

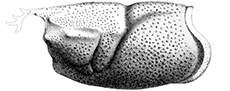
 Taxonomic revision of the extinct clawed lobster genus Oncopareia Bosquet, 1854 (Decapoda, Astacidea, Nephropidae)
Taxonomic revision of the extinct clawed lobster genus Oncopareia Bosquet, 1854 (Decapoda, Astacidea, Nephropidae)
Article number: 25.2.a20
https://doi.org/10.26879/1190
Copyright Paleontological Society, June 2022
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 1 October 2021. Acceptance: 24 May 2022.
ABSTRACT
Oncopareia Bosquet, 1854, is an extinct clawed lobster genus within the clade of thaumastocheliform decapod crustaceans that are characterized by short, quadrate pleura on the pleon and a major claw with a bulbous palm and acicular dentition. In fact, Oncopareia is the earliest thaumastocheliform lobster known to date, the oldest example being of Turonian age (~90 Ma). Originally, Oncopareia was erected for a single late Maastrichtian species, O. bredai Bosquet, 1854, from the southeast Netherlands. The genus has a convoluted taxonomic history that begins with a type species that is an accidental composite of two lobsters that differ at the genus level, the second genus being Hoploparia M’Coy, 1849. An analogous situation occurred in Hoploparia biserialis Fritsch, in Fritsch and Kafka, 1887, the description of which is partly based on material that is attributable to Oncopareia. A number of subsequently erected taxa have been assigned to Oncopareia, based solely on isolated cheliped fingers and only later to be recognized as representatives of the ghost shrimp genus Ctenocheles Kishinouye, 1926. This was due to convergent evolution of pectinate claws in several distinct decapod lineages. The present contribution summarizes the taxonomic history of Oncopareia, redefines the genus and redescribes its type species, and comments on species previously referred to the genus. Herein, in addition to the type species, three distinct Oncopareia species are recognized: Oncopareia esocina (Fritsch, in Fritsch and Kafka, 1887), Oncopareia klintebjergensis Jakobsen and Collins, 1979, and Oncopareia lunata (Fritsch, in Fritsch and Kafka, 1887). Other species previously included within Oncopareia are either reassigned to other genera or considered of uncertain affinity (incertae sedis).
Dale M. Tshudy. Department of Geosciences, Edinboro University of Pennsylvania, Edinboro, Pennsylvania 16412, USA dtshudy@edinboro.edu https://orcid.org/0000-0001-5382-4378
Matúš Hyžný. Department of Geology and Paleontology, Faculty of Natural Sciences, Comenius University, Ilkovičova 6, Bratislava, Slovakia. hyzny.matus@gmail.com https://orcid.org/0000-0002-8960-2846)
Martina Kočová Veselská. Institute of Geology of the Czech Academy of Sciences, v. v. i., Department of Paleobiology and Paleoecology, Rozvojová 269, 165 00 Prague 6, Czech Republic and Institute of Geology and Palaeontology, Charles University, Albertov 6, 128 43 Prague 2, Czech Republic. veselskamartina@gmail.com https://orcid.org/0000-0003-4322-9019
John W.M. Jagt. Natuurhistorisch Museum Maastricht, De Bosquetplein 6-7, 6211 KJ Maastricht, the Netherlands. john.jagt@maastricht.nl https://orcid.org/0000-0001-6216-1991
Key words: acicular dentition; convergent evolution; pectinate claws; thaumastocheliform lobsters
Final citation: Tshudy, Dale M., Hyžný, Matúš, Kočová Veselská, Martina, and Jagt, John W.M. 2022. Taxonomic revision of the extinct clawed lobster genus Oncopareia Bosquet, 1854 (Decapoda, Astacidea, Nephropidae). Palaeontologia Electronica, 25(2):a20. https://doi.org/10.26879/1190
palaeo-electronica.org/content/2022/3628-lobster-genus-oncopareia
Copyright: June 2022 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
Since Mesozoic times, clawed lobsters have been important components of marine ecosystems (Schweitzer and Feldmann, 2014). They comprise an array of not necessarily related taxa, representatives of which are assigned to a number of distinct families. The most diverse of these is the Nephropidae Dana, 1852, consisting of extinct and extant genera with a stratigraphical range from the Valanginian (Lower Cretaceous) onwards (Tshudy et al., 2018). The taxonomy of fossil nephropid lobsters is convoluted, partly because of usage of rather loosely defined genera such as Hoploparia (see Tshudy and Sorhannus, 2003; Tshudy et al., 2018) and Oncopareia. Untangling the taxonomy of the latter genus is the major goal of the present contribution.
Oncopareia was erected for a single late Maastrichtian species, O. bredai Bosquet, 1854, from the Kunrade limestone (now Kunrade Formation) in the Kunrade area of southern Limburg, the Netherlands. Unfortunately, the description was based on several specimens belonging to two different lobster taxa, as discussed further below. Since then, a number of extinct lobster taxa have been assigned to Oncopareia. The taxonomic history of Oncopareia bredai has been discussed on several occasions, although never fully resolved. Feldmann et al. (1990) described four specimens (USNM collections, leg. J.W.M. Jagt) from the Vijlen Member (interval 6; Gulpen Formation; late early Maastrichtian) of the Haccourt-Lixhe area in northeast Belgium and referred them to O. bredai. They noted that Bosquet (1854) had not selected a type specimen for his O. bredai and advised (Feldmann et al., 1990, p. 29) that a lectotype be designated in a future publication. Tshudy (1993, pp. 275-307), in his unpublished PhD thesis, studied the Bosquet and Ubaghs collections and attempted to untangle the taxonomic confusion surrounding Oncopareia. He redefined that genus and its type species, O. bredai, and commented on other species that were held to be referable to the redefined genus. In discussing convergent evolution in decapod crustacean claws, Tshudy and Sorhannus (2000a, figs. 3-5 = Tshudy, 1993, figs. 38-40) published three plates illustrating Oncopareia; some specimens (Tshudy and Sorhannus, 2000a, fig. 5) were later assigned to Thaumastocheles Wood-Mason, 1874, by Tshudy and Saward (2012, pp. 10-11). Still, no detailed discussion of the taxonomic history of Oncopareia has been published to date, nor has a lectotype been designated in the published literature. Nonetheless, because Thaumastocheles-like specimens do exist in the type collection, we herein redefine O. bredai on the basis of those specimens.
