

 Comparison of methods: Micro-CT visualization method and epoxy cast-embedding reveal hidden details of bioerosion in the tube walls of Cretaceous polychaete worms
Comparison of methods: Micro-CT visualization method and epoxy cast-embedding reveal hidden details of bioerosion in the tube walls of Cretaceous polychaete worms
Article number: 26.2.a18
https://doi.org/10.26879/1255
Copyright Paleontological Society, May 2023
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 23 November 2022. Acceptance: 5 May 2023
ABSTRACT
Bioerosion in three serpulid tubes of the (sub-)genera Cementula, Pyrgopolon (Septenaria), and Placostegus from the Bohemian Cretaceous Basin was studied by a combination of micro-computed tomography and vacuum cast-embedding technique producing polymer resin casts. Results gained from both methods were evaluated and compared in terms of material usability, destructive force, quality of the resulting image, and hardware/software requirements. The advantage of the micro-CT methodology is its non-destructiveness and the ability to make three-dimensional images, animations, and serial sections through the object; this method is suitable for most examined materials, but it is limited by low quality of the resulting image and demanding hardware/software requirements. To ensure good visibility of the borings in the resulting X-ray reconstructions, a material contrast between hard substrate and borings is necessary. The advantage of the vacuum cast-embedding technique followed by SEM observation is high quality of the resulting image, but it is limited by destructive force and material composition. Vacuum casting is unusable for undissolvable silicified or pyritized substrates, as happened in our Placostegus zbyslavus specimen, which suffered from incomplete dissolution in acid. However, epoxy casting provided detailed morphology of borings that were beyond the detection limit of the given spatial resolution of the micro-CT scanner, and helped with ichnotaxonomic identification. Tiny branching canals of Iramena in Cementula sp. and the finer apertural canals connecting the Entobia chambers to the substrate surface in Pyrgopolon (Septenaria) cf. tricostata and Cementula sp. were not detected using micro-CT, but were visible in SEM images of epoxy resin casts.
Zuzana Heřmanová. Palaeontological Department, Natural History Museum, Cirkusová 1740, 193 00, Prague 9, Czech Republic. zuzana.hermanova@nm.cz
Martina Kočová Veselská. Institute of Geology of the Czech Academy of Sciences, v. v. i., Department of Palaeobiology and Paleoecology, Rozvojová 269, 165 00 Prague 6, Lysolaje, Czech Republic and Institute of Geology and Palaeontology, Charles University, Albertov 6, 128 43 Prague 2, Czech Republic. martina.veselska@natur.cuni.cz
Tomáš Kočí. Palaeontological Department, Natural History Museum, Cirkusová 1740, 193 00, Prague 9, Czech Republic and Ivančická 581, 19900 Prague 9 - Letňany, the Czech Republic. protula@seznam.cz
Manfred Jäger. Lindenstrasse 53, 72348 Rosenfeld, Germany. langstein.jaeger@web.de
Jana Bruthansová. Palaeontological Department, Natural History Museum, Cirkusová 1740, 193 00, Prague 9, Czech Republic. jana.bruthansova@nm.cz
Radek Mikuláš. Institute of Geology of the Czech Academy of Sciences, v. v. i., Department of Palaeobiology and Paleoecology, Rozvojová 269, 165 00 Prague 6, Lysolaje, Czech Republic. mikulas@gli.cas.cz
Keywords: micro-computed tomography; vacuum casting; Serpulidae; boreholes; late Cretaceous
Final citation: Heřmanová, Zuzana, Kočová Veselská, Martina, Kočí, Tomáš, Jäger, Manfred, Bruthansová, Jana, and Mikuláš, Radek. 2023. Comparison of methods: Micro-CT visualization method and epoxy cast-embedding reveal hidden details of bioerosion in the tube walls of Cretaceous polychaete worms. Palaeontologia Electronica, 26(2):a18.
https://doi.org/10.26879/1255
palaeo-electronica.org/content/2023/3585-comparison-of-methods-micro-ct-and-epoxy-cast-embedding
Copyright: May 2023 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
In recent decades, vacuum cast-embedding and following-up scanning electron microscopy visualization (SEM) became the most widely applied procedure for the study of recent and fossil bioerosive structures in carbonate substrates (e.g., Golubic et al., 1970; Wisshak, 2012; Seuss and Nützel, 2019). This method was introduced by Maaløe and Birch-Andersen (1956), followed by Glauert et al. (1956) for biomedical purposes. Since the 1970s, it has been applied in ichnology with several modifications (e.g., Golubic et al., 1970). Nevertheless, this method still required pressure impregnation and heating of embedding medium at about 60°C for at least 40 hours, and thus is not suitable for material sensitive to heat or pressure. Nielsen and Maiboe (2000), followed by Wisshak (2012), introduced lesstime-consuming vacuum cast-embedding, based on filling bioerosional tunnels with a mixture of epoxy resin and hardener (Epofix) under vacuum conditions and dissolving the substrate with diluted hydrochloric acid. This procedure is used in the present paper.
Micro-computed tomography has recently experienced a boom in all branches of biology, paleontology, and geology. Micro-CT devices utilize X-rays to see inside an object, enabling virtual three-dimensional reconstruction of the specimen, and generation of videos using individual section images. It is intensively used for study of recent (Mees et al., 2003; Beuck et al., 2006; Schönberg and Shield, 2008; Lukeneder and Lukeneder, 2011; Lukeneder, 2012; Lukeneder et al., 2014; Amon et al., 2015; Färber et al., 2016; Silbiger et al., 2016) and fossil bioerosional traces (e.g., Benner et al., 2008; Beuck et al., 2008; Lee et al., 2017; Wisshak et al., 2017; Heřmanová et al., 2020). In our study, we used a combination of micro-computed tomography and vacuum cast-embedding technique producing polymer resin casts followed by SEM observations to visualize bioerosion inside serpulid polychaete tubes. Serpulids are commonly represented in the fossil record by their calcitic or calcitic/aragonitic tubes (e.g., Jäger, 1983, 2005, 2012; Vinn et al., 2008a+b+c+d; ten Hove and Kupriyanova, 2009; Vinn and Mutvei, 2009; Ippolitov et al., 2014; Vinn 2020), but to date, only a few publications referred to domichnial traces inside serpulid tubes (Kočí et al., 2017; Heřmanová et al., 2020; Kočová Veselská et al., 2021; Mikuláš et al., 2022). The low bioerosion degree is strongly affected by the serpulid tube curvature and very tiny tube walls, ranging between 0.4-0.6 mm. However, in the case of rarity of any other suitable thick-shelled fauna at the locality, serpulids could be surprisingly highly infested, as shown in Kočová Veselská et al. (2021). The aim of this paper is:
(1) to document bioeroded serpulid polychaetes from the late Cretaceous of the Bohemian Cretaceous Basin,
(2) to evaluate results from micro-CT and vacuum cast-embedding - how the method used affects the display and subsequent description of boreholes in serpulid tubes, and
(3) to compare advantages and limitations of both methods.
