

 Underwater drunken forest: Changes in growth direction and ornamentation in Conularia fragilis Barrande, 1867 (Lower Devonian, Czech Republic)
Underwater drunken forest: Changes in growth direction and ornamentation in Conularia fragilis Barrande, 1867 (Lower Devonian, Czech Republic)
Article number: 27.3.a54
https://doi.org/10.26879/1414
Copyright Palaeontological Association, November 2024
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Supplementary Figure
Submission: 2 June 2024. Acceptance: 29 October 2024.
ABSTRACT
A very high percentage of specimens of Conularia fragilis Barrande, 1867 from the Lower Devonian Prague Formation of Bohemia (Czech Republic) show appreciable curvature or bending of the steeply pyramidal periderm, which in most other conulariids tends be straight. Curved or bent sections of the periderm in C. fragilis are invariably associated with substantial departure from normal patterns of the external ornamentation (transverse ribs) of the four faces. Based on these observations and on data on the microstructure and composition of the conulariids, we conclude that bending of the periderm occurred while the conulariids were alive. As in curved or bent rugose corals from the same formation, bending in the conulariids probably involved preferential addition of new peridermal material along that part of the oral growth margin on the outside of the curve and may have occurred in response to changes in ambient currents or shifting of a sandy (crinoidal) substrate distad or within crinoid-stromatoporoid bioherms.
Jana Bruthansová. Department of Paleontology, National Museum, Cirkusová 1740, 193 00, Prague 9, Czech Republic; Department of Geology and Palaeontology, Faculty of Science, Charles University, Albertov 6, 128 43 Prague 2, Czech Republic. jana.bruthansova@nm.cz (corresponding author)
Jiří Bruthans. Department of Hydrogeology, Engineering Geology and Applied Geophysics, Faculty of Science, Charles University, Albertov 6, 128 43 Prague 2, Czech Republic. bruthans@natur.cuni.cz
Jana Schweigstillová. Department of Applied Rock Mechanics, Institute of Rock Structure and Mechanics of The Czech Academy of Sciences, v. v. i., V Holešovičkách 94/41, 182 09 Praha 8, Czech Republic. jana@irsm.cas.cz
Heyo Van Iten. Department of Geology, Hanover College, Hanover, Indiana USA 47243 and Department of Invertebrate Paleontology, Cincinnati Museum Center, 1301 Western Avenue, Cincinnati, Ohio USA. vaniten@hanover.edu
Keywords: Conulariida; Medusozoa; paleoecology; microstructure; morphogenesis
Final citation: Bruthansová, Jana, Bruthans, Jiří, Schweigstillová, Jana, and Van Iten, Heyo. 2024. Underwater drunken forest: Changes in growth direction and ornamentation in Conularia fragilis Barrande, 1867 (Lower Devonian, Czech Republic). Palaeontologia Electronica, 27(3):a54.
https://doi.org/10.26879/1414
palaeo-electronica.org/content/2024/5375-underwater-drunken-forest
Copyright: November 2024 Palaeontological Association.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
Conulariids are an extinct order (Conulariida) of sessile benthic, polypoid medusozoan cnidarians (Van Iten et al., 2006a) ranging from the uppermost Ediacaran to the topmost Triassic (Van Iten et al., 2006a, 2014a, 2016; Lucas, 2012; Amorin et al., 2020; Leme et al., 2022). The group is most diverse in the Ordovician System, steadily decreasing in generic diversity following the End Ordovician Mass Extinction Event (Van Iten and Vyhlasová, 2004; Leme et al., 2010; Van Iten et al., 2014b, 2020). In the Czech Republic (Prague Basin), the diversity of conulariids mirrors the global trend, reaching a maximum in the Middle Ordovician (Brabcová, 2001; Van Iten and Vyhlasová, 2004) and then declining to a total of just nine species in the Devonian (Brabcová, 2001; Mergl et al., 2016).
Conulariids are characterized by an elongate, steep pyramidal periderm usually having four faces and sulcate corners. The microstructure and growth of the conulariid periderm have been treated in many publications (e.g., Bouček and Ulrich, 1939; Van Iten et al., 2006b, 2006c; John et al., 2010; Robson and Young, 2013; Ford et al., 2016; Mergl et al., 2016; Kröger et al., 2021; Miller et al., 2022; Vinn, 2022). Thanks mainly to scanning electron imaging of specimens etched in dilute HCl (Ford et al., 2016), is it now known that the conulariid periderm is composed of alternating apatitic and organic microlamellae measuring approximately 1 to 3 μm thick (Van Iten, 1992; Ford et al., 2016). The apertural end of the periderm may be open, or it may be partially or fully closed by inwardly folded, lobate, or triangular lappets. Where the opposite, or apical end of the periderm is preserved, it consists of a minute, subconical holdfast that is attached, in some cases, to hard biological substrates (Vinn et al., 2019). Bohemian specimens are truncated at varying distances above the apex (the closed aperture is preserved in some specimens), and many of them terminate in an outwardly convex apical wall, or schott, which may have been produced in response to mechanical severance of the originally sessile polyp (Van Iten, 1991; Van Iten et al., 2020; Bruthansová et al., 2022). None have yet been documented in a demonstrably original life orientation and relationship to their substrate.
The purpose of the present paper is to describe and interpret a large sample (89 specimens) of Conularia fragilis Barrande, 1867, from Lower Devonian (Pragian) limestones in the Prague Basin in Bohemia. Conularia fragilis is the most common of the nine species of conulariids collectively described and figured by Barrande (1867), Bouček (1928, 1939), and Mergl et al. (2016) from the Devonian of the Czech Republic. It is restricted to the Koněprusy Limestone of the Prague Formation, which consists in large part of biohermal and biodetrital limestones. Most of the specimens studied exhibit pronounced curvature or bending of the periderm together with abnormal patterns of external ornamentation. Such bending is well exemplified by a single, nearly complete specimen illustrated by Bouček (1928, plate V, fig. 8) and collected from a “yellowish crystalline limestone” from “Koněprusy”. Detailed analysis of many such specimens may provide additional insights into the morphogenesis of the conulariid periderm and, potentially, the paleoecology of conulariids.