Oncopareia, with its pectinate claws, belongs to thaumastocheliform lobsters (sensu Chang et al., 2017), which are morphologically very similar to each other, and collectively different from other “lobsters” (= morphotype, sensu Schweitzer and Feldmann, 2014). As a group, their record tells an interesting story, of evolution during the Late Cretaceous, of dwelling at shelf depths during that time interval, of a loss of shelf-dwelling habit in the Paleogene (Tshudy and Saward, 2012) and of a reduction or even complete loss of eyes in extant, deep-water representatives (Tshudy and Saward, 2012; Chang et al., 2017). The evolution of this group (as a clade, see Karasawa et al., 2013 and Bracken-Grissom et al., 2014) has recently been assessed by cladistic analysis of morphological, molecular, and combined data (Chang et al., 2017).
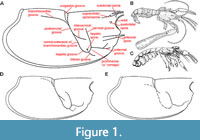 Thaumastocheliform lobsters, however, are not the only group of decapod crustaceans possessing pectinate claws (Figure 1). These have been developed independently in several lineages (Tshudy and Sorhannus, 2000a), including polychelid lobsters (Polychelida: Palaeopentachelidae), Astacidea (Erymidae, Nephropidae, Stenochiridae), Axiidea (Ctenochelidae), and Brachyura (Iphiculidae, Leucosiidae), causing difficulties in taxonomic evaluation of fossil occurrences solely based on isolated cheliped elements. In fact, a number of extinct taxa have been assigned to Oncopareia based merely on isolated cheliped fingers only to be later recognized as representatives of Ctenocheles (Tshudy, 1993). Thus, not only is the generic concept of Oncopareia complex, but also the differentiation of Oncopareia from other forms can be difficult.
Thaumastocheliform lobsters, however, are not the only group of decapod crustaceans possessing pectinate claws (Figure 1). These have been developed independently in several lineages (Tshudy and Sorhannus, 2000a), including polychelid lobsters (Polychelida: Palaeopentachelidae), Astacidea (Erymidae, Nephropidae, Stenochiridae), Axiidea (Ctenochelidae), and Brachyura (Iphiculidae, Leucosiidae), causing difficulties in taxonomic evaluation of fossil occurrences solely based on isolated cheliped elements. In fact, a number of extinct taxa have been assigned to Oncopareia based merely on isolated cheliped fingers only to be later recognized as representatives of Ctenocheles (Tshudy, 1993). Thus, not only is the generic concept of Oncopareia complex, but also the differentiation of Oncopareia from other forms can be difficult.
The aims of the present contribution, following up on Tshudy (1993) are threefold: to summarize the taxonomic history of Oncopareia, to redefine the genus and redescribe its type species, with designation of a lectotype and, lastly, to present a taxonomic treatment of species previously referred to the genus.
MATERIAL AND METHODS
The composite nature of the type series of Oncopareia bredai was revealed to one of us (DMT) in 1990 when examining specimens contained in the IRScNB collections (Brussels). The Bosquet Collection was found, after much searching with the late Annie V. Dhondt, and one of the co-authors (JWMJ), in a wooden box under a thick layer of dust in the basement. It seemed possible that the collection had not been examined since it had been transferred to Brussels after Bosquet’s death in 1880. Bosquet (1854) discussed specimens contained in three collections, including his own and those of Dr. Van Breda (now in the Department of Palaeontology, The Natural History Museum, London, UK) and the ‘Musée Géologique de Harlem’ (= Teylers Museum, Haarlem, the Netherlands).
Specimens of Oncopareia bredai available for the present study are housed in the IRScNB, MNHN, NHMM, NM, and USNM collections. Approximately 33 specimens are contained in the IRScNB collections; most (25) of these are in the Ubaghs Collection (IG 6521); five in the Bosquet Collection (IG 4285), and three in the Rutot Collection (IG 5425). Bosquet (1854) made frequent reference to specimens in the collections of Dr. Van Breda and of the ‘Musée Géologique de Harlem’; we have not re-examined these, if still extant. Other taxa discussed in detail further were studied either firsthand using the type material or via literature surveys (when the material has been lost or was not accessible to us). Details are indicated in the systematic section.
Specimens studied were documented photographically by all co-authors; in some cases, ammonium chloride was used to enhance the contrast prior to photography. Details of geographical and stratigraphical provenance and current repositories are given in respective figure captions.
Abbreviations. To denote the repositories of the material illustrated and/or referred to in the text, the following abbreviations are used: ANSP, Academy of Natural Sciences, Philadelphia, USA; IRScNB, Institut Royal des Sciences Naturelles de Belgique, Brussels, Belgium; MGUH, Statens Naturhistorisk Museum (formerly Geological Museum), Copenhagen, Denmark; MNHN, Muséum National d'Histoire Naturelle, Paris, France; NM, National Museum, Prague, Czech Republic; NHMM, Natuurhistorisch Museum Maastricht, Maastricht, the Netherlands; NHMUK, The Natural History Museum, London, UK; TM, Teylers Museum, Haarlem, the Netherlands; USNM, United States National Museum, Washington, DC, USA.
FOSSIL THAUMASTOCHELES-LIKE LOBSTERS - A HISTORICAL OVERVIEW
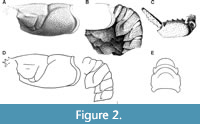 Oncopareia was erected for a single late Maastrichtian species, O. bredai Bosquet, 1854, from what is now the Kunrade Formation in the Kunrade area of southern Limburg, the Netherlands. The taxonomic history of the genus is complex, beginning with a type species that is an accidental composite of two lobsters (Figure 2A-2D) that differ at the genus level: Bosquet (1854) unknowingly established O. bredai on the basis of the cephalothorax of one species (Figure 2A) and the pleon (Figure 2B) and claws (Figure 2C) of another. One of the two lobsters comprising the unwitting composite type series of O. bredai (sensu Bosquet) is entirely Hoploparia-like. The other lobster has a very similar, Hoploparia-like cephalothorax but Thaumastocheles-like pleon and claws (as discussed further below in the systematic section). These cephalothoraxes belong with the Thaumastocheles-like pleons and chelipeds in the Bosquet Collection. Bosquet seems to have attributed the Thaumastocheles-like pleon forms to deformation (Bosquet, 1854, footnote on p. 129 [119]). Pelseneer (1886, p. 166), who examined the Bosquet and Ubaghs collections, was the first to recognize the Thaumastocheles-like pleon form as real (i.e., undeformed) and meaningful (he said it was Thalassina-like, noting especially the ridge separating the terga and pleura. He illustrated this pleon type in a line drawing (Pelseneer, 1886, fig. 4; Figure 2E here) and referred to it as a new species, Homarus bosqueti.