GEOLOGICAL AND GEOGRAPHICAL SETTING
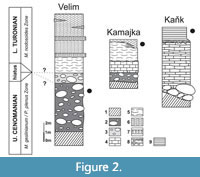
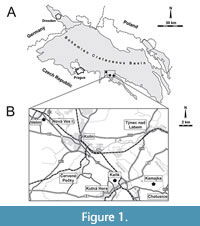 The bioeroded serpulid tubes were collected from nearshore, shallow-water deposits at Velim, Kamajka, and Kaňk “Na Vrších”, situated along the southern and eastern margin of the Bohemian Cretaceous Basin (Figure 1A), which are interpreted to have been laid down under high-energy conditions (Žítt et al., 1997). Localities are situated approximately 60-100 km east of Prague in the vicinity of Kolín and Kutná Hora (Figure 1B), where nearshore sediments are exposed in depressions of metamorphic rocks. The bioeroded serpulids from Kaňk and Kamajka treated in the present publication are early Turonian in age, whereas those from Velim are late Cenomanian (Figure 2). The exact correlation among the localities is complicated by frequent non-sequences and redeposition of older faunas (Kočová Veselská et al., 2014). The sedimentation was also influenced by palaeohydrodynamics, palaeogeography, clast distribution, and character and abundance of detrital material (Žítt and Nekvasilová, 1989, 1994, 1996; Žítt 1992; Žítt et al. 1998, 2006). Generally, the horizon yielding serpulid polychaetes is developed in characteristic facies, consisting of calcareous claystones at Kaňk “Na Vrších”, organodetritic clay limestones at Kamajka, and calcareous siltstones with abundant organodetritus at Velim and Kamajka (Figure 2). The rich fauna from Velim and Kamajka was studied and described by Žítt et al. (1997) and Košťák et al. (2010); fossil communities from Kaňk “Na Vrších” were examined by Vodrážka et al. (2013).
The bioeroded serpulid tubes were collected from nearshore, shallow-water deposits at Velim, Kamajka, and Kaňk “Na Vrších”, situated along the southern and eastern margin of the Bohemian Cretaceous Basin (Figure 1A), which are interpreted to have been laid down under high-energy conditions (Žítt et al., 1997). Localities are situated approximately 60-100 km east of Prague in the vicinity of Kolín and Kutná Hora (Figure 1B), where nearshore sediments are exposed in depressions of metamorphic rocks. The bioeroded serpulids from Kaňk and Kamajka treated in the present publication are early Turonian in age, whereas those from Velim are late Cenomanian (Figure 2). The exact correlation among the localities is complicated by frequent non-sequences and redeposition of older faunas (Kočová Veselská et al., 2014). The sedimentation was also influenced by palaeohydrodynamics, palaeogeography, clast distribution, and character and abundance of detrital material (Žítt and Nekvasilová, 1989, 1994, 1996; Žítt 1992; Žítt et al. 1998, 2006). Generally, the horizon yielding serpulid polychaetes is developed in characteristic facies, consisting of calcareous claystones at Kaňk “Na Vrších”, organodetritic clay limestones at Kamajka, and calcareous siltstones with abundant organodetritus at Velim and Kamajka (Figure 2). The rich fauna from Velim and Kamajka was studied and described by Žítt et al. (1997) and Košťák et al. (2010); fossil communities from Kaňk “Na Vrších” were examined by Vodrážka et al. (2013).
MATERIAL AND METHODS
 Thousands of serpulid tubes collected by Tomáš Kočí at Velim, Kamajka, and Kaňk were visually examined for evidence of boreholes using an Olympus DP70 light microscope at the National Museum in Prague. Of these, three serpulid tubes exemplifying the range of bioerosional structures were selected for closer examination: one tube of Cementula sp. from Velim (CZ2), one fragment of Pyrgopolon (Septenaria) cf. tricostata (Goldfuss, 1841) from Kaňk “Na Vrších” (NM O8728), and one tube of Placostegus zbyslavus (Ziegler, 1984) from Kamajka near Chotusice (NM O8727). All material is kept in the paleontological collection of the National Museum in Prague.
Thousands of serpulid tubes collected by Tomáš Kočí at Velim, Kamajka, and Kaňk were visually examined for evidence of boreholes using an Olympus DP70 light microscope at the National Museum in Prague. Of these, three serpulid tubes exemplifying the range of bioerosional structures were selected for closer examination: one tube of Cementula sp. from Velim (CZ2), one fragment of Pyrgopolon (Septenaria) cf. tricostata (Goldfuss, 1841) from Kaňk “Na Vrších” (NM O8728), and one tube of Placostegus zbyslavus (Ziegler, 1984) from Kamajka near Chotusice (NM O8727). All material is kept in the paleontological collection of the National Museum in Prague.
All selected specimens were cleaned in a 7% solution of hydrogen peroxide in an ultrasonic bath. For visualising and determining trace fossils left by bioeroders inside the serpulid tubes, X-ray micro-tomography, and a vacuum cast-embedding technique producing polymer resin casts were applied to all examined specimens.
The micro-computed tomography analysis was performed by using an X-ray micro-tomography SkyScan 1172 (Bruker) device at the Natural Museum in Prague (Czech Republic). The system provides several X-ray “shadow” transmission images from many views in different angles while the object rotates. From these shadow images, cross-section images of the object were reconstructed using N-Recon Version: 2.0.0 Software, creating a complete three-dimensional representation of internal microstructure. Images were created by Avizo (Version: 2020.3) software. Revealed information was examined using the Volume Rendering and Ortho-slice mode of surface and/or sections. Isosurface visualization and segmentation were applied to effectively display the structures of the borings. The X-ray microfocus tube was operated at 100-140 µA and 70-100 kV. Filters used were Al (0.5 mm) and combined filter Al+Cu or Cu filter (1mm). Rotation Step was 0.2-0.4°, two frames for one-step were used, random movement was between 2 and 10, and 180° rotation was used. Average image pixel size was 1.96-13.54 µm.
Subsequently, all serpulid tubes were cast in epoxy resin, using the technique of Wisshak (2012). Samples were placed in a small vacuum chamber for cast-embedding (Struers CitoVac) and infiltrated with a mixture of EpoFix low-viscosity epoxy resin and hardener in a ratio of 15/2 ml. After curing, the samples were longitudinally cut in half with a rock saw, decalcified by treatment with 10% HCl (except for silicified tube of NM O8727, whose solubility in acids was very low and did not dissolve even in 48% HF), rinsed in distilled water to prevent further etching and possible chemical precipitation, and dried. The polymer resin casts were then mounted on stubs before gold coating (NM O8727) and SEM analysis (Hitachi S-3700 N SEM at National Museum in Prague), to display the most delicate morphological features.
REMARKS ON BIOERODED SERPULID WORM TUBES
Many polychaete worms, many genera of Serpulidae and the sabellid Glomerula, differ from other polychaetes by forming a calcareous (calcitic or calcitic/aragonitic) two-layered tube (Vinn et al., 2008b). Since the Permian (Sanfilippo et al., 2018; Ramsdale, 2021), the vast majority of serpulids are typical encrusters in the marine realm, and today a single genus lives in the freshwater of karst caves. Their tubes are usually attached to hard substrates like rocks, pebbles, shells, and other solid skeletal remains of various marine animals, as well as, although less often, to algae and seagrasses. Few species become free-lying loosely on or sticking in soft ground (e.g., Ditrupa, some species of Pyrgopolon). Serpulids and sabellids are also characteristic elements of the mesofauna in the rocky coast facies situated at the southwestern and southern margin of the Bohemian Cretaceous Basin (BCB), where thousands of specimens of a dozen genera have been collected to date. They have been documented in numerous publications (Reuss, 1845, 1846; Geinitz, 1875a, 1875b; Ziegler, 1984; Jäger and Kočí, 2007; Kočí, 2008, 2010, 2012; Jäger, 2014; Kočí and Jäger, 2015).