Geological and Paleontological Settings
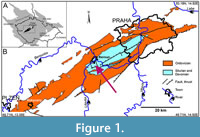 The studied conulariid specimens are from limestone deposits in the Prague Basin (Figure 1), which extends about 110 km from just east of Prague to the city of Pilsen. The Prague Basin originated as a rift basin (Havlíček, 1981, 1998) in which subsidence and sediment accumulation took place from the Early Ordovician (Tremadocian) to the Middle Devonian (Givetian). The basin is now a folded and faulted denudation relic. The Devonian sequence in the Prague Basin consists of six formations collectively ranging from Lochkovian to Givetian in age (Figure 2) (Chlupáč, 1998). The six rock units consist predominantly of carbonates, with Middle Devonian paleomagnetic signatures recording the northward drift of the basin from high southern to subtropical southern (ca. 10-20°) paleolatitudes (Krs et al., 1988, 1997; Patočka et al., 2003). Gradual deepening of the Prague Basin is recorded in the Lower Devonian (Pragian) Prague Formation (Figure 2), which has
The studied conulariid specimens are from limestone deposits in the Prague Basin (Figure 1), which extends about 110 km from just east of Prague to the city of Pilsen. The Prague Basin originated as a rift basin (Havlíček, 1981, 1998) in which subsidence and sediment accumulation took place from the Early Ordovician (Tremadocian) to the Middle Devonian (Givetian). The basin is now a folded and faulted denudation relic. The Devonian sequence in the Prague Basin consists of six formations collectively ranging from Lochkovian to Givetian in age (Figure 2) (Chlupáč, 1998). The six rock units consist predominantly of carbonates, with Middle Devonian paleomagnetic signatures recording the northward drift of the basin from high southern to subtropical southern (ca. 10-20°) paleolatitudes (Krs et al., 1988, 1997; Patočka et al., 2003). Gradual deepening of the Prague Basin is recorded in the Lower Devonian (Pragian) Prague Formation (Figure 2), which has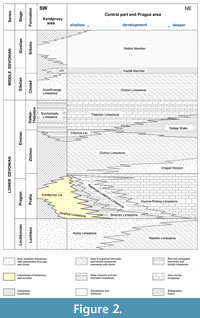 been subdivided into five members ranging in origin from proximal marine near the base of the unit (Koněprusy, Slivenec, and Vinařice limestones) to distal or deep-water marine (Řeporyje and Dvorce-Prokop limestones; see Chlupáč, 1998, fig. 5) near the top (Chlupáč, 1998). The 89 study specimens were collected from the Koněprusy Limestone near the village of Koněprusy (Figure 3) in the northwestern portion of the Prague Basin. Aspects of the stratigraphy and depositional environments of Devonian strata in the Koněprusy area have been discussed by Hladil and Slavík (1997), Hladil (1997, 1998), Janoušek et al. (2000, 2001), Chlupáč (2003), Slavík (2004), Slavík and Hladil (2007), Koptíková et al. (2010), Weinerová et al. (2017), Slavík and Hladil (2020), and Mergl (2022). The Koněprusy Limestone measures up to 150 m in thickness and consists of white to light grey, bioclastic crinoidal limestones and bioherms. The bioherms consist mainly of crinoids with massive stems and root systems (e.g., Pernerocrinus Bouška, 1946 and Torrocrinus Prokop and Petr, 1991), calcareous algae, and stromatoporoids (Chlupáč, 1998) along with minor tabulate and rugose corals. The bioherms also contain stromatolite-like, laminated stromatactis structures and localized accumulations of trilobite debris filling depressions and holes in the reef framework. The bioherms pass laterally into mostly unsorted, bioclastic material (size 1-20 cm, large, coarse) consisting of diverse crinoids, brachiopods, fenestellid bryozoans, rugose corals, algae, gastropods, and bivalves. Moving away from the reef cores, in the vicinity of Zlatý kůň, the amount of well-sorted crinoidal limestone increases (up to several cm large, fine), and stromatoporoids, algae, colonial anthozoans, and massive crinoid stems become absent (Plešivec Quarry area near Měňany) (Turek, personal commun., 2023). From Koněprusy towards the eastern and north-eastern portions of the basin, the proportion of coarse bioclastic material decreases with increasing water depth, and the proportion of fine-grained, silty carbonate material increases until it becomes the dominant sediment (Turek, personal commun., 2023).
been subdivided into five members ranging in origin from proximal marine near the base of the unit (Koněprusy, Slivenec, and Vinařice limestones) to distal or deep-water marine (Řeporyje and Dvorce-Prokop limestones; see Chlupáč, 1998, fig. 5) near the top (Chlupáč, 1998). The 89 study specimens were collected from the Koněprusy Limestone near the village of Koněprusy (Figure 3) in the northwestern portion of the Prague Basin. Aspects of the stratigraphy and depositional environments of Devonian strata in the Koněprusy area have been discussed by Hladil and Slavík (1997), Hladil (1997, 1998), Janoušek et al. (2000, 2001), Chlupáč (2003), Slavík (2004), Slavík and Hladil (2007), Koptíková et al. (2010), Weinerová et al. (2017), Slavík and Hladil (2020), and Mergl (2022). The Koněprusy Limestone measures up to 150 m in thickness and consists of white to light grey, bioclastic crinoidal limestones and bioherms. The bioherms consist mainly of crinoids with massive stems and root systems (e.g., Pernerocrinus Bouška, 1946 and Torrocrinus Prokop and Petr, 1991), calcareous algae, and stromatoporoids (Chlupáč, 1998) along with minor tabulate and rugose corals. The bioherms also contain stromatolite-like, laminated stromatactis structures and localized accumulations of trilobite debris filling depressions and holes in the reef framework. The bioherms pass laterally into mostly unsorted, bioclastic material (size 1-20 cm, large, coarse) consisting of diverse crinoids, brachiopods, fenestellid bryozoans, rugose corals, algae, gastropods, and bivalves. Moving away from the reef cores, in the vicinity of Zlatý kůň, the amount of well-sorted crinoidal limestone increases (up to several cm large, fine), and stromatoporoids, algae, colonial anthozoans, and massive crinoid stems become absent (Plešivec Quarry area near Měňany) (Turek, personal commun., 2023). From Koněprusy towards the eastern and north-eastern portions of the basin, the proportion of coarse bioclastic material decreases with increasing water depth, and the proportion of fine-grained, silty carbonate material increases until it becomes the dominant sediment (Turek, personal commun., 2023).
MATERIAL AND METHODS
Material
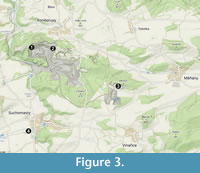 Nearly all 89 specimens of Conularia fragilis were obtained from museum collections dating from the nineteenth or early twentieth centuries. All are now housed in the collections of the National Museum in Prague (collection number prefix NML or NMS). The specimens are incomplete, having been truncated well above the apex and below the aperture, and preserve peridermal material. None of the conulariids were collected in situ, and therefore, their original orientations in the outcrop and the specific localities from which they were collected are unknown. All are labelled with the name Koněprusy, which covers a large area including the village of Koněprusy and encompassing multiple quarries (Figure 3). Owing to the absence of host rock material and diagnostic facies indicators within the peridermal cavity of the conulariids, the facies provenance of the conulariids likewise cannot be established.
Nearly all 89 specimens of Conularia fragilis were obtained from museum collections dating from the nineteenth or early twentieth centuries. All are now housed in the collections of the National Museum in Prague (collection number prefix NML or NMS). The specimens are incomplete, having been truncated well above the apex and below the aperture, and preserve peridermal material. None of the conulariids were collected in situ, and therefore, their original orientations in the outcrop and the specific localities from which they were collected are unknown. All are labelled with the name Koněprusy, which covers a large area including the village of Koněprusy and encompassing multiple quarries (Figure 3). Owing to the absence of host rock material and diagnostic facies indicators within the peridermal cavity of the conulariids, the facies provenance of the conulariids likewise cannot be established.