Oncopareia was erected for a single late Maastrichtian species, O. bredai Bosquet, 1854, from what is now the Kunrade Formation in the Kunrade area of southern Limburg, the Netherlands. The taxonomic history of the genus is complex, beginning with a type species that is an accidental composite of two lobsters (Figure 2A-2D) that differ at the genus level: Bosquet (1854) unknowingly established O. bredai on the basis of the cephalothorax of one species (Figure 2A) and the pleon (Figure 2B) and claws (Figure 2C) of another. One of the two lobsters comprising the unwitting composite type series of O. bredai (sensu Bosquet) is entirely Hoploparia-like. The other lobster has a very similar, Hoploparia-like cephalothorax but Thaumastocheles-like pleon and claws (as discussed further below in the systematic section). These cephalothoraxes belong with the Thaumastocheles-like pleons and chelipeds in the Bosquet Collection. Bosquet seems to have attributed the Thaumastocheles-like pleon forms to deformation (Bosquet, 1854, footnote on p. 129 [119]). Pelseneer (1886, p. 166), who examined the Bosquet and Ubaghs collections, was the first to recognize the Thaumastocheles-like pleon form as real (i.e., undeformed) and meaningful (he said it was Thalassina-like, noting especially the ridge separating the terga and pleura. He illustrated this pleon type in a line drawing (Pelseneer, 1886, fig. 4; Figure 2E here) and referred to it as a new species, Homarus bosqueti.
Subsequent to Bosquet’s erection of a composite type species, the taxonomic confusion surrounding fossil Thaumastocheles-like lobsters became even more serious. Workers in the late 1800s erected new genera at a time when access to information and communication among taxonomists was not what it is today. This led to synonymous genera, even for specimens that were anatomically fairly complete. Other scholars in the late 1800s and early 1900s described isolated chelipeds with bulbous palms and slender fingers armed with acicular denticles; most of these were misidentified at the infraordinal level until recently (Tshudy, 1993).
Schlüter (1862) was the first to describe extinct lobsters with a morphology similar to that of extant Thaumastocheles and erected (p. 728) the genus Nymphaeops. The type species, N. coesfeldiensis (considered herein to be a junior synonym of O. bredai; see below), was based on specimens which, although lacking well-preserved claws, were otherwise essentially complete. Schlüter (1862) rightly considered his new species to be distinct from Hoploparia mainly because it had a different (i.e., Thaumastocheles-like) pleon, with wide, trapezoidal terga and short pleura but also, in his opinion, a different pattern of carapace grooves. In the same paper, he named another new species, Nymphaeops sendenhorstensis (considered herein as indeterminate at the generic level), based on a specimen with cephalothorax, partial pleon, and large, isochelous claws.
From the Bosquet Collection, a single claw with a bulbous palm and acicular dentition was referred by Pelseneer (1886, p. 162 [2], fig. 1) to a new genus and species, Ischnodactylus inaequidens. Why he described this particular single claw, but failed to mention other claws with acicular denticles, is uncertain. Ironically, the claw he described was one of probably only two specimens at his disposal, which is not an astacidean lobster at all, but an axiidean ghost shrimp referable to the genus Ctenocheles. In the same paper, Pelseneer (1886) moved Hoploparia macrodactyla Schlüter, in von der Marck and Schlüter, 1868, to Ischnodactylus, based on the similarity of its claws to that of the type species.
With two extinct, Thaumastocheles-like genera now published (i.e., Nymphaeops and Ischnodactylus), Fritsch and Kafka (1887) erected the third one, Stenocheles, on the basis of three specimens from three different localities. Fritsch and Kafka (1887) described two species, Stenocheles esocinus and S. parvulus, for this material. Although resembling in many aspects the concept of Ischnodactylus published a year before, they made no comparison with it indicating that they were unaware of the latter when they prepared their own manuscript. Later, Hyžný et al. (2014) revised the original material referred to Stenocheles by Fritsch and Kafka (1887) and considered all three specimens distinct enough to be attributed to three different species, and erected a new one, Ctenocheles fritschi Hyžný, Kočová Veselská, and Dvořák, 2014, to accommodate one of the syntypes of S. parvulus. Fritsch and Kafka (1887) did not select a type species for Stenocheles; this was done by Glaessner (1969), who designated Stenocheles esocinus as such. This species is based on a pleon and associated claw with long, slender fingers and acicular dentition.
Subsequent to Fritsch and Kafka’s (1887) work came the discovery of more claws with bulbous palms and long, slender fingers and acicular dentition. These claws were referred mostly to Ischnodactylus by various authors. Böhm (1891) erected I. pectiniformis for a single claw, while Lőrenthey and Beurlen (1929) introduced Hoploparia eocenica (listed as I. (?) eocaenicus by Mertin, 1941) for partly preserved internal molds of cephalothoraxes and separate claws. Rathbun (1935) referred four new species, I. cookei, I. cultellus, I. texanus, and I.? dentatus to this genus; all were based on claw fragments. Finally, Beurlen (1939) assigned claws from the Oligocene of Hungary to Thaumastocheles. Most of these chelipeds were misidentified, as will be discussed in detail below.
Mertin (1941, p. 178) expanded the genus Oncopareia from one species, i.e., the type, O. bredai, to several on the basis of his examination of new, anatomically fairly complete material from the Upper Cretaceous of Braunschweig, northwest Germany. That material revealed the association of fossil remains of Thaumastocheles-like claws and pleons for the first time. Much earlier, Pelseneer (1886) had noticed the thaumastocheliform pleons (i.e., with short quadrate pleura) in the Bosquet Collection but he had not recognized the association with any pectinate chelipeds. With that recognition, Mertin (1941) attempted to resolve a situation wherein extinct Thaumastocheles-like lobsters had been described under different names. He synonymized Stenocheles Fritsch, in Fritsch and Kafka, 1887, and Ischnodactylus Pelseneer, 1886, with Nymphaeops Schlüter, 1862, and these, in turn, with Oncopareia.
The similarity of a Braunschweig pleon with that of Stenocheles esocinus (both of the Thaumastocheles type), in addition to some similarities between the Thaumastocheles- type claws of the two genera, led Mertin (1941) to relegate Stenocheles into the synonymy of Ischnodactylus. The similarity of the cephalothorax and pleon of the Braunschweig lobster to that of the type species, Nymphaeops coesfeldiensis, led Mertin (1941) to declare Stenocheles (= Ischnodactylus) a synonym of Nymphaeops.