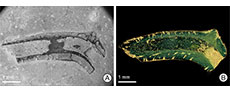
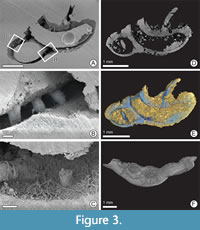 Despite such a large number of serpulids in the BCB, only a few specimens show signs of domichnial bioerosion, of which three tubes were selected for closer examination: one specimen each of Cementula sp. from Velim, Placostegus zbyslavus (Ziegler, 1984) from Kamajka near Chotusice and Pyrgopolon (Septenaria) cf. tricostata (Goldfuss, 1841) from Kaňk Na Vrších. Cementula sp. (Figure 3) has a coiled tube with its outer whorls usually attached to the inner whorls; boundaries between whorls are smoothed by deposition of calcareous tube material. Placostegus zbyslavus (Figure 4) is preserved as a 6 mm long fragment of the
Despite such a large number of serpulids in the BCB, only a few specimens show signs of domichnial bioerosion, of which three tubes were selected for closer examination: one specimen each of Cementula sp. from Velim, Placostegus zbyslavus (Ziegler, 1984) from Kamajka near Chotusice and Pyrgopolon (Septenaria) cf. tricostata (Goldfuss, 1841) from Kaňk Na Vrších. Cementula sp. (Figure 3) has a coiled tube with its outer whorls usually attached to the inner whorls; boundaries between whorls are smoothed by deposition of calcareous tube material. Placostegus zbyslavus (Figure 4) is preserved as a 6 mm long fragment of the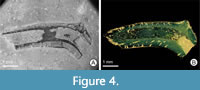 possibly curved or coiled attached posterior tube portion, followed by the long, nearly straight free anterior tube part, which rises almost vertically, is slightly twisted around its longitudinal axis, bears three keels, and is triangular in cross-section. Pyrgopolon (Septenaria) cf. tricostata (Figure 5) is represented by a 6 mm long horn-shaped tube with three prominent and wide longitudinal ridges subcircular in cross-section, laterally divided by furrows on the external surface.
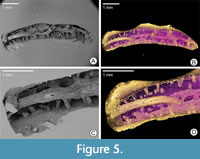
possibly curved or coiled attached posterior tube portion, followed by the long, nearly straight free anterior tube part, which rises almost vertically, is slightly twisted around its longitudinal axis, bears three keels, and is triangular in cross-section. Pyrgopolon (Septenaria) cf. tricostata (Figure 5) is represented by a 6 mm long horn-shaped tube with three prominent and wide longitudinal ridges subcircular in cross-section, laterally divided by furrows on the external surface.
 Obviously the very thin wall of serpulid tubes, ranging between 0.4-0.6 mm, is the reason for the low bioerosion degree in such tubes. The nearshore deposits in the BCB yielded numerous thicker-shelled fauna (bivalves, brachiopods, echinoids) as well as basal conglomerates representing a more suitable substrate for domichnia (e.g., Žítt et al., 2006, 2015; Vodrážka et al., 2013). Moreover, all studied serpulids come from high-energy environment, where it is difficult for organisms to settle on substrates that are frequently buried and exhumed; the degree of bioerosion thus depends on the duration of exposure before burial in the sediment (El-Hedeny, 2007; Madani et al., 2022). Because serpulids were moved and rolled on the seafloor, they all are only fragmentarily preserved and strongly mechanically damaged. Besides fragmentation, tube surfaces show evidence of selective dissolution and abrasion generated by abiotic processes. The most detailed paper on sclerobionts associated with polychaetes from BCB is that of Kočí and Jäger (2015), concerning borings and encrusters on sabellid and serpulid worms from the rocky coast facies (late Cenomanian) at Předboj near Prague.
Obviously the very thin wall of serpulid tubes, ranging between 0.4-0.6 mm, is the reason for the low bioerosion degree in such tubes. The nearshore deposits in the BCB yielded numerous thicker-shelled fauna (bivalves, brachiopods, echinoids) as well as basal conglomerates representing a more suitable substrate for domichnia (e.g., Žítt et al., 2006, 2015; Vodrážka et al., 2013). Moreover, all studied serpulids come from high-energy environment, where it is difficult for organisms to settle on substrates that are frequently buried and exhumed; the degree of bioerosion thus depends on the duration of exposure before burial in the sediment (El-Hedeny, 2007; Madani et al., 2022). Because serpulids were moved and rolled on the seafloor, they all are only fragmentarily preserved and strongly mechanically damaged. Besides fragmentation, tube surfaces show evidence of selective dissolution and abrasion generated by abiotic processes. The most detailed paper on sclerobionts associated with polychaetes from BCB is that of Kočí and Jäger (2015), concerning borings and encrusters on sabellid and serpulid worms from the rocky coast facies (late Cenomanian) at Předboj near Prague.
The surface of the investigated serpulids is highly pitted by circular to oval borehole apertures measuring 0.01-0.4 mm in diameter. Because the thickness of serpulid tube walls is not constant throughout the tube, but decreases towards the aperture and increases at the longitudinal ridges and keels, domichnia should be expected rather in the thicker parts of the tube wall. However, combining micro-CT and vacuum cast-embedding proved that boreholes are evenly distributed, regardless of the morphology of the host tubes, and revealed borings attributable to four ichnogenera: Entobia Bronn, 1837, Trypanites Mägdefrau, 1932, Maeandropolydora Voigt, 1965, and Iramena Boekschoten, 1970. Borings were analyzed separately in each serpulid tube to look for any specific associations or relationships, and their characteristics are described below:
Entobia has ovoid or bag-like chambers, connected to the surface by narrow tunnels. As the chambers occur very close to the surface of the serpulid tubes, because of the very thin tube walls, some connecting tunnels are very short (length ranges from 0.1 to 2 mm) and narrow (diameter varies from 0.01 to 0.4 mm). Because SEM images of the resulting casts have significantly better resolution than micro-CT, tiny chambers are clearly visible only on SEM images of epoxy casts. Specimen NM O8728 shows a trinity of two-camerate, small (up to 100 μm) spherical bodies, partially penetrating each other (Figure 5C). From the outside of the substrate, the spheres appear smooth at a particular resolution. Their surfaces correspond to the grain size (much smaller than the object being examined) of the mineralized filling (<10 μm). On the sediment-facing side, six “stems,” shorter than 10 μm, protrude from the two spherical bodies, corresponding to exploratory threads of entobians (Bromley and D’Alessandro, 1984). Another two Entobia specimens on sample NM O8728 show comparable morphology and dimensions, but are in the back of the epoxy cast and so not easily accessible for study (Figure 5A). A sole well-casted bag-like chamber was also found in sample CZ2 (Figure 3A).