Methods
The studied conulariids were examined using optical microscopy (Olympus SZX12 binocular microscope) and photographed under low angle illumination using a Nikon EOS 6D digital camera. All conulariids greater than 30 mm in length (54 specimens) were photographed in orientations where the curvature or bending of the periderm was maximally apparent. From these photographs, the rate of expansion of the periderm along the longitudinal axis was estimated, as was whether the bending was gradual or abrupt. To establish the chemical composition of the conulariids and to obtain information bearing on their bending and malformed ornament, some of the conulariids were sectioned and analyzed using scanning electron microscopy (SEM) and energy dispersive X-ray spectroscopy (EDS) in the Institute of Rock Structure and Mechanics of the Academy of Sciences of the Czech Republic in Prague. Sectioned specimens were cut perpendicular or parallel to the longitudinal axis and following air drying were impregnated with SpeciFix Resin epoxy resin before high-speed polishing with 0.5-micron aluminum oxide powder. After ultrasonic cleaning in deionized water the polished surfaces were coated with gold. The specimens were imaged and analyzed using a QUANTA 450 (FEI) scanning electron microscope equipped with a photomultiplier detector (Centaurus PMD) for backscattered electrons (BSE). This work was carried out under a high vacuum using BSE to detect differences in chemical composition. Powder X-ray diffraction (XRPD) data for small pieces of extracted peridermal material were collected using a PANalytical X´Pert Pro diffractometer operating at 40 kV and 30 mA, with a secondary monochromator producing CuKα1,2 radiation and a X´Celerator detector (X-ray diffraction Laboratory, Institute of Geochemistry, Mineralogy and Mineral Resources, Faculty of Science, Charles University, Prague). Identification of phases was achieved using the High-Score search-match algorithm with PDF-2 (ICDD) database.
To examine the peridermal cavity, micro-computed tomographic analysis was performed using the X-ray micro-tomography SkyScan 1172 (Bruker) instrument in the Natural Museum in Prague. This instrument produces multiple X-ray “shadow” transmission images at different angles while the object rotates. From these shadow images, cross-sections of the conulariids were constructed using N-Recon Version 2.0.0 software to create a three-dimensional visualization of both the exterior and interior surfaces of the periderm. Digital images were created using Avizo (Version: 2020.3) software, and information thus revealed was processed using the volume rendering and ortho-slice modes for surfaces, iso-surfaces, and cross-sections. The X-ray microfocus tube was operated at 100 µA and 100 kV with a 1 mm Cu filter. The rotation step was 0.3° (one frame per step) through 360°, and the average image pixel size was 27.08 µm
RESULTS
Preservation and Irregularities in Skeletal Growth
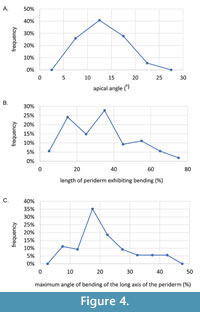 The studied conulariids exhibit four equally wide faces, with the two corners bordering any one face diverging at approximately 5 to 20° (Figure 4A-C and Supplementary Figure 1). In transverse cross sections the specimens are square or rhombic, and there are no terminal or internal schotts (apical walls) or longitudinal carinae (Figure 5). The largest specimen measures 127 mm in length, but it is truncated well above the apex and so its original length may have been about 200 mm. All specimens exhibit low, narrow, finely nodose transverse ribs, spaced along the longitudinal axis from 8 to 15 per 5 mm, which arch towards the apertural end of the periderm and cross the facial midline without interruption.
The studied conulariids exhibit four equally wide faces, with the two corners bordering any one face diverging at approximately 5 to 20° (Figure 4A-C and Supplementary Figure 1). In transverse cross sections the specimens are square or rhombic, and there are no terminal or internal schotts (apical walls) or longitudinal carinae (Figure 5). The largest specimen measures 127 mm in length, but it is truncated well above the apex and so its original length may have been about 200 mm. All specimens exhibit low, narrow, finely nodose transverse ribs, spaced along the longitudinal axis from 8 to 15 per 5 mm, which arch towards the apertural end of the periderm and cross the facial midline without interruption.
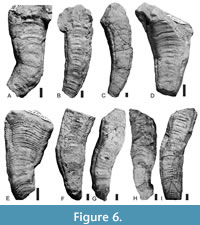
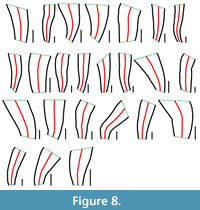
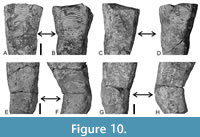
All specimens measuring at least 30 mm in length exhibit one or (less frequently) two bends along the long axis (Figure 6, Figure 7, Figure 8). The bending ranges from gentle to pronounced (Figure 7, Figure 8, Figure 9), with the total angle of bending (measured on the straight portions of the conulariids below and above the zone of bending) ranging from 7 to 45°, though with most specimens (n = 25) bending at about 20° (Figure 4C). Both on the outside and the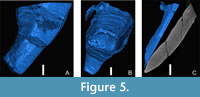 inside of a given bend, the transverse ribs are crowded, with approximately twice as many transverse ribs per 5 mm than below (adapically) or above (adaperturally) the bend. Additionally, the transverse ribs may be distorted, being irregularly undulatory and/or terminating well short of the nearest corner (Figure 10A-H). Segments that exhibit gradual bending constitute anywhere from 10 to 70% of the total preserved length of the conulariids (Figure 6, Figure 7, Figure 8). The square or rhombic transverse cross section of the periderm is preserved through the bend. Most of the specimens show a single bend, but 10 of them exhibit two bends (Figure 9F-G). In the latter specimens, the oral-most bend restores the original direction of extensional growth of the periderm. Because the conulariids are truncated above the apex and below the aperture, it is difficult to determine with certainty in which portion of the skeleton (apical, middle, or apertural) the observed bending is most frequent. Nevertheless, it appears from the size and shape of the specimens that bending occurs most frequently in the apical portion, with other specimens being bent in the middle or upper third.
inside of a given bend, the transverse ribs are crowded, with approximately twice as many transverse ribs per 5 mm than below (adapically) or above (adaperturally) the bend. Additionally, the transverse ribs may be distorted, being irregularly undulatory and/or terminating well short of the nearest corner (Figure 10A-H). Segments that exhibit gradual bending constitute anywhere from 10 to 70% of the total preserved length of the conulariids (Figure 6, Figure 7, Figure 8). The square or rhombic transverse cross section of the periderm is preserved through the bend. Most of the specimens show a single bend, but 10 of them exhibit two bends (Figure 9F-G). In the latter specimens, the oral-most bend restores the original direction of extensional growth of the periderm. Because the conulariids are truncated above the apex and below the aperture, it is difficult to determine with certainty in which portion of the skeleton (apical, middle, or apertural) the observed bending is most frequent. Nevertheless, it appears from the size and shape of the specimens that bending occurs most frequently in the apical portion, with other specimens being bent in the middle or upper third.