Mertin’s (1941) action, however, was too extensive, because not all of the species synonymized possess the Thaumastocheles-like pleon and pectinate claw upon which the grouping was based. Far more perplexing, however, is that he synonymized the above forms with Oncopareia, a genus which, as described by Bosquet (1854), has neither the Thaumastocheles-like pleon, nor claw which unite the three other fossil genera. In fact, the entire anatomy of Oncopareia in the original description of Bosquet (1854; but not Oncopareia as redefined herein) is highly typical of Hoploparia. Mertin’s synonymization seems to have been based entirely on the shared possession of a spiny rostrum in his Braunschweig material and in Oncopareia (sensu Bosquet, 1854).
Mertin (1941) transferred to Oncopareia all three fossil genera, Nymphaeops, Ischnodactylus, and Stenocheles, as well as the fossil species Hoploparia macrodactylus and Enoploclytia paucispina Schlüter, in von der Marck and Schlüter, 1868. Into the same group, he tentatively moved Stenocheles parvulus, Hoploparia biserialis, Homarus muncki Pelseneer, 1885, and, with some reservation, the extant genus Thaumastocheles. He excluded O. heterodon Bosquet, 1854, and mentioned that Nymphaeops belgicus Forir, 1887, and Ischnodactylus pectiniformis were based on material that was insufficient to permit comparison.
Mertin (1941, p. 178) redefined Oncopareia to include Thaumastocheles-like characteristics; most notably, the pleon and claw with acicular dentition. He did this not because the type species, O. bredai, had these features, but because some different, Thaumastocheles-like species possessed them, and some of these, in turn, had a spiny rostrum - a generic character for Oncopareia Bosquet, 1854. Thus, on the basis of the shared possession of a spiny rostrum, the Thaumastocheles-like extinct species were allied with O. bredai, and the latter - being first-named - became the type species. Apparently, Mertin (1941) disregarded the Hoploparia/Homarus-like pleon and the absence of Thaumastocheles-like claws in Bosquet’s original publication. Nonetheless, he did establish Oncopareia as an extinct ancestor of Thaumastocheles - a concept that gained traction in the literature.
Mertin (1941, p. 180) noted the strong similarity of Oncopareia to the extant Thaumastocheles (i.e., carapace groove pattern, a very spiny rostrum, a smooth carapace, a lateral ridge on the pleon, pleonal pleura, etc.). He opined (Mertin, 1941, pp. 178, 180) that it was doubtful that the fossil and extant forms could be separated on the genus level.
Oncopareia was treated in detail by Tshudy (1993), who examined the type material of Oncopareia bredai. Although never published, results of his research found their way into several works dealing with thaumastocheliform lobsters (Tshudy and Sorhannus, 2000a, 2000b; Tshudy and Saward, 2012; Chang et al., 2017). Differences in the cephalothorax groove pattern in Oncopareia bredai and Hoploparia beyrichi were graphically presented by Tshudy and Sorhannus (2000b, fig. 2) and are redrawn here in Figures 1D and 1E. The present contribution, finally, aims to resolve the taxonomic status of Oncopareia bredai as the type species of the genus Oncopareia and discusses all taxa treated as Oncopareia at any time in the light of the present revision.
SYSTEMATIC PALEONTOLOGY
The systematic section presents all taxa classified as Oncopareia at any time. The section starts with species recognized as representatives of Oncopareia as revised herein, followed by taxa referable to other genera. All taxa are arranged within the higher-level classification proposed by De Grave et al. (2009); dubious ones are included as well.
DECAPODA Latreille, 1802
ASTACIDEA Latreille, 1802
NEPHROPIDAE Dana, 1852
Genus Oncopareia Bosquet, 1854
= Nymphaeops Schlüter, 1862 [partim]
= Ischnodactylus Pelseneer, 1886 [partim]
= Stenocheles Fritsch, 1887 [partim]
Type species. Oncopareia bredai Bosquet, 1854, by monotypy.
Diagnosis. Cephalothorax slightly laterally compressed, with well-developed postcervical, cervical, intercervical, branchiocardiac, and antennal grooves; combined postcervical-branchiocardiac groove parallel with cervical groove; pleon with short, quadrate pleura, and pectinate claw with bulbous palm and acicular dentition.
Emended description. Rostrum long, dorso-laterally compressed, spinose. Postcervical groove extending anteroventrally from dorsal midline at about mid-length. Combined branchiocardiac-postcervical groove extending toward hepatic groove, bifurcating at above mid-height on carapace. Anterior extension of postcervical groove extending toward middle of cervical groove. Cervical and postcervical grooves parallel. Ventral extension of branchiocardiac groove extending toward, or to, hepatic groove. Branchiocardiac groove long dorsally; extending from near posterodorsal margin to postcervical groove. Postorbital and supraorbital spine present. Pleonal terga wide, transversely convex; lateral carina at tergum-pleuron boundary. Pleura quadrate, wider than long. Pereiopod 1 chelipeds unequal in size and form. Pectinate claw with short, bulbous palm and very long, slender fingers bearing acicular denticles of varying length. Crusher-type claw robustly constructed.
Remarks. Oncopareia, as redefined herein, strongly resembles other thaumastocheliform lobsters (nephropids with pectinate claws), such as Thaumastocheles, Thaumastochelopsis Bruce, 1988 and Dinochelus Ahyong, Chan and Bouchet, 2010. All of these genera, being representatives of the same clade (Karasawa et al., 2013; Bracken-Grissom et al., 2014), are characterized as having short, quadrate pleura and a major claw with a bulbous palm and acicular dentition. Oncopareia differs from these in having: 1) a carapace that is laterally compressed (not dorsoventrally compressed); 2) parallel cervical and postcervical grooves; 3) a major claw palm that is not bilobed; and in lacking: 1) an oblique carina near the posterior corner of the pleura; 2) pleural marginal spines (Tshudy and Saward, 2012, table 1; including additional comparisons). Our present redefinition of Oncopareia is based on careful examination of the type material of the type species, i.e., O. bredai.
 Bosquet’s illustrations (1854, pl. 10, figs. 5‒8) show the cephalothorax of the lobster (resembling that of both Hoploparia and Thaumastocheles; his figs. 5, 61 and 8; Figure 3A-3C here) and the pleon and claws of another (Hoploparia-like; his figs. 6, 7, 71 a-b). Bosquet (1854) established Oncopareia bredai on the basis of the cephalothorax of one species (which is similar to that of both the extant Thaumastocheles and fossil Hoploparia [e.g., similar overall shape, groove pattern and spines]) and the pleon and claws of another (Hoploparia-like [e.g., a pleon with cordate, pointed pleura; claws of normal nephropid shape]). Bosquet never realized this and failed to recognize the Thaumastocheles-like pleons as real (undeformed) and significant.