Trypanites are represented by thin, short, smooth, and unbranched shafts, usually passing roughly vertically through the whole preserved wall of a serpulid tube. The diameter of individual shafts tends to be constant, ranging from 0.1 to 0.2 mm, oval in cross-section with rounded or gently oval openings. The usual length (depth) of the tubes is 0.25 to 0.4 mm, which corresponds to the thickness of the serpulid tube walls. Specimens NM O8727 and O8728 display a minimum of 30 short shafts of Trypanites borings, which includes about one-half of the original serpulid tube. Whereas Trypanites in O8728 are visible in micro-CT (Figure 5B, D) as well as in SEM images of polymer resin cast (Figure 5A, C), Trypanites in NM O8727 were readily discernible only by using micro-CT reconstruction (Figure 4B). The epoxy casting was unsuccessful because of an incompletely dissolved tube, and the resulting image shows only signs of a few short shafts (Figure 4A). Specimen CZ2 has at least three incompletely preserved borings visible in micro-CT (Figure 3E) and SEM pictures (Figure 3A, B).
Maeandropolydora found in tube of Pyrgopolon (Septenaria) cf. tricostata (NM O8728) consists of long, cylindrical galleries with several apertures, running through the substrate in irregular, not too sharp turns. Some of the views of the epoxy cast show that the galleries are duplicated, partially touching each other; in the remaining part, they are somewhat less densely spaced but remain roughly parallel (Figure 5). The length of the individual boring is 5.8 mm, which corresponds to the overall length of the studied serpulid. The diameter of the galleries is somewhat variable but basically circular; it ranges from 0.3 to 0.5 mm. The surface of the galleries is smooth. Serpulid tube O8728 was filled with calcium carbonate and was redeposited in a high physical energy environment of the rocky shore. Then this cylindrical object was available for available for boring. The resulting configuration of borings is therefore quite different from the other studied specimens, which have the character of relatively thin-walled tubes.
Cementula sp. (specimen CZ2) micro CT and the epoxy cast revealed three short cylindrical to somewhat conical fillings of borings through the entire wall. In the micro-CT image, these borings resemble short shafts oriented perpendicular to the tube surface (Figure 3E). The axes of the bored cylinders/cones form an angle of 70-85° with the surface of the host tube, which is close to the values found for Trypanites. However, the SEM image of the epoxy cast shows “branching” of these main borings (Figure 3C). The branching stolons, below the lower limit of micro-CT resolution, are characterized by a significantly smaller and variable diameter (10-30 µm), compared to the main boring. The axes of the stolons form an angle of 30-60° with the axis of the main borings. The presence of tunnels of different diameters and their branching detected by SEM images of the epoxy cast resembles the ichnogenus Iramena .
DISCUSSION
Both micro-CT and epoxy vacuum cast-embedding followed by SEM observations are widely regarded as very helpful and necessary tools for studying and visualizing the external and internal surfaces as well as the two-dimensional and three-dimensional (3D) architecture of tunnel systems left by bioeroders in various substrates (Tapanila, 2008, Wisshak, 2012), see Table 1. Nevertheless, both methods also have limitations, and using each method alone could make it difficult to diagnose and describe minute bioerosive structures as discussed below:
Material of hard substrate and borings filling. In general, most substrates can be examined using X-ray, so there are no restrictions for hard substrate in micro-CT device. The micro-CT can detect material of different densities if the contrast between the fossil material and the respective boring infilling is sufficient. For serpulid tubes and their infillings, the contrast was enhanced by using Al, Cu, or combined Al+Cu filters. A suitable sample size of specimens for micro-CT is approximately 0.5-10 cm. Significantly smaller samples are studied with nano-CT, and significantly larger samples are studied with common medical CT. However, these devices were not used in our study.
The vacuum embedding method is a fast and powerful tool for the study of bioerosion within calcareous substrates. The preparation of samples, including ultrasonic cleaning treatment as discussed above, usually precedes the resin infiltration and casting and subsequent chemical carbonate removal steps. This process is thus inaccessible for undissolvable silicified or pyritized substrates (Wisshak et al., 2017). This occurred with specimen NM O8727; it had a high degree of silicification, which led to incomplete dissolution of the walls in HCl and HF, thus making it unsuitable for the epoxy-casting methodology. The borings were identified using micro-CT, because the SEM image shows only short indeterminate non-branching shafts (Figure 4A). Vacuum casting is also unsuitable for boreholes infilled by a dense matrix. If boreholes are not empty, the resulting quality of epoxy casts is highly dependent on adequate porosity and permeability of the boring fill, since it must be impregnated by epoxy resin. Especially if the deepest and thinnest parts of macroborings are filled with residual fine-grained sediment impermeable by epoxy resin, complete casting of all of the borings is often not successful (Tapanila, 2008). In specimens CZ2 and NM O8727, epoxy resin did not infill all boreholes available, compared to what is seen in micro-CT reconstruction (Figure 4A, B), or filled their proximal parts only (Figure 3B). Dense carbonate inside the deepest part of some boreholes in CZ2 did not allow the resin to infiltrate the borings completely, so these structures were partly destroyed by subsequent dissolving in diluted acid. Boreholes in NM O8727 were partly filled within soluble silicified matrix. Specimen NM O8728 proved to be the ideal sample for epoxy castings. Its tube was filled with impermeable, acid-soluble carbonate matrix that prevented resin intrusion, as boreholes were empty, ready for resin impregnation. After filling and hardening the resin, the tube was longitudinally cut in half, and together with tube filler was decalcified in HCl. After this procedure, SEM images of the resulting cast provided a detailed three-dimensional view of the ichnological processes inside the entire serpulid tube (Figure 5A, C).
External and internal morphology of hard substrate. Micro-CT is useful in non-invasively visualizing gross internal and external morphological features in the µm to mm size range. This faciliated the taxonomic identification of serpulid specimen Cementula. In outer morphology, the tubes of Cementula are very similar to those of Spiraserpula, but Spiraserpula possesses internal longitudinal ridges on the inner surface of the tube wall (internal tube structures, see Pillai,1993; Pillai and ten Hove, 1994), while Cementula lacks them (Figure 3F).
Destructiveness. Because micro-CT allows non-invasive visual depicting of internal structures of various objects, it is suitable for valuable specimens, e.g., holotypes, paratype series, and other types that should be preserved for further studies (Abel et al., 2012). Non-destructivity is also needed for museum practice: once a sample enters a museum’s database, it becomes a subject of often strict regulations. The raw data obtained from micro-CT can be processed and visualized as virtual sections in diverse planes, three-dimensional views, animations in a full 360° range, or segmentation. Micro-CT produces so-called “virtual fossils” (Abel et al., 2012; Sutton et al. 2014). Once the specimen is X-rayed, the images obtained are stored in the computer and can be worked with further. If fossils are then destroyed during vacuum cast-embedding, they are at least preserved and remain accessible as virtual fossils.