 |
 |
 |
 |
 |
 |
Microstructure and Composition
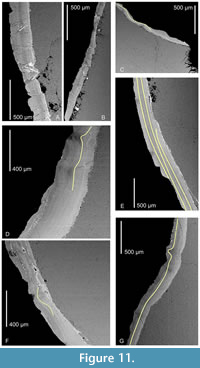
The periderm of Conularia fragilis ranges from 200-400 µm in thickness and consists of numerous, microscopic lamellae (microlamellae; Ford et al., 2016) measuring approximately 1-3 µm thick. As seen in backscattered electron mode the microlamellae are alternately light and dark (Figure 11A-G). The lighter-colored laminae are fluorine-rich and contain more Ca and P than the darker ones, which appear to be rich in organic matter. The microlamellae generally parallel each other, gradually and smoothly widening within the transverse ribs and thus making the periderm thicker there than between the ribs. Importantly, the microlamellae continue without disruption through curved or bent portion(s) of the periderm, and thus there is no internal evidence of injury or interruption of the growth of the periderm. Small cracks crosscutting the microlamellae and filled (in some cases) with diagenetic iron oxides or apatite clearly are post-morten in origin, indicating that the originally pliable periderm had become brittle.
DISCUSSION
Origin of the Peridermal Bending
Previous authors (e.g., Ford et al., 2016; Mergl et al., 2016) have agreed that the growth of the conulariid periderm involved accretion of new material along the apertural (oral) margin and thickening of the periderm by accretion of whole lamellae to its inner surface. Thus, the results of the present study suggest that bending of the periderm was brought about by preferential production of new peridermal material along that part of the apertural margin on the outside of a given bend. The bending clearly is neither tectonic nor diagenetic in origin (again, the microstructure of the periderm is unaffected by it), nor is there any evidence suggesting that it was associated with repair of damage caused by durophagous predation (see illustrations of healed conulariid injuries and growth abnormalities in Babcock et al., 1987; Mapes et al., 1989; Van Iten, 1992; Van Iten et al., 2022). Preferential addition of new periderm may have occurred at the oral edge of a single, outermost lamella or set of lamellae, followed later by accretion of additional lamellae to the inner surface of that or those lamellae involved in the formation of the bend. Associated with this process was disruption of normal patterns (spacing and geometry) of the transverse ribbing, which patterns were restored once the new growth direction was established. The cause of the observed bending of the periderm is unclear, but by analogy with solitary corals it may have taken place in response to changes in ambient currents or shifting of the conulariids’ substrate, which may have consisted of crinoid debris (Figure 12). Thus, in present-day subaqueous environments, ambient currents and substrate movement affect the body shapes of diverse, sessile benthic organisms, including various algae, corals, sponges, and bryozoans. All these organisms can modify their growth direction in response to changes in the prevailing water currents, enabling them to feed more efficiently and/or to deal more effectively with drag (see e.g., Jebram, 1970; Koehl, 1999; Denny and Gaylord, 2002; Boller and Carrington, 2006; Martone and Denny, 2008; Infantes et al., 2011; Luhar and Nepf, 2011; Rummel, 2014; Hays, 2017; Falcucci et al., 2021). In the Koněprusy Limestone, the animals most like the conulariids probably were the solitary rugose corals, many of which likewise are curved or bent (Figure 9E, H-J). These possibly semi-erect or recumbant, semi-infaunal polyps are thought to have changed the direction of extensional growth to maintain an optimal life position for feeding and reproduction (Neuman, 1988; Scrutton, 1998; Sorauf, 2001; Berkowski, 2012; Adomat et al., 2016). Hence, if analogy with horn corals is appropriate in the present case, it is tempting to suggest that like these animals, Conularia fragilis may have been semi-erect or even recumbent, changing its growth direction continuously (if, say, recumbent) or suddenly (if, say, semi-erect and subject to toppling) in order to maintain an optimal life posture relative to a mobile substrate.
Associated with this process was disruption of normal patterns (spacing and geometry) of the transverse ribbing, which patterns were restored once the new growth direction was established. The cause of the observed bending of the periderm is unclear, but by analogy with solitary corals it may have taken place in response to changes in ambient currents or shifting of the conulariids’ substrate, which may have consisted of crinoid debris (Figure 12). Thus, in present-day subaqueous environments, ambient currents and substrate movement affect the body shapes of diverse, sessile benthic organisms, including various algae, corals, sponges, and bryozoans. All these organisms can modify their growth direction in response to changes in the prevailing water currents, enabling them to feed more efficiently and/or to deal more effectively with drag (see e.g., Jebram, 1970; Koehl, 1999; Denny and Gaylord, 2002; Boller and Carrington, 2006; Martone and Denny, 2008; Infantes et al., 2011; Luhar and Nepf, 2011; Rummel, 2014; Hays, 2017; Falcucci et al., 2021). In the Koněprusy Limestone, the animals most like the conulariids probably were the solitary rugose corals, many of which likewise are curved or bent (Figure 9E, H-J). These possibly semi-erect or recumbant, semi-infaunal polyps are thought to have changed the direction of extensional growth to maintain an optimal life position for feeding and reproduction (Neuman, 1988; Scrutton, 1998; Sorauf, 2001; Berkowski, 2012; Adomat et al., 2016). Hence, if analogy with horn corals is appropriate in the present case, it is tempting to suggest that like these animals, Conularia fragilis may have been semi-erect or even recumbent, changing its growth direction continuously (if, say, recumbent) or suddenly (if, say, semi-erect and subject to toppling) in order to maintain an optimal life posture relative to a mobile substrate.
Unfortunately, all the conulariid specimens examined in the present study came from old collections, and therefore we could not determine whether they occurred in their original life orientation (erect and with the apertural end up) in the outcrops or had undergone transport prior to final burial. Previously, Simões et al. (2000, 2003) and Rodrigues et al. (2003) investigated the taphonomy of Devonian conulariids from Brazil, some of which exhibit broad transverse folding (undulation or wrinkling) of the faces like that displayed by some of the conulariids examined in the present study (e.g., Figure 5B, Figure 6B, 6I). In their material, Simões et al. (2003) observed the folding in erect specimens preserved in situ and interpreted it as a taphonomic artifact of vertical compaction of the host sediment following final burial of the conulariids. Because the Czech material lacks information on the original orientation of the conulariids in the outcrops, we could not evaluate the hypothesis that the observed folding in these specimens is a taphonomic artifact (as opposed to a primary anatomical feature).
Finally, we observed bending and irregular ornamentation in conulariids other than Conularia fragilis, namely Paraconularia planicostata (Dawson, 1868) from the Upper Mississippian of southwestern Newfoundland and Nova Scotia (see Van Iten et al., 2024, fig. 2), in Archaeoconularia insignis (Barrande, 1867) from the Upper Ordovician of Bohemia, and in Archaeoconularia consobrina (Barrande, 1855) from the Upper Ordovician of Morocco (Figure 9A-D). The possible causes of bending in these conulariids is a problem requiring further investigation, but it is important to note here that the phenomenon may be widespread taxonomically and thus indicative of fundamental growth processes common to the group.