Bosquet’s illustrations (1854, pl. 10, figs. 5‒8) show the cephalothorax of the lobster (resembling that of both Hoploparia and Thaumastocheles; his figs. 5, 61 and 8; Figure 3A-3C here) and the pleon and claws of another (Hoploparia-like; his figs. 6, 7, 71 a-b). Bosquet (1854) established Oncopareia bredai on the basis of the cephalothorax of one species (which is similar to that of both the extant Thaumastocheles and fossil Hoploparia [e.g., similar overall shape, groove pattern and spines]) and the pleon and claws of another (Hoploparia-like [e.g., a pleon with cordate, pointed pleura; claws of normal nephropid shape]). Bosquet never realized this and failed to recognize the Thaumastocheles-like pleons as real (undeformed) and significant.
Bosquet (1854, p. 128) referred to his pl. 10, fig. 5 (Figure 3A here) as depicting the most perfect cephalothorax of Oncopareia he had examined. The specimen is identified in the plate caption as belonging to his own collection, but it is unknown which specimen of the IRScNB lot this is. However, it does bear a resemblance to IRScNB 90-32c (Ubaghs Collection) (see Tshudy, 1993, fig. 38.1; Tshudy and Sorhannus,2000a, fig. 3.1; Figure 3D here), although, if it is, it is imperfectly drawn because in this particular specimen the postorbital carina angles upward, instead of downward as illustrated, and the granules in Bosquet’s plate 10, figure 5, added by inference from other specimens [as explained in the plate caption] are drawn too coarsely.
The rostrum in Bosquet’s plate 10, figure 8 (Figure 3B here), described (footnote on p. 128) as an “impreinte” and added (dashed line) by him to his plate 10, figure 5, might be based on IRScNB 90-15b (Bosquet Collection) (Tshudy and Sorhannus, 2000a, fig. 3.4; Figure 3F here). If so, this “impreinte” is no longer distinct on the actual specimen.
Bosquet’s plate 10, figure 61 (Figure 3C here, mirrored) depicts a cephalothorax (Van Breda Collection) that is similar to that of plate 10, figure 5, but likewise with unknown repository data. This illustration resembles IRScNB90-17 (Tshudy and Sorhannus, 2000a, fig. 3.5; Figure 3E here) of the Ubaghs Collection.
Oncopareia (sensu Bosquet, 1854) included a Hoploparia-like (not Thaumastocheles-like) pleon, one that Bosquet (1854, p. 128) said bore a close resemblance to that of Nephrops norvegicus (Linnaeus, 1758). It seems that Bosquet (1854) did not recognize the Thaumastocheles-like pleons as being “real” and informative. It would seem that he attributed the lateral carinae on the pleon to deformation; his footnote on p. 129 says that all of the specimens found are more or less compressed or deformed in the matrix. Specimens IRScNB90-35c (Ubaghs Collection) (Tshudy, 1993, figs. 38.2, 3; Tshudy and Sorhannus, 2000a, figs. 3.2-3.3; Figure 3H-I here) and IRScNB 90-23b (Ubaghs Collection) (Tshudy, 1993, fig. 38.6; Tshudy and Sorhannus, 2000a, fig. 3.6; Figure 3J here) show the distinctive, Thaumastocheles-like terga and short pleura, separated by a prominent ridge.
Bosquet made no mention of the single Thaumastocheles-like claw pair in his collection (IRScNB 90-36a), likely because very little of the distinctive major claw is exposed. There may be others in the Bosquet Collection that we have not seen. The Ubaghs Collection contains several Thaumastocheles-like claws, some in obviously heterochelous pairs. Specimen IRScNB 90-28 (Tshudy, 1993, fig. 39.1; Tshudy and Sorhannus, 2000a, figs. 4.5-4.8; Figure 3K here) constitutes a heterochelous claw pair, including a Thaumastocheles-like pectinate claw (fingers only). Specimen IRScNB 90-27b (Tshudy, 1993, fig. 39.3; Tshudy and Sorhannus, 2000a, fig. 3.3; Figure 3N here) is a complete, Thaumastocheles-like pectinate claw, with bulbous palm and delicate fingers bearing acicular dentition. Specimen IRScNB 90-19f (Tshudy, 1993, fig. 39.5-8) is another bulbous, Thaumastocheles-like palm, while specimen IRScNB 90-3b (Tshudy, 1993, fig. 39.2; Tshudy and Sorhannus, 2000a, fig. 3.2; Figure 3L here) shows acicular dentition. Finally, specimen IRScNB 90-32a (Tshudy, 1993, fig. 39.4; Tshudy and Sorhannus, 2000a, fig. 3.4; Figure 3M here) is a nearly complete crusher claw. The crusher claw in Bosquet’s plate 10, figure 7 (Bosquet Collection) and the claw in plate 10, figure 71a, b (Van Breda Collection) are referred herein to Hoploparia beyrichi (see below).
The other species of Oncopareia recognized herein are confidently referred to the genus. Published illustrations reveal nothing to distinguish O. coesfeldiensis from O. bredai, leading us to synonymize both taxa. The holotype of O. esocina suggests that its pleonal pleura are bordered by a marginal carina, narrower than that on O. bredai, while O. lunata has a slightly different groove pattern. Oncopareia klintebjergensis differs from O. bredai in several aspects, including the longer, shallower orbits, the more ventral position of prominence “ω” (omega) on the carapace, and in the claw denticles which are directed more sharply distally.
Oncopareia bredai Bosquet, 1854
Figure 1D, Figure 2A, Figure 3A-N, Figure 4A-E, Figure 5A-L, Figure 6A-C, Figure 7A-K
partim 1854 Oncopareia bredai Bosquet, p. 128, pl. 10, figs. 5, 61 and 8 [non pl. 10, figs. 6, 7 and 71a, b = Hoploparia beyrichi ].
1862 Nymphaeops coesfeldiensis Schlüter, p. 728, pl. 13, figs. 3, 6.
 1868 Nymphaeops coesfeldiensis Schlüter; Schlüter in von der Marck and Schlüter, p. 295.
1868 Nymphaeops coesfeldiensis Schlüter; Schlüter in von der Marck and Schlüter, p. 295.
1868 Enoploclytia paucispina Schlüter in von der Marck and Schlüter, p. 303, pl. 44, fig. 6.
1879 Nymphaeops coesfeldiensis Schlüt.; Schlüter, p. 597, pl. 15, fig. 1, 2.
1886 Homarus bosqueti Pelseneer, p. 166 [6], fig. 4.
partim 1887 Hoploparia biserialis Fritsch in Fritsch and Kafka, p. 35, pl. 5, fig. 2 [non text-fig. 56; pl. 3, fig. 5, pl. 5, fig. 1 = Hoploparia biserialis ].