Image resolution and three-dimensional animation. The most serious limits of micro-CT are related to resolution. The ideal smallest resolution for micro-CT is about 5 μm, which may limit studies on microborings as well as tiny macroborings (Heřmanová et al., 2020). Domichnia in serpulid polychaetes are smaller than those in bivalves, brachiopods, or echinoderms, which suggests that serpulids were mainly occupied by small, dwarfed but adult tracemakers (Mikuláš et al., 2022). Because resolution of less than 10 μm could not be effectively achieved for the serpulid samples, and we suspected that micro-CT did not fully detect possible details in the tube substrates, we decided for epoxy casting. Although vacuum cast-embedding producing polymer resin casts is a destructive technique, it is an excellent method to display the most delicate morphological features, because the SEM images of the resulting casts have significantly better resolution than micro-CT (Golubic et al., 1983; Wisshak, 2012; Heřmanová et al., 2020).
In specimen CZ2, both serpulid tube interiors were filled with epoxy resin and boreholes located beneath the epoxy infilling are obscured. However, epoxy casting provided detailed morphology of borings that were beyond the detection limit of the given spatial resolution of the micro-CT scan, and helped with ichnotaxonomic identification. These are tiny branching canals of Iramena (compare Figure 3C and 3E) and the finer apertural canals connecting the Entobia chambers to the tube surface in specimens CZ2 and NM O8728. Epoxy casting also helped in displaying microbioerosion (about 10 µm in diameter) just beneath the tube surface (Figure 3B). These structures were not detected using micro-CT, but are visible only in epoxy resin casts.
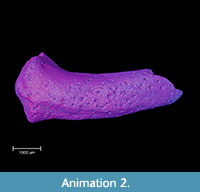
 Three-dimensional animations are a standard output of the micro-CT methodology. This technology is not limited to a two-dimensional view, so short movies showing complete internal structures or surfaces from all sides are often generated. In three-dimensional animation it is possible to simultaneously view both the internal and external structures of the fossil, and make some parts of it semi-transparent (see Animation 1 and Animation 2).
Three-dimensional animations are a standard output of the micro-CT methodology. This technology is not limited to a two-dimensional view, so short movies showing complete internal structures or surfaces from all sides are often generated. In three-dimensional animation it is possible to simultaneously view both the internal and external structures of the fossil, and make some parts of it semi-transparent (see Animation 1 and Animation 2).
 Hardware and software. Micro-CT equipment (hardware) is an expensive device, the price is not quantified here. The price depends on the agreement of the buyer and the seller. Basically, the price of a micro-CT depends on the types of samples that need scanning scanning and the results desired from the system. This determines the resolution needed for the appropriate micro-CT analysis, which determines the choice of X-ray source and detector. Micro-CT also makes serious demands on the attendant computer technology. Reconstruction systems must run on a powerful computer, and they produce very large datasets, bringing problems with long-term storage of such data (Paterson et al., 2014; Keklikoglou et al., 2019). In addition, there are legal restrictions on the operation of X-ray equipment in many countries.
Hardware and software. Micro-CT equipment (hardware) is an expensive device, the price is not quantified here. The price depends on the agreement of the buyer and the seller. Basically, the price of a micro-CT depends on the types of samples that need scanning scanning and the results desired from the system. This determines the resolution needed for the appropriate micro-CT analysis, which determines the choice of X-ray source and detector. Micro-CT also makes serious demands on the attendant computer technology. Reconstruction systems must run on a powerful computer, and they produce very large datasets, bringing problems with long-term storage of such data (Paterson et al., 2014; Keklikoglou et al., 2019). In addition, there are legal restrictions on the operation of X-ray equipment in many countries.
On the other hand, epoxy vacuum-embedding equipment can be acquired orders of magnitude more cheaply than micro-CT. Epoxy vacuum-embedding is followed by SEM observation. The purchase price of scanning electron microscope equipment is relatively high, and as with micro-CT equipment, depends on the agreement between the seller and the buyer. However, the software requirements for the epoxy vacuum-embedding method are significantly lower. Data production is much smaller, and arranging for long-term storage of images is usually trivial. In many institutions, it is possible to rent micro-CT and SEM equipment. Prices per hour of use of the device vary. Documenting one sample (in our case sample CZ2) on the micro-CT equipment took 21 hours (net working time). Making resin casts is much more time-consuming. Curing, cutting, and decalcifying the samples took 5 days, and subsequent SEM analysis another 2 hours.
CONCLUSIONS
Studying serpulid tubes as a hard substrate for sclerobionts shows that different imaging techniques bias the diagnosis and description of bioerosion. The result of micro-CT compared to vacuum cast-embedding of the same specimen show the benefits and limits of both methods. The main advantage of SEM images of casted specimens is the depiction of fine details, which helped in the correct taxonomic determination: bifurcation of Iramena in CZ2 or Entobia chambers with the finer apertural canals in CZ2 and NM O8728. The image resolution is much better than those obtained by micro-CT. The disadvantages of the vacuum cast-embedding method are its destructiveness and the restrictions caused by insolubility of the fossil specimens. It proved unsuitable for specimen NM O8727, which is silicified and therefore insoluble in acid. Moreover, vacuum casting requires empty borings or at least porous and permeable boring fills. Distal parts of some boreholes in CZ2 were filled with a dense carbonate matrix; they could not be impregnated with epoxy resin and were dissolved in acid. The non-destructive nature is the greatest benefit of micro-CT. It is suitable for rare and valuable specimens which cannot be replaced. Micro-CT also helped with the taxonomic identification of Cementula sp. (specimen CZ2) by visualizing internal morphological structures. Micro-CT is also able to display boreholes filled with dense matrix, if sufficient contrast exists between boring filling and substrate. Micro-CT has an advantage in allowing the object to be viewed in full 3D, including the ability to make three-dimensional animations and serial sections through the object. SEM of casted specimens can also provide three-dimensional images of a borehole system inside serpulid tubes, but only when vacuum casting is applied to specimens with empty borings impregnated by epoxy resin, while the tube interior is filled with dense carbonate matrix and subsequently dissolved in acid (specimen NM O8728). The disadvantage of micro-CT, apart from storage requirements of gigabytes per specimen, is the inability to achieve the resolution of SEM images of casted specimens. Although macroborings are easily recognizable on maximally magnified micro-CT images (long, cylindrical galleries of Maeandropolydora and unbranched shafts of Trypanites), they lack details smaller than 10 μm in diameter, which are available from SEM images (meandering and tangled microborings just beneath the serpulid tube surface, Entobia chambers, and bifurcation of Iramena). In practice, micro-CT, although non-destructive, is not an optimal method for investigation of μm-sized objects in mm- to cm-sized serpulid tubes, and epoxy casting thus plays an important role in visualizing and identifying of such tiny bioerosional structures. For pyritized or silicified substrates unsuitable for the cast-embedding technique, micro-CT is currently the only option for imaging their bioerosion structures. Our experience shows that both methods may complement and support each other.
ACKNOWLEDGEMENTS
This work was financially supported by Ministry of Culture of the Czech Republic (DKRVO 2019–2023/2.I.e, National Museum, 00023272) and Centre for Geosphere Dynamics (UNCE/SCI/006).This study was financially supported by the Czech Science Foundation (GACR 18-05935S) and by the institutional projects of the Czech Academy of Sciences, Institute of Geology (RVO 67985831).