Facies Provenance
As noted above in the introduction, we were not able to determine the facies provenance of the conulariids examined in the present study. Be that as it may, based on direct examination of multiple exposures of the Koněprusy Limestone and on discussions with colleagues having detailed knowledge of the unit, we think that the most plausible hypothesis is that Conularia fragilis inhabited relatively sheltered areas within the body of the reefs and/or on distal flanking sediments, in both cases among dense growths of calcareous green algae that may have acted to shelter the conulariids against current action (Figure 12). We have found fragmentary conulariid specimens at a few localities, for example the active quarries at Čertovy schody, Zlatý Kůň, and Cisař quarry, but the conulariid is a very rare faunal element, and no clear conclusions may be drawn from such a small sample.
Independent sedimentological evidence from these localities indicates high energy conditions on bioherm slopes, in waters up to 10 m deep (Bourillot et al., 2009; Denayer and Aretz, 2012; Duval-Arnould et al., 2024). However, such high energy environments are not generally suitable for larval attachment, and indeed larval supply is one of the principal factors determining the establishment, structure, and diversity of sessile benthic assemblages on reefs and bioherms (Bento et al., 2017). Thus, a more suitable environment for the attachment and further growth of conulariid larvae may have been sheltered, topographical heterogeneities within the bodies of the reefs, which in modern settings provide cryptic spaces that can act as refugia for coral and other larvae (e.g., Gallagher and Doropoulos, 2017; Martínez-Quintana et al., 2023). Sheltered algal meadows analogous to modern concentrations of seaweed and green alga may also have been favorable for conulariids (see e.g., Van Tussenbroek, 2011; Di Martino and Taylor, 2014; Wang et al., 2014; McNeil et al., 2021; East et al., 2023). Lastly, and again by analogy with solitary rugose corals, as well as by analogy with Conularia aff. desiderata Hall, 1847, from the Middle Devonian of New York State (USA) (Van Iten et al., 2013), we hypothesize that C. fragilis was semi-infaunal and oriented in life with the oral end facing directly or obliquely upward. The conulariid larvae probably attached to hard, biological substrates such as echinoderm ossicles, though, again, in relatively low energy settings where sudden shifting of the substrate was relatively infrequent, and where subsequent extensional growth of the conulariids was sufficiently rapid to prevent smothering by shifting or aggrading sediment.
CONCLUSIONS
Gross morphological and microstructural analyses of curved or bent, abnormally ornamented specimens of Conularia fragilis (Barrande, 1867), all from the Middle Devonian Koněprusy Limestone of the Prague Formation in Bohemia (Prague Basin), indicate that the bending occurred during the lifetimes of the conulariids, possibly in response to changes in ambient current action and/or to shifting of a (presumably) mobile sandy substrate. Similar bending is exhibited by solitary rugose corals from the same limestone unit, and in both cases was achieved through preferential addition of new skeletal material along one side of the oral (growth) margin. During the period of adjustment in growth direction, the spacing and relief of the transverse ribs along the bent or curved section of the conulariids changed markedly from the previous (pre-bending) stage, becoming “normal” again after establishment of the new and final growth direction. Owing in part to the lack of host rock matrix, the facies provenance of the bent conulariids is not known precisely, but it may have been sheltered portions of vuggy, shallow-water bioherm cores and/or deeper-water flank sediments farther from the paleoshoreline. Lastly, our results reinforce previously known similarities in anatomy and growth between conulariids and other, polypoid cnidarians.
ACKNOWLEDGEMENTS
We thank Š. Manda, F. Vacek, R. Labuťa, and V. Turek for useful discussions of Devonian stratigraphy and paleontology. A. Šulc prepared the conulariids, and V. Goliáš conducted the geochemical analyses. R. Labuťa and P. Lukeš donated several specimens. M. Mergl kindly provided the stratigraphical chart. This investigation was supported financially by the Ministry of Culture of the Czech Republic (DKRVO 2024-2028/2.III.a, National Museum, 00023272 to J.B.), and by the Czech Grant Foundation (GAČR; project No. 23-05217S). The work presented in this paper was partially supported by the Institutional Research Plan RVO67985891 of the Institute of Rock Structure and Mechanics of the Czech Academy of Sciences. We also thank the VLČS Quarry for long-term cooperation in rescue paleontological research in the area of Čertovy schody. Finally, we are grateful to the reviewers for their time spent on the manuscript and valuable suggestions.
REFERENCES
Adomat, F., Munnecke, A., and Kido, E. 2016. Mass occurrence of the large solitary rugose coral Phaulactis angusta at the boundary Lower/Upper Visby Formation in the Silurian of Gotland, Sweden: palaeoecology and depositional implications. GFF, 138:393–409.
https://doi.org./10.1080/11035897.2015.1103780
Amorin, K.B., Afonso, J.W.L., Leme, J.M.L., Diniz, C.Q.C., Rivera, L.C.M., Gómez-Gutiérrez, J.C., Boggiani, J.C., and Trindade, R.I.F. 2020. Sedimentary facies, fossil distribution and depositional setting of the late Ediacaran Tamengo Formation (Brazil). Sedimentology, 67(7):3422–3450.
https://doi.org/10.1111/sed.12749
Babcock, L.E., Feldmann, R.M., and Wilson, A.T. 1987. Teratology and pathology of some Paleozoic conulariids. Lethaia, 20:93–105.
https://doi.org/10.1111/j.1502-3931.1987.tb02025.x
Barrande, J. 1855. Remplissage organique du siphon dans certains céphalopodes paléozoïques. Bulletin de la Société Géologique de France, 2:441–489.
Barrande, J., 1867. Systême Silurien du centre de la Bohême. Ière Partie: Recherches Paléontologiques. Vol. III. Texte et 16 Planches. Classe des Mollusques. Ordre des Ptéropodes. Prague.
Bento, R., Feary, D.A., Hoey, A.S., and Burt., J.A. 2017. Settlement patterns of corals and other benthos on reefs with divergent environments and disturbances histories around the northeastern Arabian Peninsula. Frontiers in Marine Science, 4(305):1–12.
https://doi.org/10.3389/fmars.2017.00305
Berkowski, B. 2012. Life strategies and function of disseptiments in rugose coral Catactotoechus instabilis from the Lower Devonian of Morocco. Acta Palaeontologica Polonica, 57(2):391–400. https://doi.org/10.4202/app.2010.0048
Boller, M.L. and Carrington, E. 2006. The hydrodynamic effects of shape and size change during reconfiguration of a flexible macroalga. The Journal of Experimental Biology, 209:1894–1903.
https://doi.org/10.1242/jeb.02225
Bouček, B. 1928. Revise českých paleozoických konularií (Revision des conulaires Paléozoiques de la Bohême). Palaeontographica Bohemiae, 11:1–108.
Bouček, B. 1939. Conularida. Handbuch der Paläozoologie Band 2A, A113–A131.