1929 Nymphaeops coesfeldiensis Schlüter; Glaessner, p. 279.
1941 Oncopareia bredai Bosquet; Mertin, p. 183, fig. 9 [cephalothorax, but not pleon, and necessarily rostrum]
 ?1941 Oncopareia coesfeldensis [sic] (Schlüter); Mertin, p. 182, text-figs. 9a, ?9b, 10b-d, k-m; pl. 3, figs. 2-7.
?1941 Oncopareia coesfeldensis [sic] (Schlüter); Mertin, p. 182, text-figs. 9a, ?9b, 10b-d, k-m; pl. 3, figs. 2-7.
?1941 Oncopareia cf. coesfeldensis [sic] (Schlüter); Mertin, p. 187, pl. 3, fig. 9.
1964 Ctenocheles coesfeldiensis (Schlüter); Secretan, p. 152.
1990 Oncopareia bredai Bosquet; Feldmann et al., p. 26, text-fig. 4; pl. 1, figs. 4-6.
2000a Oncopareia bredai Bosquet; Tshudy and Sorhannus, p. 484, figs. 3, 4.
2000b Oncopareia bredai Bosquet; Tshudy and Sorhannus, fig. 2.2.
2010 Oncopareia bredai Bosquet; Schweitzer et al., p. 31.
2010 Oncopareia coesfeldiensis (Schlüter); Schweitzer et al., p. 31.
2016 Oncopareia bredai Bosquet; Feldmann et al., p. 23.
2016 Oncopareia coesfeldiensis (Schlüter); Feldmann et al., p. 23, fig. 20.3b.
Type material. IRScNB 90-33f (lectotype; specimen consisting of cephalothorax, partial pleon, left [crusher] palm, and basis through merus of right claw), IRScNB 90-3b (Tshudy, 1993, fig. 39.2; incomplete cheliped); IRScNB 90-15a, b (Tshudy, 1993, fig. 38.4; cephalothorax fragment), IRScNB 90-17 (Tshudy, 1993, fig. 38.5; cephalothorax), IRScNB 90-18, 90-19b, c, g, i, k, q (Tshudy, 1993, fig. 39.5-8; incomplete cheliped), IRScNB 90-22a, c, 90-23b (Tshudy, 1993, fig. 38.6; partial pleon), IRScNB 90-26a, 90-27a, b (Tshudy, 1993, fig. 39.3; b: complete cheliped), IRScNB 90-28a, b (Tshudy, 1993, fig. 39.1; incomplete pair of chelipeds), IRScNB 90-32a, c, e (a: cheliped; c: cephalothorax [Tshudy, 1993, fig. 38.1]), IRScNB 90-34, 90-35a, c (Tshudy, 1993, fig. 38.2-3; pleon) and IRScNB 90-36a, 90-37 and 90-40.
 Bosquet (1854, pl. 10, figs. 5-8) did not explicitly select a type when erecting Oncopareia bredai, but did illustrate five specimens, all unregistered. However, he did cite collections by name. As Bosquet did not designate type specimens, his illustrated specimens were not identified by museum/specimen numbers and none of the specimens that we have examined could be confidently matched up with his illustrations, it seems best to designate a lectotype for Oncopareia bredai. Specimen IRScNB 90-33f (Figure 3G here), from the “type series” of O. bredai (sensu Bosquet, 1854), according to the label accompanying the specimen, is designated lectotype here. The specimen consists of a cephalothorax, partial pleon, left (crusher) palm, and basis through merus of the right claw. This specimen serves well as the lectotype because it includes a cephalothorax and its associated, distinctive, Thaumastocheles-like pleon.
Bosquet (1854, pl. 10, figs. 5-8) did not explicitly select a type when erecting Oncopareia bredai, but did illustrate five specimens, all unregistered. However, he did cite collections by name. As Bosquet did not designate type specimens, his illustrated specimens were not identified by museum/specimen numbers and none of the specimens that we have examined could be confidently matched up with his illustrations, it seems best to designate a lectotype for Oncopareia bredai. Specimen IRScNB 90-33f (Figure 3G here), from the “type series” of O. bredai (sensu Bosquet, 1854), according to the label accompanying the specimen, is designated lectotype here. The specimen consists of a cephalothorax, partial pleon, left (crusher) palm, and basis through merus of the right claw. This specimen serves well as the lectotype because it includes a cephalothorax and its associated, distinctive, Thaumastocheles-like pleon.
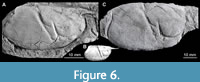
Additional material. Four specimens from the upper lower Maastrichtian (Gulpen Formation, Vijlen Member, interval 6) of Haccourt, north-east Belgium, originally described by Feldmann et al. (1990), i.e., a near-complete cephalothorax and pleon with fragments of appendages (USNM 444296), an incomplete pleon with tailfan (USNM 444297), an incomplete cephalothorax with fragments of pleon and appendages (USNM 444298), as well as a fragmentary cephalothorax and chelipeds (USNM 444299). From the same locality and stratigraphical level come seven additional specimens (Figure 5), preserving partial cephalothoraxes, pleons, claws, and partial tailfans (NHMM JJ 6741, JJ 6764, JJ 6839, JJ 7194, JJ 14618, JJ 15534, and JJ 15836). One claw (NHMM JJ 5932) is from the Zeven Wegen Member (Gulpen Formation, late Campanian) at the CBR-Lixhe quarry (Lixhe, north-east Belgium). One specimen, a cephalothorax with associated pleonal segments 1 and 2 and both claws (MNHN R03408; Figure 4) is from the upper Maastrichtian (Kunrade Formation) at Kunrade, the Netherlands. Two syntypes of Hoploparia biserialis Fritsch, in Fritsch and Kafka, 1887 (NM O3470 and NM O6861) originated from the Turonian of Vinary, Czech Republic, and representing isolated cephalothoraxes (Figure 6); NM O3470 was figured by Fritsch and Kafka (1887, pl. 5, fig. 2). These two specimens are removed from the type series of H. biserialis herein (see below for further details).
