5478, Hamiltons Gap; SL2346, Mokau.
Identification. The relatively large, round stomatal complexes, with a prominent Outer Stomatal Ledge and relatively isodiametric epidermal cell outlines identify this as Grisellinia lucida (see Pole 2008a, figure 35).
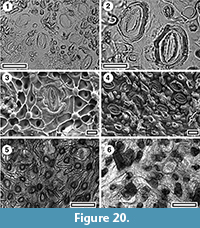
Asteraceae
Olearia sp.
Figure 20
Specimen. SL5480, Hamiltons Gap.
Identification. The key characters are an ovoid stomatal complex surrounded by a sharply-defined, narrow rim, and a mixture of abundant smooth, discrete papillae on the epidermal cells and trichome bases. This combination of characters is found in some of the extant New Zealand Olearia, for example O. arborescens.
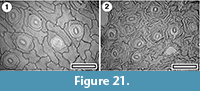
Myrsinaceae
Myrsine sp.
Figure 21
Specimen. SL4655, Rangitawa-6.
Identification. Cuticle with anisocytic stomatal complexes with either a pervasive ornamentation of ridges or restricted to the subsidiary cells is probably Myrsine. The peltate trichomes that would confirm the identification (see Pole, 2008a) have not been noted.
 |
 |
 |
CONCLUSIONS
Leaf cuticle has been found to be a common component in the Plio-Pleistocene ‘lignites’ in the North Island. A variety of taxa is represented, and is almost entirely of extant New Zealand species of forest and wetland angiosperms and conifers. However, some taxa are outside of their present-day range - the araucarian specimen at Rangitawa is notably far south of the range of Agathis today, while the presence of Halocarpus at sea level in the North Island is probably lower than it would occur today at that latitude. The existence of Cryptocarya, a possible Araucaria, and an indeterminate conifer from Mokau, support palynological evidence, of relatively recent plant extinctions in New Zealand.
There is clear scope for further work on this kind of material. Some sites still need to be dated, for example, Huriwai, and refining some identifications to species level would be useful. The New Zealand record has potential for providing stomatal indice variation of extant taxa, and thus further refining atmospheric carbon dioxide levels over several glacial-interglacial cycles. The macrofossils belonging to the Acacia pollen that stimulated this work still await discovery.
ACKNOWLEDGEMENTS
I thank J. McKee, J. Ogden, P. Moore, the Department of Conservation, Wanganui, for their help and assistance at various times. The Otago University Herbarium (OTA), the Allan Herbarium (CHR), Queensland Herbarium (AQ), Dunedin Botanical Gardens, and J. Ladley (University of Canterbury) are all thanked for contributing reference material for this project. The comments of two anonymous reviewers improved the manuscript and are appreciated.
REFERENCES
Atkinson, I.A.E. 1981. Vegetation map of Tongariro National Park, North Island, New Zealand. Scale 1:50,000. Wellington, DSIR
Beu, A.G. 2001. Local stages to be used for the Wanganui Series (Pliocene-Pleistocene), and their means of definition. New Zealand Journal of Geology and Geophysics, 44:113-125. https://doi.org/10.1080/00288306.2001.9514928
Bussell, M.R. 1986. Palynological evidence for upper Putikian (middle Pleistocene) interglacial and glacial climates at Rangitawa Stream, South Wanganui Basin, New Zealand. New Zealand Journal of Geology and Geophysics, 29:471-479. https://doi.org/10.1080/00288306.1986.10422168
Bussell, M.R. 1990. Palynology of oxygen isotope stage 6 and substage 5e from the cover beds of a marine terrace, Taranaki. New Zealand. Quaternary Research, 34:86-100. https://doi.org/10.1016/0033-5894(90)90074-u