Bouček, B. and Ulrich, F. 1939. O skořápce rodu Conularia Miller. Věstník Státního geologického ústavu Československé republiky, 5:1–25.
Bourillot, R., Vennin, E., Kolodka, Ch., Rouchy, J.-M., Caruso, A., Durlet, Ch., Chaix, Ch., and Rommevaux, V. 2009. The role of topography and erosion in the development and architecture of shallow-water coral bioherms (Tortonian-Messinian, Cabo de Gata, SE Spain). Palaeogeography, Palaeoclimatology, Palaeoecology, 281:92–114.
https://doi.org/10.1016/j.palaeo.2009.07.015
Bouška, J. 1946. On Crotalocrinitidae (Angelin) from the Silurian and Devonian of Bohemia. Rozpravy České Akademie Věd a Umění, Třída II, 56(4):1–17.
Brabcová, Z. 2001. Species diversification of conulariids from the Lower Palaeozoic of the Prague Basin (only in the Czech). Zprávy o geologických výzkumech v roce 2000:28–30.
Bruthansová, J., Bruthans, J., Van Iten, H., Rak, Š., and Schweigstillová J. 2022. Monospecific mass accumulations of Anaconularia anomala (Cnidaria, Scyphozoa) from the Upper Ordovician of the Czech Republic: sedimentological and palaeobiological significance. Lethaia, 55(2):1–18.
https://doi.org/10.18261/let.55.2.7
Chlupáč, I. 1998. Devonian, 148–198. In Chlupáč, I., Havlíček, V., Kříž, J., Kukal, Z., and Štorch, P. (eds.), Palaeozoic of the Barrandian (Cambrian to Devonian). Czech Geological Survey, Prague.
Chlupáč, I. 2003. Comments on facies development and stratigraphy of the Devonian, Barrandian area, Czech Republic. Bulletin of Geosciences, 78(4):299–312.
Dawson, J.W. 1868. Acadian geology: the geological structure, organic remains, and mineral resources of Nova Scotia, New Brunswick, and Prince Edward Island. Second Edition. Macmillan and Co., London, 694 p.
Denayer, J. and Aretz, M. 2012. Discovery of a Mississippian reef in Turkey: The Upper Viséan microbial-sponge-bryozoan-coral bioherm from Kongul Yayla (Taurides, S Turkey). Turkish Journal of Earth Sciences, 21:375–389.
https://doi.org/10.3906/yer-1008-3
Denny, M. and Gaylord, B. 2002. The mechanics of wave-swept algae. The Journal of Experimental Biology, 205:1355–1362.
https://doi.org/10.1242/jeb.205.10.1355
Di Martino, E. and Taylor, P.D. 2014. A brief review of seagrass-associated bryozoans, Recent and fossils. Studi Trentini di Scienze Naturali, 94:79–94.
Duval-Arnould, A., Bulot, L., Charton, R., Jain, S., Masrour, M., Pomar, L., Redfern, J., Simmons, M., and Schröder, S. 2024. Architecture of Oxfordian coral buildups along the Atlantic margin of Morocco. Journal of African Earth Sciences, 213, 105206:1–22.
https://doi.org/10.1016/j.jafrearsci.2024.105206
East, H.K., Johnson, J.A., Perry, Ch.T., Finlay, G., Musthag, A., Zahir, H., and Floyd, M. 2023. Seagrass meadows are important sources of reef island-building sediment. Communications Earth & Environment, 4(1):33:1–8.
https://doi.org/10.1038/s43247-023-00675-y
Falcucci, G., Amati, G., Fanelli, P., Krastev, V.K., Polverino, G., Porfiri, M., and Succi, S. 2021. Extreme flow simulations reveal skeletal adaptations of deep-sea sponges. Nature, 595:537–541.
https://doi.org/10.1038/s41586-021-03658-1
Ford, R.C., Van Iten, H., and Clark, G.R. 2016. Microstructure and composition of the periderm of conulariids. Journal of Paleontology, 90(3):389–399.
https://doi.org/10.1017/jpa.2016.63
Gallagher, C. and Doropoulos, C. 2017. Spatial refugia mediate juvenile coral survival during coral-predator interactions. Coral Reefs, 36:51–61.
https://doi.org/10.1007/s00338-016-1518-9
Hall, J. 1847. Palaeontology of New York: Containing descriptions of the organic remains of the lower division of the New-York system (equivalent to the Lower Silurian rocks of Europe). Volume I. C. Van Benthuysen, Albany.
Havlíček, V. 1981. Development of a linear sedimentary depression exemplified by the Prague basin (Ordovician-Middle Devonian; Barrandian area - central Bohemia). Sborník geologických věd, Geologie, 35:7–48.
Havlíček, V. 1998. Variscan folding and faulting in the Lower Palaeozoic basins, 165–169. In Chlupáč, I., Havlíček, V., Kříž, J., Kukal, Z., and Štorch, P. (eds.), Palaeozoic of the Barrandian (Cambrian to Devonian). Czech Geological Survey, Prague.
Hays, G.C. 2017. Ocean currents and marine life. Current Biology, 27(11): R431–R510.
https://doi.org/10.1016/j.cub.2017.01.044
Hladil, J. 1997. Lower Devonian open-sea elevation of Koněprusy: evolution of reef facies, pp. 31–36. In Čejchan, P. and Hladil, J. (eds.), Field Trip Book. Unesco-IGCP Project #335 "Biotic Recoveries from Mass Extinctions", Final Conference "Recoveries `97". Geological Institute AS CR, Eurocongress Centre, Prague.
Hladil, J. 1998. Facie koněpruských vápenců ve Velkolomu Čertovy schody (1994-1997). Český kras, 24:70–74.
Hladil, J. and Slavík, L. 1997. Facie a stratigrafie koněpruských vápenců (Koněprusy, Velkolom Čertovy schody, spodní devon, stupeň prag). Český kras, 23:5–18. (only in the Czech)
Infantes, E., Orfila, A., Bouma, T.J., Simarro, G., and Terrados, J. 2011. Posidonia oceanica and Cymodocea nodosa seedling tolerance to wave exposure. Limnology and Oceanography, 56(6):2223–2232.
https://doi.org/10.4319/lo.2011.56.6.2223
Janoušek, V., Hladil, J., Frýda, J., and Slavík, L. 2000. Strontium chemostratigraphy as an indicator of age and duration of reef sedimentation; a case study from Koneprusy reef of Pragian age (Devonian, central Bohemia). Journal of Conference Abstracts, 5:552, Goldschmidt 2000, Oxford.
Janoušek, V., Hladil, J., Frýda, J., Slavík, L., and Šmíd, J. 2001. Strontium chemostratigraphy of carbonate sediments - pilot study of Silurian and Devonian brachiopods from the Prague Basin. Geolines, 13:68–69.