REFERENCES
Abel, R.L., Laurini, C.R., and Richter, M. 2012. A palaeobiologist’s guide to ‘virtual’ micro-CT preparation. Palaeontologia Electronica, 15.2.6T. https://doi.org/10.26879/284
Amon, D.J., Sykes, D., Ahmed, F., Copley, J.T., Kemp, K.M., Tyler, P.A., Young, C.M., and Glover, A.G. 2015. Burrow forms, growth rates and feeding rates of wood-boring Xylophagaidae bivalves revealed by micro-computed tomography. Frontiers in Marine Science, 2(10):1-13. https://doi.org/10.3389/fmars.2015.00010
Benner, J.S., Ekdale, A.A., and de Gilbert, J.M. 2008. Enigmatic organisms preserved in early Ordovician macroborings, western Utah, USA, p. 55-64. In Wisshak, M. and Tapanila, L. (eds.), Current Developments in Bioerosion. Springer, Berlin, Heidelberg.
https://doi.org/10.1007/978-3-540-77598-0_3
Beuck, L., Vertio, A., Stepina, E., Karolczak, M., and Pfannkuche, O. 2006. Skeletal response of Lophelia pertusa (Scleractinia) to bioeroding sponge infestation visualised with micro-computed tomography. Facies, 53:157-176.
https://doi.org/10.1007/s10347-006-0094-9
Beuck, L., Wisshak, M., Munnecke, A., and Freiwald, A. 2008. A giant boring in a Silurian stromatoporoid analysed by computer tomography. Acta Palaeontologica Polonica, 53(1):149-160. https://doi.org/10.4202/app.2008.0111
Boekschoten, G.J. 1970. On bryozoan borings from the Danian at Fakse, Denmark, p. 43-48. In Crimes, T.P. and Harper, J.C. (eds.), Trace Fossils. Geological Journal, Special Issue 3, London.
Bromley, R.G. and D'Alessandro, A. 1984. The ichnogenus Entobia from the Miocene, Pliocene and Pleistocene of southern Italy. Rivista Italiana di Paleontologia e Stratigrafia, 90(2):227-296.
Bronn, H.G. 1837. Lethaea Geognostica, Abbildungen und Beschreibungen der für die Gebirgs-Formationen bezeichnendsten Versteinerungen 1. Schweizerbart, Stuttgart.
El-Hedeny, M. 2007. Encrustation and bioerosion on Middle Miocene bivalve shells and echinoid skeletons: paleoenvironmental implications. Revue de Paléobiologie, 26(2):381-389.
Färber, C., Titschack, J., Schönberg, Ch.H.L., Ehrig, K., Boos, K., Baum, D., Illerhaus, B., Asgaard, U., Bromley, R.G., Freiwald, A., and Wisshak, M. 2016. Long-term macrobioerosion in the Mediterranean Sea assessed by micro-computed tomography. Biogeosciences, 13:3461-3474. https://doi.org/10.5194/bg-13-3461-2016
Geinitz, H.B. 1875a. Vermes. Würmer, p. 282-287. In Geinitz, H.B. 1871-1875 (ed.), Das Elbthalgebirge in Sachsen. Erster Theil. Der untere Quader. Palaeontographica, 20(1), Cassel.
Geinitz, H.B. 1875b. Würmer, Krebse, Fische und Pflanzen. V. Würmer, p. 200-202. In Geinitz, H.B. 1872-1875 (ed.), Das Elbthalgebirge in Sachsen. Zweiter Theil. Der mittlere und obere Quader. Palaeontographica, 20(2), Cassel.
Glauert, A.M., Rogers, G.E., and Glauert, R.H. 1956. A new embedding medium for electron microscopy. Nature, 178:803. https://doi.org/10.1038/178803a0
Goldfuss, A. 1841. Petrefacta Germaniae 3(1):1-20, pls 166-171. Arnz and Co., Düsseldorf.
Golubic, S. 1983. Stromatolites, fossil and recent: a case history, p. 313-327. In Westbroek, P. and de Jong, E.W. (eds.), Biomineralization and biological metal accumulation. Springer, Dordrecht.
Golubic, S., Brent, G., and Le Campion, T. 1970. Scanning electron microscopy of endolithic algae and fungi using a multipurpose casting-embedding technique. Lethaia, 3(2):203-209. https://doi.org/10.1111/j.1502-3931.1970.tb01858.x
Heřmanová, Z., Bruthansová, J., Holcová, K., Mikuláš, R., Kočová Veselská, M., Kočí, T., Dudák, J., and Vohník, M. 2020. Benefits and limits of X-ray micro-computed tomography for visualization of colonization and bioerosion of shelled organisms. Palaeontologia Electronica, 23.2.23A. https://doi.org/10.26879/1048
Ippolitov, A.P., Vinn, O., Kupriyanova, E.K., and Jäger, M. 2014. Written in stone: history of serpulid polychaetes through time. Memoirs of Museum Victoria, 71:123-159.
http://doi.org/10.24199/j.mmv.2014.71.12
Jäger, M. 1983. Serpulidae (Polychaeta sedentaria) aus der norddeutschen höheren Oberkreide. Systematik - Stratigraphie - Ökologie. Geologisches Jahrbuch, Reihe A, 68:3-219.
Jäger, M. 2005. Serpulidae und Spirorbidae (Polychaeta sedentaria) aus Campan und Maastricht von Norddeutschland, den Niederlanden, Belgien und angrenzenden Gebieten. Geologisches Jahrbuch, Reihe A, 157:121-249.
Jäger, M. 2012. Sabellids and serpulids (Polychaeta sedentaria) from the type Maastrichtian, the Netherlands and Belgium. Scripta Geologica, Special Issue, 8:45-82.
Jäger, M. 2014. Serpuliden und Sabelliden, p. 57-81. In Niebuhr, B. and Wilmsen, M. (eds.), Kreide-Fossilien in Sachsen, Cretaceous fossils of Saxony, part 1. Geologica Saxonica. Senckenberg Gesellschaft für Naturforschung, Frankfurt am Main.
Jäger, M. and Kočí, T. 2007. A new serpulid, Placostegus velimensis sp. nov., from the Lower Turonian of the Bohemian Cretaceous Basin. Acta Geologica Polonica, 57(1):89-96.
https://geojournals.pgi.gov.pl/agp/article/view/9751
Keklikoglou, K., Faulwetter, S., Chatzinikolaou, E., Wils, P., Brecko, J., Kvaček, J., Metscher, B., and Arvanitidis, C. 2019. Micro-computed tomography for natural history specimens: a handbook of best practice protocols. European Journal of Taxonomy, 522:1-55.
https://doi.org/10.5852/ejt.2019.522
Kočí, T. 2008. Nové nálezy a předběžná zpráva o revizi serpulidních červů z lokalit Velim a Kaňk- Na Vrších (příbojová facie české křídové pánve). Vlastivědný zpravodaj Polabí, 39:207-238.
Kočí, T. 2010. The subgenus Septenaria Regenhardt, 1961 (Polychaeta: Serpulidae) from Lower Turonian (Upper Cretaceous) nearshore facies of the Bohemian Cretaceous Basin, Czech Republic. Journal of the National Museum Prague, Natural History Series, 179(10): 119-126.