Bussell, M.R. 1992. Late Pleistocene palynology of terrestrial cover beds at the type section of the Rapanui Terrace, Wanganui, New Zealand. Journal of the Royal Society of New Zealand, 22 :77-90. https://doi.org/10.1080/03036758.1992.10420807
Bussell, M.R. and Mildenhall, D.C. 1990. Extinct palynomorphs from middle and late Pleistocene terrestrial sediments, South Wanganui Basin, New Zealand. New Zealand Journal of Geology and Geophysics, 33:439-447. https://doi.org/10.1080/00288306.1990.10425699
Bussell, M.R. and Pillans, B. 1992. Vegetational and climatic history during oxygen isotope stage 9, Wanganui district, New Zealand, and correlation of the Fordell ash. Journal of the Royal Society of New Zealand, 22:41-60. https://doi.org/10.1080/03036758.1992.10420816
Byrami, M.L., Newnham, R.M., Alloway, B.V., Pillans, B., Ogden, J., Westgate, J., and Mildenhall, D.C. 2005. A late Early Pleistocene tephrochronological and pollen record from Auckland, New Zealand. Geological Society, London, Special Publications, 247 :183-208. https://doi.org/10.1144/gsl.sp.2005.247.01.10
Carpenter, R.J., Bannister, J.M., Jordan, G.J., and Lee, D.E. 2010. Leaf fossils of Proteaceae tribe Persoonieae from the Late Oligocene-Early Miocene of New Zealand. Australian Systematic Botany, 23:1-15. https://doi.org/10.1071/sb09015
Carter, R.M. and Naish, T.R. 1998a. A review of Wanganui Basin, New Zealand: global reference section for shallow marine, Plio-Pleistocene (2.5-0 Ma) cyclostratigraphy. Sedimentary Geology, 122:37-52. https://doi.org/10.1016/s0037-0738(98)00097-9
Carter, R.M. and Naish, T.R. 1998b. Have local stages outlived their usefulness for the New Zealand Pliocene-Pleistocene? New Zealand Journal of Geology and Geophysics, 41:271-279. https://doi.org/10.1080/00288306.1998.9514810
Chappell, J.M.A. 1970. Quaternary geology of the south-west Auckland coastal region. Transactions of the Royal Society of New Zealand, Earth Science, 8:133-153.
Christophel, D.C. and Rowett, A.I. 1996. Leaf and cuticle atlas of Australian leafy Lauraceae. Flora of Australia Supplementary Series Number 6. Australian Biological Resources Study, Canberra.
Conran, J.G., Bannister, J.M., Lee, D.E., Carpenter, R.J., Kennedy, E.M., Reichgelt, T., and Fordyce, R.E. 2015. An update of monocot macrofossil data from New Zealand and Australia. Botanical Journal of the Linnean Society, 178:394-420. https://doi.org/10.1111/boj.12284
Cookson, I.C. and Pike, K.M. 1954. The fossil occurrence of Phyllocladus and two other podocarpaceous types in Australia. Australian Journal of Botany, 2:60-67. https://doi.org/10.1071/bt9540060
Couper, R.A. and McQueen, D.R. 1954. Pliocene and Pleistocene plant fossils of New Zealand and their climatic interpretation. New Zealand Journal of Science and Technology, 35B:398-420.
Dawson, J.W. and Lucas, R. 2000. Nature Guide to the New Zealand Forest. Godwit, Auckland.
Ecroyd, C.E. 1982. Biological flora of New Zealand 8. Agathis australis (D. Don) Lindl. (Araucariaceae Kauri). New Zealand Journal of Botany, 20:17-36. https://doi.org/10.1080/0028825x.1982.10426402
Ferguson, D.K., 1985. The origin of leaf-assemblages--new light on an old problem. Review of Palaeobotany and Palynology, 46:117-188. https://doi.org/10.1016/0034-6667(85)90041-7
Fergusson, G.J. and Rafter, T.A. 1959. New Zealand C14 age measurements-4. New Zealand Journal of Geology and Geophysics, 2:208-241. https://doi.org/10.1080/00288306.1959.10431324
Fleming, C.A. 1953. The Geology of Wanganui Subdivision. New Zealand Geological Survey Bulletin n.s., 52:1-361.