Jebram, D. 1970. Preliminary experiments with Bryozoa in a simple apparatus for producing continuous water currents. Helgoländer Wissenschaftliche Meeresuntersuchungen, 20(1-4):278–292.
https://doi.org/10.1007/bf01609906
John, D.L., Hughes, N.G., Galaviz, M.I., Gunderson, G.O., and Meyer, R. 2010. Unusually preserved Metaconularia manni (Roy, 1935) from the Silurian of Iowa, and the systematics of the genus. Journal of Paleontology, 84(1):1–31.
https://doi.org/10.1666/09-025.1
Koehl, M.A.R. 1999. Ecological biomechanics of benthic oganisms: Life history, mechanical design and temporal patterns of mechanical stress. The Journal of Experimental Biology, 202:3469-3476.
https://doi.org/10.1242/jeb.202.23.3469
Koptíková, L., Bábek., O., Hladil, J., Kalvoda, J., and Slavík, L. 2010. Fine-grained non-carbonate particles embedded in neritic to pelagic limestones (Lochkovian to Emsian, Prague Synform, Czech Republic): composition, provenance and links to magnetic susceptibility and gamma-ray logs. Geologica Belgica, 13(4):407–430.
Kröger, B., Vinn, O., Toom, U., Corfe I.J., Kuva, J., and Zatoń, M. 2021. On the enigma of Palaenigma wrageli (Schmidt), a conulariid with a partly non-mineralized skeleton. PeerJ 9:e12374.
https://doi.org/10.7717/peerj.12374
Krs, M., Krsová, M., Pruner, P., and Havlíček, V. 1988. Palaeomagnetism, magnetism and palaeography of the Middle and Upper Cambrian rocks of the Barrandian area in the Bohemian Massif. Journal of Geological Sciences, Applied Geophysics, 22:9–48.
Krs, M., Krsová, M., and Pruner, P. 1997. Palaeomagnetism and palaeogeography of the Variscan and pre-Variscan formations of the Bohemian Massif, 162–173. In Vrána, S. and Štědrá, V. (eds.), Geological model of western Bohemia related to the KTB borehole in Germany. Journal of Geological Sciences, Geology, 47.
Leme, J.M., Simões, M.G., and Van Iten, H. 2010. Phylogenetic Systematics and Evolution of Conulariids. Lambert Academic Publishing, Berlin.
Leme, J.M., Van Iten, H., and Simões, M.G. 2022. A new conulariid (Cnidaria, Scyphozoa) from the terminal Ediacaran of Brazil. Frontiers in Earth Science, 10:777746.
https://doi.org/10.3389/feart.2022.777746
Lucas, S.G. 2012. The extinction of the conulariids. Geosciences, 2(1):1–10.
https://doi.org/10.3390/geosciences2010001
Luhar, M. and Nepf, H.M. 2011. Flow-induced reconfiguration of buoyant and flexible aquatic vegetation. Limnology and Oceanography, 56(6):2003–2017.
https://doi.org/10.4319/lo.2011.56.6.2003
Mapes, R.H., Fahrer, T.R., and Babcock, L.E. 1989. Sublethal and lethal injuries of Pennsylvanian conulariids from Oklahoma. Journal of Paleontology, 63:34–37.
https://doi.org/10.1017/s0022336000040920
Martínez-Quintana, Á., Lasker, H.R., and Wilson, A.M. 2023. Three-dimensional species distribution modelling reveals the realized spatial niche for coral recruitment on contemporary Caribbean reefs. Ecology Letters, 26:1497–1509.
https://doi.org/10.1111/ele.14281
Martone, P.T. and Denny, M.W. 2008. To bend a coralline: effect of joint morphology on flexibility and stress amplification in an articulated calcified seaweed. The Journal of Experimental Biology, 211:3421–3432.
https://doi.org/10.1242/jeb.020479
McNeil, M.A., Nothdurft, L.D., Dyriw, N.J., and Webster, J.M. 2021. Morphotype differentiation in the Great Barrier Reef Halimeda bioherm carbonate factory: International architecture and surface geomorphometrics. The Depositional Record, 7:176–199.
https://doi.org/10.1002/dep2.122
Mergl, M., Ferrová, L., and Frýda, J. 2016. Armoured test of Early Devonian Mesoconularia (Conulariida) from the Prague Basin (Cech Republic): probable adaptation to increased predation pressure. Bulletin of Geosciences, 91(3):561–581.
https://doi.org/10.3140/bull.geosci.1601
Mergl, M. 2022. Devonský koněpruský útes jako cíl geologické exkurze. Arnica, 12(1-2):16–42. (in Czech)
Mergl, M. and Kraft, P. 2023. Byronids and similar tubular fossils from the Devonian of the Barrandian area (Czech Republic). Earth and Environmental Science Transactions of the Royal Society of Edinburgh: 1–18.
https://doi.org/10.1017/S1755691023000099
Miller, A.A., Jacquet, S.M., Anderson, E.P., and Schiffbauer, J.D. 2022. Conulariids from the Silurian (late Telychian) Waukesha Lagerstätte, Wisconsin. Historical Biology, 34(12):2374–2394.
https://doi.org/10.1080/08912963.2021.2017917
Neuman, B.E.E. 1988. Some aspects of life strategies of Early Paleozoic rugose corals. Lethaia, 21:97–114.
https://doi.org/10.1111/j.1502-3931.1988.tb02061.x
Patočka, F., Pruner, P., and Štorch, P. 2003, Palaeomagnetism and geochemistry of Early Palaeozoic rocks of the Barrandian (Teplá-Barrandian Unit, Bohemian Massif): palaeotectonic implications. Physics and Chemistry of the Earth, 28:735–749.
https://doi.org/10.1016/s1474-7065(03)00126-8
Prokop, R.J. and Petr, V. 1991. Torrocrinus grandis, gen. et sp. n. (Crinoidea) from Lower Devonian Koněprusy Limestone of the Barrandian area (Czechoslovakia). In Roček, Z. (ed.), Czechoslovak Paleontology 1990 (Abstract): 21. Carolinum Press, Praha
Robson, S.P. and Young, G.A. 2013. Late Ordovician conulariids from Manitoba, Canada. Journal of Paleontology, 87(5):775–785.
https://doi.org/10.1666/12-0370
Rodrigues, A.C., Simões, M.G., and Leme, J.M. 2003. Tafonomia Comparada dos Conulatae (Cnidaria), Formação Ponta Grossa (Devoniano), Bacia do Paraná, Estado do Paraná. Revista Brasileira de Geociências, 33(4):381–390.
Rummel, A. 2014. Flexion of hydroid colonies in a current. Unpublished Thesis, Department of Biology, University of Chicago, Chicago, USA:1–11.
Scrutton, T.C. 1998. The Palaeozoic corals, II: structure, variation and palaeoecology. Proceedings of the Yorkshire Geological Society, 52(1):1–57.
https://doi.org/10.1144/pygs.52.1.1
Simões, M.G, Mello, L.H.C., Rodrigues, S.C., Leme, J.M., and Marques, A.C. 2000. Conulariid taphonomy as a tool in paleoenvironmental analysis. Revista Brasileira de Geociencias, 30(4):757–762.