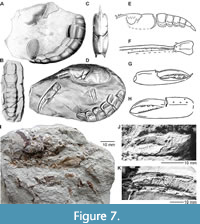 The only material referred to Nymphaeops coesfeldiensis (Figure 7) that we have studied in person includes a few fragmentary specimens of pectinate and crusher claw (NHMM HB 393 and HB 756; Figure 7J–K) from the Vaals Formation (lower Campanian) of Haccourt, north-east Belgium, recorded by Jagt and Bongaerts (1986). In addition, there is a single, near-complete cephalothorax, with associated appendages (NHMM JJ 9974), from the same locality and stratigraphical level (Figure 7I). In this respect, it is of note that Bosquet (1860, 1868) listed, but did not illustrate, O. bredai, from the so-called ‘Hervien’ in the Liège-Limburg Basin. This corresponds nowadays with the Vaals Formation of early Campanian age. We have attempted to borrow Schlüter’s 1862 type specimen and other individuals from the ‘Akademie zu Münster’, but these may have fallen victim to air raids during World War II. We are aware of larger suites of specimens of nephropid lobsters contained in the Karl-Heinz Hilpert Collection at the Ruhrmuseum (Essen, Germany; see Jagt et al., 2004; Van Bakel et al., 2005) that may include topotypical material of Nymphaeops coesfeldiensis. A detailed assessment of this material is deferred to another occasion.
The only material referred to Nymphaeops coesfeldiensis (Figure 7) that we have studied in person includes a few fragmentary specimens of pectinate and crusher claw (NHMM HB 393 and HB 756; Figure 7J–K) from the Vaals Formation (lower Campanian) of Haccourt, north-east Belgium, recorded by Jagt and Bongaerts (1986). In addition, there is a single, near-complete cephalothorax, with associated appendages (NHMM JJ 9974), from the same locality and stratigraphical level (Figure 7I). In this respect, it is of note that Bosquet (1860, 1868) listed, but did not illustrate, O. bredai, from the so-called ‘Hervien’ in the Liège-Limburg Basin. This corresponds nowadays with the Vaals Formation of early Campanian age. We have attempted to borrow Schlüter’s 1862 type specimen and other individuals from the ‘Akademie zu Münster’, but these may have fallen victim to air raids during World War II. We are aware of larger suites of specimens of nephropid lobsters contained in the Karl-Heinz Hilpert Collection at the Ruhrmuseum (Essen, Germany; see Jagt et al., 2004; Van Bakel et al., 2005) that may include topotypical material of Nymphaeops coesfeldiensis. A detailed assessment of this material is deferred to another occasion.
Diagnosis. Oncopareia with pleura having prominent, rounded, marginal carina; intercervical groove extending toward below upper end of cervical groove; pectinate claw denticles perpendicular to longitudinal axis of fingers.
Emended description. Carapace laterally compressed, twice as long as high. Postcervical groove intersecting dorsal median line slightly posteriorly to mid-length on carapace. Postcervical groove long, well incised dorsally and laterally; extending ventrally over dorsal surface, then, as combined branchiocardiac-postcervical groove, angling anteroventrally to hepatic groove. Hepatic groove deeply incised, joining cervical and antennal grooves in semicircular loop. Cervical groove originating at level of base of orbit, paralleling postcervical groove except at upper end of cervical groove, which is deflected anteroventrally. Prominence “ω” distinct. Intercervical groove narrow, extending anteroventrally from postcervical groove, at level just above cervical groove origin, toward, but not reaching, lower portion of cervical groove; intercervical groove terminating at level of mid-height on cervical groove. Cervical groove and antennal groove together forming a smooth, semicircular arc. Antennal groove vague anterior to prominence “ω”; curving toward antennal spine, effacing well before reaching spine. Branchiocardiac carina broad, subtle; extending from near posterior margin to postcervical groove, curving downward near groove. Antennal region flat, featureless. Very shallow branchiocardiac groove present; most evident near postcervical groove. Posteromarginal carina wide, smooth, with a few pits. Cephalic region granulose, with a row of setal pits extending from top of cervical groove to near antennal spine. Antennal spine strong; supraorbital and postorbital spines slightly smaller. Thoracic region pitted dorsally, with granules generally sparse laterally but dense ventrally.
Pleonal terga wide, convex. Each segment with lateral carina at tergum-pleuron boundary. Pleura short; at least pleura 2 and 3 with blunt ventral margin.
Tergum 2 rectangular. Anterior margin straight, posterior margin subtly convex backward, lateral margins subtly convex, locally indented near posterior margin. Anterior margin depressed laterally; depressed area progressively widening toward pleuron. Tergum coarsely and densely pitted. Lateral carina prominent, rounded, widest anteriorly. Pleuron trapezoidal, convex axially; upper surface coarsely pitted, lower surface smooth.
Terga 3-6 of similar width. Tergum 3 rectangular. Anterior margin slightly concave forward, lateral margins slightly convex, widest at mid-length, subtly indented locally near posterior margin. Anterior margin depressed laterally. Tergum coarsely, densely pitted except on smooth dorsomedian keel, depression along anterior margin, and in subtle furrow adjacent to upper limit of lateral carina. Summit of lateral carina well defined, extending obliquely ventrally posteriorly. Pleuron trapezoidal; ventral and posterior margins shorter than dorsal and anterior margins. Pleuron concave overall; margins convex. Surface mostly smooth, granulose ventrally. Surface locally swollen at anteroventral corner, maybe with spine.
Tergum 4 trapezoidal, wider anteriorly than posteriorly. Lateral margins convex along most of length; straight for a short distance posteriorly. Tergum coarsely and densely pitted except on smooth dorsomedian keel. Lateral carina extending from near anterior margin to posterior margin, curving posteroventrally over anterior half and extending on level over posterior half. Upper surface of carina very broad, inflated. Carina summit rounded, distinct. Pleuron margins convex, with sharply rounded, posteroventrally directed termination. Upper surface almost smooth, lower surface granulose. Small, dome-like swelling anteroventrally.
Tergum 5 rectangular. Lateral margins convex. Tergum smooth on dorsomedian keel, otherwise coarsely and densely pitted. Lateral carina as on segment 4. Pleuron round. Small, dome-like swelling anteroventrally. Pleuron upper surface granulose, lower surface densely granulose. Pleural margins 4-5 with very narrow, rounded, raised margin.
Sixth tergum trapezoidal, much wider anteriorly. Lateral margins slightly convex. Tergum subtly convex, densely and coarsely pitted, sparsely so dorsoventrally. Tergum dropping off sharply laterally. Posteriorly, tergum concave laterally between subtly convex dorsum and drop off. Pleuron triangular, very short, highest near anteroventral corner. Small, dome-like swelling anteroventrally. Pleuron granulose. Telson with round granules.