Florin, R. 1931. Untersuchungen zur stammesgeschichte der Coniferales und Cordaitales. Kungliga Svenska Vetenskapakademiens handlinigar, 10:1-588.
Florin, R. and Boutelje, J.B. 1954. External morphology and epidermal structure of leaves in the genus Libocedrus, s.lat. Acta Horti Bergiani, 17:7-37.
Gastaldo, R.A. Staub, J.R. 1999. A mechanism to explain the preservation of leaf litter lenses in coals derived from raised mires. Palaeogeography, Palaeoclimatology, Palaeoecology, 149:1-14. https://doi.org/10.1016/s0031-0182(98)00188-6
Grant-Taylor, T.L. and Rafter, T.A. 1963. New Zealand natural radiocarbon measurements 1-5. Radiocarbon, 5:118-162. https://doi.org/10.1017/s0033822200036845
Greenwood, D.R. 1991. The taphonomy of plant macrofossils, p. 141-169. In Donovan, S.K. (ed.), The Processes of Fossilization. Columbia University Press, New York.
Greenwood, D.R., Hill, C.R., and Conran, J.G., 2013. Prumnopitys anglica sp. nov. (Podocarpaceae) from the Eocene of England. Taxon, 62:565-580. https://doi.org/10.12705/623.15
Grove, E. and Brandon, A. 2006. Red Bridge walk, Tairua. Waikato Botanical Society Newsletter, 23:4-5.
Hayward, B.W. 1977. North Island fossil forests. New Zealand Geological Society Newsletter, 114:28-32.
Hayward, J.J. and Hayward, B.W. 1995. Fossil forests preserved in volcanic ash and lava at Ihumatao and Takapuna, Auckland. Tane, 35:127-142.
Heads, M. 2017. Biogeography and Evolution in New Zealand. CRC Press, Taylor and Francis, Boca Raton.
Heenan, P.B. 1998. An emended circumscription of Carmichaelia, with new combinations, a key, and notes on hybrids. New Zealand Journal of Botany, 36:53-63. https://doi.org/10.1080/0028825X.1998.9512546
Henderson, J. and Ongley, M. 1923. The Geology of the Mokau Subdivision. New Zealand Geological Survey Bulletin n.s., 24:1-83.
Hill, R.S. 1986. Lauraceous leaves from the Eocene of Nerriga, New South Wales. Alcheringa, 10:327-351. https://doi.org/10.1080/03115518608619144
Hill, R.S. 1989. New species of Phyllocladus (Podocarpaceae) macrofossils from south eastern Australia. Alcheringa, 13:193-208. https://doi.org/10.1080/03115518908527820
Hönisch, B., Hemming, N.G., Archer, D., Siddall, M., and McManus, J.F. 2009. Atmospheric carbon dioxide concentration across the Mid-Pleistocene Transition. Science, 324:1551-1554. https://doi.org/10.1126/science.1171477
Jordan, G.J., Carpenter, R.J., Bannister, J.M., Lee, D.E., Mildenhall, D.C., and Hill, R.S. 2011. High conifer diversity in Oligo-Miocene New Zealand. Australian Systematic Botany, 24:121-136. https://doi.org/10.1071/sb11004
Kamp, P.J.J., Vonk, A.J., Bland, K.J., Hansen, R.J., Hendy, A.J.W., McIntyre, A.P., Ngatai, M., Cartwright, S.J., Hayton, S., and Nelson, C.S. 2004. Neogene stratigraphic architecture and tectonic evolution of Wanganui, King Country, and eastern Taranaki Basins, New Zealand. New Zealand Journal of Geology and Geophysics, 47:625-644. https://doi.org/10.1080/00288306.2004.9515080
Kershaw, P., Quilty, P.G., David, B., van Huet, S., and McMinn, A. 2000. Palaeobiogeography of the Quaternary of Australasia. Memoir of the Association of Australasian Palaeonotologists, 23:471-516.