Kočí, T. 2012. Sabellidae and Serpulidae (Polychaeta, Canalipalpata) from the locality Kaňk - Na Vrších in Kutná Hora (Upper Cenomanian - Lower Turonian, Bohemian Cretaceous Basin, the Czech Republic). Acta Musei Nationalis Pragae, Series B, Historia Naturalis, 68(1-2):7-14.
Kočí, T. and Jäger, M. 2015. Sabellid and serpulid worms (Polychaeta, Canalipalpata, Sabellida, Sabellidae, Serpulidae) from the rocky coast facies (Late Cenomanian) at Předboj near Prague. Acta Musei Nationalis Pragae, Series B, Historia Naturalis, 71(1-2):31-50.
https://doi.org/10.14446/AMNP.2015.31
Kočí, T., Jäger, M., and Morel, N. 2017. Sabellid and serpulid worm tubes (Polychaeta, Canalipalpata, Sabellida) from the historical stratotype of the Cenomanian (Late Cretaceous; Le Mans region, Sarthe, France). Annales de Paléontologie, 103(1):45-80.
https://doi.org/10.1016/j.annpal.2016.11.004
Kočová Veselská, M., Kočí, T., and Kubajko, M., 2014. Dynomenid crabs (Decapoda, Brachyura) and stalked barnacles (Cirripedia, Scalpelliformes) from upper Cenomanian-lower Turonian nearshore, shallow-water strata in the Bohemian Cretaceous Basin, Czech Republic. Scripta Geologica, 147:49-81.
Kočová Veselská, M., Kočí, T., Jäger, M., Mikuláš, R., Heřmanová, Z., Morel, N., and Šamánek, J. 2021. Sclerobionts on tubes of the serpulid Pyrgopolon (Pyrgopolon) deforme (Lamarck, 1818) from the upper Cenomanian of Le Mans region, France. Cretaceous Research, 125:104873. https://doi.org/10.1016/j.cretres.2021.104873
Košťák, M., Vodrážka, R., Frank, J., Mazuch, M., and Marek, J. 2010. Late Cretaceous nautilid beaks from near-shore/shallow water deposits of the Bohemian Cretaceous Basin (Czech Republic). Acta Geologica Polonica, 60(3):417-428.
Lee, S., Shi, G.R., Park, T.Y.S., Oh, J.R., Mii, H.S., and Lee, M. 2017. Virtual Palaeontology: the effects of mineral composition and texture of fossil shell and hosting rock on the quality of X-ray microtomography (XMT) outcomes using Palaeozoic brachiopods. Palaeontologia Electronica, 20.2.3T.
https://doi.org/10.26879/753
Lukeneder, A. 2012. Computed 3D visualisation of an extinct cephalopod using computer tomographs. Computers & Geosciences, 45:68-74.
https://doi.org/10.1016/j.cageo.2012.04.003
Lukeneder, S. and Lukeneder, A. 2011. Methods in 3D modelling of Triassic ammonites from Turkey (Taurus, FWF P22109-B17). Conferences Proceedings of the 14th International Association for Mathematical Geosciences, Salzburg, Austria, p. 496-505.
Lukeneder, A., Lukeneder, S., and Gusenbauer, C., 2014. Computed tomography of fossils and sulphide minerals from the Mesozoic of Turkey. Conferences Abstracts of the 5th Conference on Industrial Computed Tomography, Wels, Austria, 2 pp.
Maaløe, O. and Birch-Andersen, A. 1956. On the organization of the "nuclear material" in Salmonella typhimurium, p. 261-278. In Spooner, E.T.C. and Stocker, B.A.D. (eds.), Bacterial Anatomy, Sixth Symposium of the Society for General Microbiology, Cambridge University.
Madani, B., Ghenim, A.F., Ferré, B., Ayoub-Hannaa, W., Kočí, T., Adaci, M., Guendouz, M.L., Zaoui, D., Bendella, M., and Bensalah, M. 2022. Taxonomy, palaeoecology and palaeobiogeography of upper Cenomanian (Upper Cretaceous) oysters and plicatulids from southern Algeria. Historical Biology, 34(7):1274-1304.
https://doi.org/10.1080/08912963.2021.1974018
Mägdefrau, K. 1932. Über einige Bohrgänge aus dem Unteren Muschelkalk von Jena. Paläontologische Zeitschrift, 14(3):150-160. https://doi.org/10.1007/BF03041628
Mees, F., Swennen, R., Van Geet, M., and Jacobs, P. 2003. Applications of X-ray Computed Tomography in the Geosciences. Geological Society, London, Special Publications, 215:1-6. https://doi.org/10.1144/GSL.SP.2003.215.01.01
Mikuláš, R., Kočová Veselská, M., Kočí, T., Šamánek, J., Jäger, M., Heřmanová, Z., and Bruthansová, J. 2022. Domichnial borings in serpulid tube walls: Prosperous benthic assemblages in the Cretaceous of France and the Czech Republic. Frontiers in Ecology and Evolution, 10:882450. https://doi.org/10.3389/fevo.2022.882450
Nielsen, J.K. and Maiboe, J. 2000. Epofix and vacuum: an easy method to make casts of hard substrates. Palaeontologia Electronica, 3(1):2.
https://palaeo-electronica.org/2000_1/epofix/issue1_00.htm
Paterson, G.L.J., Sykes, D., Faulwetter, S., Merks, R., Ahmed, F., Hawkins, L.E., Dinley, J., Ball, A.D., and Arvanitidis, C. 2014. The pros and cons of using micro-computed tomography in gross and microanatomical assessments of polychaetous annelids. Memoirs of Museum Victoria, 71:237-246.
Pillai, T.G. 1993. A review of some Cretaceous and Tertiary serpulid polychaetes of the genera Cementula and Spiraserpula Regenhardt 1961, Laqueoserpula Lommerzheim 1979 and Protectoconorca Jäger 1983. Paläontologische Zeitschrift, 67(1-2):69-88.
https://doi.org/10.1007/BF02985871
Pillai, T.G. and ten Hove, H.A. 1994. On Recent species of Spiraserpula Regenhardt, 1961, a serpulid polychaete genus hitherto known only from Cretaceous and Tertiary fossils. Bulletin of the Natural History Museum London Zoology, 60(1):39-104.
Ramsdale, R. 2021. A possible serpulid tube worm of the genus Filograna from the upper Permian Cadeby Formation of South Yorkshire, UK. Proceedings of the Yorkshire Geological Society, 63(3):249-256. https://doi.org/10.1144/pygs2020-016
Reuss, A.E. 1845. Die Versteinerungen der böhmischen Kreideformation. Mit Abbildungen der neuen oder weniger bekannten Arten, Erste Abtheilung. Schweizerbart, Stuttgart.
Reuss, A.E. 1846. Die Versteinerungen der böhmischen Kreideformation. Mit Abbildungen der neuen oder weniger bekannten Arten, Zweite Abtheilung. Schweizerbart, Stuttgart.