Simões, M.G, Rodrigues, S.C., Leme, J.M., and Van Iten, H. 2003. Some middle Paleozoic conulariids (Cnidaria) as possible examples of taphonomic artifacts. Journal of Taphonomy, 1(3):165–181.
Slavík, L. 2004. A new conodont zonation of the Pragian Stage (Lower Devonian) in the stratotype area (Barrandian, central Bohemia). Newsletters on Stratigraphy, 40(1-2):39–71.
https://doi.org/10.1127/0078-0421/2004/0040-0039
Slavík, L. and Hladil, J. 2007. Early Pragian conodont-based correlations between the Barrandian area and the Spanish Central Pyrenees. Geological Journal, 42, 499-512.
https://doi.org/10.1002/gj.1087
Slavík, L. and Hladil, J. 2020. Early Devonian (Lochkovian - Early Emsian) bioevents and conodont response in the Prague Synform (Czech Republic). Palaeogeography, Palaeoclimatology, Palaeoecology, 549(2020):109148.
https://doi.org/10.1016/j.palaeo.2019.04.004
Sorauf, J.E. 2001. External morphology and paleobiology of Heliophyllum halli (Zoantharia, Rugosa), from the Middle Devonian Hamilton Group of New York State. Journal of Paleontology, 75(1):24–33.
https://doi.org/10.1017/s0022336000031887
Van Iten, H. 1991. Anatomy, patterns of occurrence, and nature of the conulariid schott. Palaeontology, 34:939–954.
Van Iten, H. 1992. Microstructure and growth of the conulariid test: implications for conulariid affinities. Palaeontology, 35:359–372.
Van Iten, H. and Vyhlasová, Z. 2004. Conulariids, 119-123. In Webby, B.D., Droser, M.L., and Paris, F. (eds.), The Great Ordovician Biodiversification Event. Columbia University Press, New York.
https://doi.org/10.7312/webb12678-015
Van Iten, H., Simões, M.G., Marques, A.C., and Collins, A.G. 2006a. Reassessment of the phylogenetic position of conulariids (?Ediacaran-Triassic) within the subphylum Medusozoa (phylum Cnidaria). Journal of Systematic Palaeontology, 4:109–118.
https://doi.org/10.1017/s1477201905001793
Van Iten, H., Leme, J.M., Rodrigues, S.C., and Simões, M.G. 2006b. New data on the anatomy of Conularia milwaukeensis Cleland, 1911 (Middle Devonian, Iowa and Wisconsin). Journal of Paleontology, 80:393–395.
https://doi.org/10.1666/0022-3360(2006)080[0392:ndotao]2.0.co;2
Van Iten, H., Leme, J.M., and Simões, M.G. 2006c. Additional observations on the gross morphology and microstructure of Baccaconularia Hughes, Gunderson et Weedon, 2000, a Cambrian (Furongian) conulariid from the north-central USA. Palaeoworld, 15:294–306.
https://doi.org/10.1016/j.palwor.2006.10.006
Van Iten, H., Tollerton, V.P., Ver Straeten, C.F., Leme, J.M., Simões, M.G., and Rodrigues, S.C. 2013. Life mode of in situ Conularia in a Middle Devonian epibole. Palaeontology, 56(1): 29–48.
https://doi.org/10.1111/j.1475-4983.2012.01146.x
Van Iten, H., Marques, A.C., Leme, J.M., Forancelli Pacheco, M.L.A., and Simões, M.G. 2014a. Origin and early evolution of the phylum Cnidaria Verrill: major developments in the analysis of the taxon´s Proterozoic-Cambrian history. Palaeontology, 54:677–690.
https://doi.org/10.1111/pala.12116
Van Iten, H., Burkey, M., Leme, J.M., and Marques, A.C. 2014b. Cladistics and mass extinctions: the example of conulariids (Scyphozoa, Cnidaria) and the End Ordovician Extinction Event. GFF, 136:275–280.
https://doi.org/10.1080/11035897.2014.880506
Van Iten, H., Muir, L., Simões, M.G., Leme, J.M., Marques, A.C., and Yoder, N. 2016. Palaeobiogeography, palaeoecology and evolution of Lower Ordovician conulariids and Sphenothallus (Medusozoa, Cnidaria), with emphasis on the Fezouata Shale of southeastern Morocco. Paleogeography, Palaeoclimatology, Palaeoecology, 460:170–178.
https://doi.org/10.1016/j.palaeo.2016.03.008
Van Iten, H., Gutiérrez-Marco, J.C., Muir, L.A., Simões, M.G., and Leme, J.M. 2020. Ordovician conulariids (Scyphozoa), from the Upper Tiouririne Formation (Katian), eastern Anti-Atlas Mountains, southern Morocco. Geological Society Special Publication, 485:1–23.
https://doi.org/10.1144/SP485.5
Van Iten, H., Mironenko, A., and Vinn, O. 2022. A new conulariid from the Upper Mississippian (early Serpukhovian) of Central Russia (Moscow Basin): systematics, microstructure, and growth abnormalities. PalZ, 97(2):311-322.
https://doi.org/10.1007/s12542-022-00636-4
Van Iten, H., Snyder, M., and Tollerton, V.M., Jr. 2024. Type material of Paraconularia planicostata (Dawson) from the Upper Mississippian of Nova Scotia, Atlantic Canada. Journal of Paleontology, 98(1): 152–155.
https://doi.org/10.1017/jpa.2023.86
Van Tussenbroek, B.I. 2011. Dynamics of seagrasses and associated algae in coral reef lagoons. Hidrobiolégica, 21(3):293–310.
Vinn, O., Ernst, A., Wilson, M.A., and Toom, U. 2019. Symbiosis of conulariids with trepostome bryozoans. Palaeogeography, Palaeoclimatology, Palaeoecology, 518:89–96.
https://doi.org/10.1016/j.palaeo.2019.01.018
Vinn, O. 2022. Phosphatic biomineralization in Scyphozoa (Cnidaria): A review. Minerals, 12(10):131–6.
https://doi.org/10.3390/min12101316
Wang, Y., Jin, J., Zhan, R., and Copper, P. 2014. Early Silurian ‘algal meadows’ of Anticosti Island, eastern Canada: an analogue to modern sea grass meadows? Geology Today, 30(2): 67–70.
https://doi.org/10.1111/gto.12043
Weinerová, H., Hron, K., Bábek, O., Šimíček, D., and Hladil, J. 2017. Quantitative allochem compositional analysis of Lochkovian-Pragian boundary sections in the Prague Basin (Czech Republic): results from multivariate statistic. Sedimentary Geology, 354:43–59.
https://doi.org/10.1016/j.sedgeo.2017.04.002
Zicha, O., Bruthansová, J., and Kraft, P. 2020. Epibionts on shells in the Šárka Formation: a sparsely occupied niche in the lower to middle Darriwilian (Oretanian, Ordovician) in the Prague Basin (Czech Republic). Palaeogeography, Palaeoclimatology, Palaeoecology, 550: 109401.
https://doi.org/10.1016/j.palaeo.2019.109401