Kohn, B.P., Pillans, B., and McGlone, M.S. 1992. Zircon fission track age for middle Pleistocene Rangitawa Tephra, New Zealand: stratigraphic and paleoclimatic significance. Palaeogeography, Palaeoclimatology, Palaeoecology, 95:73-94. https://doi.org/10.1016/0031-0182(92)90166-3
Maciunas, E., Conran, J.G., Bannister, J.M., Paull, R., and Lee, D.E. 2011. Miocene Astelia (Asparagales: Asteliaceae) macrofossils from southern New Zealand. Australian Systematic Botany, 24:19-31. https://doi.org/10.1071/sb10035
Marra, M.J., Alloway, B.V., and Newnham, R.M. 2006. Paleoenvironmental reconstruction of a well-preserved Stage 7 forest sequence catastrophically buried by basaltic eruptive deposits, northern New Zealand. Quaternary Science Reviews, 25:2143-2161. https://doi.org/10.1016/j.quascirev.2006.01.031
McDougall, I., Polach, H.A., and Stipp, J.J. 1969. Excess radiogenic argon in young subaerial basalts from the Auckland volcanic field, New Zealand. Geochimica et Cosmochimica Acta, 33:1485-1520. https://doi.org/10.1016/0016-7037(69)90152-5
McGlone, M.S. 1983. History of New Zealand lowland forest since the last glaciation, p. 1-17. In Thompson, K., Hodder, A.P.W., Edmonds, A.S. (eds.), Lowland forests in New Zealand. University of Waikato, University of Waikato, Hamilton, New Zealand.
McGlone, M.S. 1988. New Zealand, p. 557-602. In Huntley, B. and Webb, III, T. (eds.), Vegetation History. Dordrecht, Kluwers.
McGlone, M.S., Neall, V.E., and Pillans, B.J. 1984. Inaha Terrace deposits: A late Quaternary terrestrial record in South Taranaki, New Zealand. New Zealand Journal of Geology and Geophysics, 27:35-49. https://doi.org/10.1080/00288306.1984.10422290
McGlone, M.S., Wardle, P., and Worthy, T.H. 2003. Environmental change since the last glaciation, p. 108-127. In Darby, J., Fordyce, R.E., Mark, A., Probert, K., and Townsend, C. (eds.), The Natural History of Southern New Zealand. Otago University Press in association with the Otago Museum, Dunedin.
McGregor, W.R. 1948. The Waipoua Kauri Forest. Abel, Dykes, Auckland.
McQueen, D.R. 1954. Fossil leaves, fruits and seeds from the Wanganui Series (Plio-Pleistocene) of New Zealand. Transactions of the Royal Society of New Zealand, 82:667-676.
Mildenhall, D.C. 1975a. Palynology of the Acacia -bearing beds in the Komako District, Pohangina Valley, North Island, New Zealand. New Zealand Journal of Geology and Geophysics, 18:209-228. https://doi.org/10.1080/00288306.1975.10418194
Mildenhall, D.C. 1975b. Lower Pleistocene palynomorphs from the Ohuka Carbonaceous Sandstone, south-west Auckland, New Zealand. New Zealand Journal of Geology and Geophysics, 18:675-681. https://doi.org/10.1080/00288306.1975.10421568
Mildenhall, D.C. 1985. Quaternary palynology: North Kaipara Barrier. Appendix in: Richardson, R.J.H. Quaternary geology of the north Kaipara Barrier, Northland, New Zealand. New Zealand Journal of Geology and Geophysics, 28:121-127. https://doi.org/10.1080/00288306.1985.10422280
Mildenhall, D.C. 1986. Middle Quaternary pollen diagrams from Judgeford, Lower Hutt, and the significance of kauri pollen in a herbaceous assemblage. New Zealand Geological Survey Record, 8:76-81.