Sanfilippo, R., Rosso, A., Reitano, A., Viola, A., and Insacco, G. 2018. New serpulid polychaetes from the Permian of western Sicily. Acta Palaeontologica Polonica, 63(3):579-584.
https://doi.org/10.4202/app.00448.2017
Schönberg, C.H.L. and Shield, G. 2008. Micro-computed tomography for studies on Entobia: transparent substrate versus modern technology, p. 147-163. In Wisshak, M. and Tapanila L. (eds.), Current Developments in Bioerosion. Springer, Berlin, Heidelberg.
https://doi.org/10.1007/978-3-540-77598-0_8
Seuss, B. and Nützel, A. 2019. Bioerosion in fossil cephalopods: a case study from the Upper Carboniferous Buckhorn Asphalt Quarry Lagerstätte, Oklahoma, USA. Facies, 65(2):6-17. https://doi.org/10.1007/s10347-018-0547-y
Silbiger, N.J., Guadayol, O., Thomas, F.I.M., and Donahue, M.J. 2016. A novel µCT analysis reveals different responses of bioerosion and secondary accretion to environmental variability. PLoS ONE, 11(4):e0153058. https://doi.org/10.1371/journal.pone.0153058
Sutton, M., Rahman, I., and Garwood, R. 2014. Techniques for Virtual Palaeontology. Wiley-Blackwell.
Tapanila, L. 2008. The medium is the message: imaging a complex microboring (Pyrodendrina cupra igen. n., isp. n.) from the early Paleozoic of Anticosti Island, Canada, p. 123-145. In Wisshak, M. and Tapanila L. (eds.), Current Developments in Bioerosion. Springer-Verlag, Berlin, Heidelberg. https://doi.org/10.1007/978-3-540-77598-0_7
ten Hove, H.A. and Kupriyanova, E.K. 2009. Taxonomy of Serpulidae (Annelida, Polychaeta): The state of affairs. Zootaxa, 2036:1-126. https://doi.org/10.11646/zootaxa.2036.1.1
Vinn, O. 2020. Biomineralization of Polychaete Annelids in the Fossil Record. Minerals, 10, 858:1-11. https://doi.org/10.3390/min10100858
Vinn, O., Jäger, M., and Kirsimäe, K. 2008a. Microscopic evidence of serpulid affinities of the problematic fossil tube ‘Serpula’ etalensis from the Lower Jurassic of Germany. Lethaia, 41:417-421. https://doi.org/10.1111/j.1502-3931.2008.00093.x
Vinn, O., ten Hove, H.A., and Mutvei, H. 2008b. On the tube ultrastructure and origin of calcification in sabellids (Annelida, Polychaeta). Palaeontology, 51(2):295-301.
https://doi.org/10.1111/j.1475-4983.2008.00763.x
Vinn, O., Mutvei, H., ten Hove, H.A., and Kirsimäe, K. 2008c. Unique Mg-calcite skeletal ultrastructure in the tube of the serpulid polychaete Ditrupa. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 248(1):79-89.
https://doi.org/10.1127/0077-7749/2008/0248-0079
Vinn, O., ten Hove, H.A., Mutvei, H., and Kirsimaee, K. 2008d. Ultrastructure and mineral composition of serpulid tubes (Polychaeta, Annelida). Zoological Journal of the Linnean Society, 154(4):633-650. https://doi.org/10.1111/j.1096-3642.2008.00421.x
Vinn, O. and Mutvei, H. 2009. Calcareous tube worms of the Phanerozoic. Estonian Journal of Earth Sciences, 58(4):286-296. https://doi.org/10.3176/earth.2009.4.07
Vodrážka, R., Bubík, M., Švábenická, L., and Žítt, J., 2013. Late Cretaceous (Upper Cenomanian - Lower Turonian) transgressive deposits near Kutná Hora and Kolín (Central Bohemia, Bohemian Cretaceous Basin). The Micropaleontological Society, Spring Meeting 2013, Excursion Guide. Charles University, Prague.
Voigt, E. 1965. Über parasitische Polychaeten in Kreide-Austern sowie einige andere in Muschelschalen bohrende Würmer. Palaeontologische Zeitschrift, 39:193-211.
https://doi.org/10.1007/BF02990164
Wisshak, M. 2012. Microbioerosion, In Knaust, D. and Bromley, R. (eds.), Trace fossils as indicators of sedimentary environments. Developments in Sedimentology, 64:213-243. Elsevier, Amsterdam.
https://doi.org/10.1016/B978-0-444-53813-0.00008-3
Wisshak, M., Titschack, J., Kahl, W.-A., and Girod, P. 2017. Classical and new bioerosion trace fossils in Cretaceous belemnite guards characterised via micro-CT. Fossil Record, 20(2):173-199. https://doi.org/10.5194/fr-20-173-2017
Ziegler, V. 1984. Family Serpulidae (Polychaeta, Sedentaria) from the Bohemian Cretaceous Basin. Acta Musei Nationalis Pragae, Series B, 39(4):213-254.
Žítt, J. 1992. Bored and mineralized limestone surfaces in the Upper Cretaceous of Bohemia. A preliminary report. Bulletin of the Czech Geological Survey, 67:109-115.
Žítt, J. and Nekvasilová, O. 1989. Paleontologicko-geologická charakteristika navrhovaného CHPV Karlov (Kutná Hora). Bohemia Centralis, 18:15-40.
Žítt, J. and Nekvasilová, O. 1994. Běstvina u Ronova nad Doubravou - pozoruhodný výskyt spodnoturonských fosílií v příbřežních sedimentech české křídové pánve (Kolínská litofaciální oblast). Bohemia centralis, 23:23-30.
Žítt, J. and Nekvasilová, O. 1996. Epibionts, their hard-rock substrates, and phosphogenesis during the Cenomanian-Turonian boundary interval (Bohemian Cretaceous Basin, Czech Republic). Cretaceous Research, 17(6):715-739. https://doi.org/10.1006/cres.1996.0038
Žítt, J., Nekvasilová, O., Bosák, P., Svobodová, M., Štemproková-Jírová, D., and Šťastný, M. 1997. Rocky coast facies of the Cenomanian-Turonian Boundary interval at Velim (Bohemian Cretaceous Basin, Czech Republic). Parts 1-2. Bulletin of the Czech Geological Survey, 72(1):83-102 (First Part); 72(2):141-156 (Second Part).
Žítt, J., Nekvasilová, O., Hradecká, L., Svobodová, M., and Záruba, B. 1998. Rocky coast facies of the Unhošť-Tursko High (Late Cenomanian-Early Turonian, Bohemian Cretaceous Basin). Acta Musei Nationalis Pragae, Series B, 54:79-116.
Žítt, J., Vodrážka, R., Hradecká, L., Svobodová, M., and Zágoršek, K. 2006. Late Cretaceous environments and communities as recorded at Chrtníky (Bohemian Cretaceous Basin, Czech Republic). Bulletin of Geosciences, 81(1):43-79.
https://doi.org/10.3140/bull.geosci.2006.01.043
Žítt, J., Vodrážka, R., Hradecká, L., Svobodová, M., Šťastný, M., and Švábenická, L. 2015. Depositional and palaeoenvironmental variation of lower Turonian nearshore facies in the Bohemian Cretaceous Basin, Czech Republic. Cretaceous Research, 56:293-315.
https://doi.org/10.1016/j.cretres.2015.05.007