Mildenhall, D.C., Stokes, S., Nelson, C.S. 1992. Palynology, age, and paleoenvironments of carbonaceous facies in the Kaihu Group (Late Pliocene-Pleistocene), northern North Island. New Zealand Geological Survey Record, 46:1-44.
Moar, N.T. and McKellar, I.C. 2001. Interglacial vegetation in South Westland, South Island, New Zealand. New Zealand Journal of Geology and Geophysics, 44:17-24. https://doi.org/10.1080/00288306.2001.9514918
Moar, N.T. and Suggate, R.P. 1996. Vegetation history from the Kaihinu (last) Interglacial to the present, West Coast, South Island, New Zealand. Quaternary Science Reviews, 15:521-547. https://doi.org/10.1016/0277-3791(96)00004-2
Naish, T. and Kamp, P.J.J. 1995. Pliocene-Pleistocene marine cyclothems, Wanganui Basin, New Zealand: a lithostratigraphic framework. New Zealand Journal of Geology and Geophysics, 38:223-243. https://doi.org/10.1080/00288306.1995.9514651
Newnham, R. and Lusk, C. 1987. Comparison of plant micro- and macrofossils, Kariotahi, Awhitu Peninsula. Tane, 32:171-178.
Newnham, R.M., Lowe, D.J., and Williams, P.W. 1999. Quaternary environmental change in New Zealand; a review. Progress in Physical Geography, 23:567-610. https://doi.org/10.1177/030913339902300406
Ogden, J., Wilson, A., Hendy, C., and Newnham, R. 1992. The Late Quaternary history of kauri (Agathis australis ) in New Zealand and its climatic significance. Journal of Biogeography, 19:611-622. https://doi.org/10.2307/2845704
Ogden, J., Newnham, R.M., Palmer, J.G., Serra, R.G., and Mitchell, N.D. 1993. Climatic implications of macro-and microfossil assemblages from Late Pleistocene deposits in northern New Zealand. Quaternary Research, 39:107-119. https://doi.org/10.1006/qres.1993.1013
Pilllans, B.J. 1983. Upper Quaternary marine terrace chronology and deformation, South Taranaki, New Zealand. Geology, 11:292-297. https://doi.org/10.1130/0091-7613(1983)11<292:uqmtca>2.0.co;2
Pillans, B.J. 1988. Loess chronology in Wanganui Basin. New Zealand, p. 175-191. In Eden, D.N. and Furkert, R.J. (eds.), Loess, its Distribution, Geology and Soils. Rotterdam, A.A. Balkema.
Pillans, B.J. 1990. Late Quaternary marine terraces, South Taranaki - Wanganui. NZ Geological Survey Micellaneous Series Map 18.
Pillans, B. 1994. Direct-marine-terrestrial correlations, Wanganui Basin, New Zealand: the last 1 million years. Quaternary Science Reviews, 13:189-200. https://doi.org/10.1016/0277-3791(94)90024-8
Pillans, B. 2017. Quaternary stratigraphy of Whanganui Basin--A globally significant archive, p. 141-170. In Shulmeister, J. (ed.), Landscape and Quaternary Environmental Change in New Zealand, Advances in Quaternary Science, 3. Atlantis Press, Amsterdam. https://doi.org/10.2991/978-94-6239-237-3_4
Pocknall, D.T. and Mildenhall, D.C. 1984. Late Oligocene -Early Miocene spores and pollen from Southland, New Zealand. New Zealand Geological Survey Paleontological Bulletin, 51:1-66.
Pocknall, D.T. and Waterhouse, B.C. 1984. Age, paleoenvironment and correlation of carbonaceous horizons, Aotea Harbour, South Auckland, New Zealand. New Zealand Geological Survey Record, 3:8-16.
Polach, H.A., Chappell, J., and Lovering, J.F. 1969. Australian National University radiocarbon date list. Radiocarbon, 11:245-262. https://doi.org/10.1017/s0033822200011206
Pole, M.S. 1998. The Proteaceae record in New Zealand. Australian Systematic Botany, 11:343-372. https://doi.org/10.1071/sb97019
Pole. M. 2007a. Plant-macrofossil assemblages during Pliocene uplift, South Island, New Zealand. Australian Journal of Botany, 55:118-142. https://doi.org/10.1071/bt06055
Pole, M. 2007b. Lauraceae macrofossils and dispersed cuticle from the Miocene of southern New Zealand. Palaeontologia Electronica Vol. 10.1.3A: 1-38
https://palaeo-electronica.org/2007_1/zealand/index.html
Pole, M. 2007c. Conifer and cycad distribution in the Miocene of southern New Zealand. Australian Journal of Botany, 55:143-164. https://doi.org/10.1071/bt06056
Pole, M. 2007d. Monocot macrofossils from the Miocene of southern New Zealand. Palaeontologia Electronica Vol. 10.3.15A: 1-21
https://palaeo-electronica.org/2007_3/125/index.html
Pole, M.S. 2008a. Dispersed Leaf Cuticle from the Early Miocene of Southern New Zealand. Palaeontologia Electronica Vol. 11.3.15A: 1-117
https://palaeo-electronica.org/2008_3/153/index.html
Pole, M. 2008b. The record of Araucariaceae macrofossils in New Zealand. Alcheringa, 32:409-430. https://doi.org/10.1080/03115510802417935
Pole, M., Dawson, J., and Denton, T. 2008. Fossil Myrtaceae from the Early Miocene of southern New Zealand. Australian Journal of Botany, 56:67-81. https://doi.org/10.1071/bt07032
Raine, J.I., Beu, A.G., Boyes, A.F., Campbell, H.J., Cooper, R.A., Crampton, J.S., Crundwell, M.P., Hollis, C.J., and Morgans, H.E.G. 2015. Revised calibration of the New Zealand Geological Timescale: NZGT2015/1. GNS Science Report 2012/39, 1-53.
Ricketts, B.D. 1977. The Coastal Geology of the Ihumatao area, Auckland. Tane, 23:119-124.
Richardson, R.J.H. 1985. Quaternary geology of the north Kaipara Barrier, Northland, New Zealand. New Zealand Journal of Geology and Geophysics, 28:111-127. https://doi.org/10.1080/00288306.1985.10422280
Spicer, R.A. 1991. Plant taphonomic processes, p. 71-113. In Allison, P.A. and Briggs, D.E.G. (eds.), Taphonomy: Releasing the Data Locked in the Fossil Record. Plenum Press, New York.
Te Punga, M.T. 1952. The geology of Rangitikei Valley. New Zealand Geological Survey Memoir, 8:1-46.
Te Punga, M.T. 1962. The Rangitawa fossil beds of northwest Wellington. New Zealand Journal of Geology and Geophysics, 5:493-498. https://doi.org/10.1080/00288306.1962.10420102
Wardle, P. 1991. Vegetation of New Zealand. Cambridge University Press, Cambridge.
Wards, A.J. 1996. An Investigation of the Rapanui and Ngarino Marine Terraces in South Taranaki: Coverbed Stratigraphy, Distribution and Tectonics. Unpublished MSc thesis, Massey University, Palmerston North, New Zealand.
Wells, P.M. and Hill, R.S. 1989. Leaf morphology of the imbricate-leaved Podocarpaceae. Australian Systematic Botany, 2:369-386. https://doi.org/10.1071/sb9890369
Wood, I. 1991. Thermoluminescence dating of the Auckland and Kerikeri Basalt fields. Unpublished M.Sc. thesis, University of Auckland, New Zealand.
Wright, A.E. 1984 Beilschmiedia Nees (Lauraceae) in New Zealand. New Zealand Journal of Botany, 22:109-125. https://doi.org/10.1080/0028825x.1984.10425238