Claws strongly dissimilar in form; relative size of left and right claw varying. Crusher claw very Hoploparia-like; delicate pectinate claw with bulbous palm, long fingers and acicular dentition much like that of extant Thaumastocheles.
Pectinate claw with bulbous palm approximately twice longer than thick. Large process on at least one side of palm receiving spine at base of dactylus. Palm surface with minute granules. On at least one surface of palm, spine distally near base of fixed finger. Fingers long, slender, with acicular denticles of varying length, of formula ADCDBDCDA (“A” being largest), in one plane. Fixed finger nearly straight, widest at base; width tapering very gradually distally. Dactylus slightly curved; widest over proximal portion near mid-length. Fingers proximally triangular in cross section; otherwise compressed. Tips of fingers not occludent; one slightly shorter than other. Most denticles straight, angling slightly distally. Some larger denticles slightly curving, angling distally at base, proximally at end. Surface of both fingers pitted.
Crusher palm shape varying between specimens. On most, left palm nearly twice as long as wide. Palm widening distally over proximal half; margins parallel over distal half. Palm compressed, slightly thicker proximally; upper and lower surfaces gently convex. Palm surface variously granulose, pitted. Upper and lower palm surfaces very subtly concave at base of fixed finger. Both surfaces with spine submedially, near base of fixed finger. Submedial process on upper and lower distal margin articulating with spines on dactylus base. Inner margin somewhat flattened, with a few alternating spines on upper and lower edges. Outer margin rounded; axis with band of setal pits; upper surface of outer margin with irregular row of spines directed slightly distally. Fingers moderately strong, ovate in cross section. Fixed finger occlusal surface with multi-cusped platform proximally; otherwise with blunt, conical denticles. Dactylus with upward and distally-directed spine proximally on corner of upper/non-occluding surfaces. Dactylus dentition conical; pattern varying between specimens. Merus (IRScNB 90-17 and 90-33f) large, strongly compressed laterally, widening distally; upper proximal corner terminating in long point. Upper palm surface variously pitted, granulose.
On two specimens (IRScNB 90-33f and 90-32a; latter with small, Thaumastocheles-like claw), crusher claw palm proportionately shorter than on other specimens, and widening distally over entire length, giving subtriangular outline; upper and lower surfaces more convex than on other specimens; palm thickens distally.
Remarks. Pelseneer (1886) was the first to note the Thaumastocheles-like morphology in Bosquet’s type series. He examined both the Bosquet and Ubaghs collections but did not recognize Bosquet’s combining of the two lobster species. He did, however, realize the uniqueness of one of the Thaumastocheles-like pleons and referred to this as a new species, Homarus bosqueti. Pelseneer (1886) correctly described the terga as being less convex than those of O. bredai, but his figures 3 and 4 show the opposite. Homarus bosqueti is considered herein to be a junior objective synonym of Oncopareia bredai.
A re-examination of the syntype series of Hoploparia biserialis has revealed that two isolated cephalothoraxes assigned to this species by Fritsch himself (one of them being figured in Fritsch and Kafka, 1887, pl. 5, fig. 2), belonged to a different genus. Indeed, there is an imminent difference in the groove pattern between the two cephalothoraxes assigned to H. biserialis shown in Fritsch and Kafka (1887, pl. 5, figs. 1 and 2), one of them belonging to H. biserialis and another being reassigned to O. bredai herein. We were not able to identify any morphological differences between the type material of O. bredai and the Czech material here attributed to that species.
The type specimen of Nymphaeops coesfeldiensis has not been located; it is here considered lost. Our examination of the original figures (Schlüter, 1862, pl. 13, figs. 3, 6; Schlüter, 1879, pl. 15, figs. 1, 2, refigured herein in Figure 7) has not revealed any difference from the material of O. bredai leading us to recognize N. coesfeldiensis as a junior subjective synonym of O. bredai. In this respect, it is important to note that Mertin (1941) lumped material from several ‘Senonian’ localities into one species, i.e., O. coesfeldiensis. Whether or not all of the specimens Mertin (1941) assigned to O. coesfeldiensis are correctly referred to one species is uncertain. His unwhitened photographs are difficult to interpret and his line drawings do not seem to be consistent with the photographs. For example, he did not connect the branchiocardiac and cervical grooves ventrally (as hepatic groove) in his figure 9a. Cephalothoraxes and pleons in his plates strongly resemble those of O. bredai as revised herein. Mertin (1941, p. 187, pl. 3, fig. 9) also listed Oncopareia cf. coesfeldiensis from the lower ‘Senonian’ of north-central Germany and Oncopareia sp. (Mertin, 1941, p. 187, text-fig. 10e, f and n) from the upper ‘Senonian’ of the same area. The latter shows heterochelous claws, the pectinate claw having acicular dentition.
Oncopareia bredai differs from its congeners by the intercervical groove extending toward below the top of the cervical groove. In O. bredai, the marginal carina on pleonal pleura is wider than that on O. esocina. The pectinate claw of O. bredai possesses denticles which are perpendicular to the longitudinal axis of the fingers, unlike those in O. klintebjergensis which are angled distally.
Range and occurrence. Oncopareia bredai was first described from what is now the Kunrade Formation, a lateral equivalent of part of the Maastricht Formation, of late Maastrichtian age, and exposed in the eastern part of southern Limburg, the Netherlands. Subsequent records are from the Vijlen Member, interval 6 (Gulpen Formation), of late early Maastrichtian age, as exposed at the former SA Ciments Portland Liegeois (CPL) quarry near Haccourt and the adjacent Ciments Belge Réunie (CBR) quarry, province of Liège, north-east Belgium. Czech occurrences, referred to Hoploparia biserialis by Fritsch, in Fritsch and Kafka (1887) originate from the uppermost part of the Jizera Formation (upper Turonian) at Vinary near Vysoké Mýto in the Bohemian Cretaceous Basin, Czech Republic. The stratigraphical level of N. coesfeldiensis was indicated as ‘Senonian’, both lower and upper, of north-central and northwest Germany by Mertin (1941); in current terminology, this would correspond to the lower and upper Campanian.
Oncopareia esocina (Fritsch in Fritsch and Kafka, 1887)
Figure 8A-E
1887 Stenocheles esocinus Fritsch in Fritsch and Kafka, p. 40, text-fig. 59; pl. 4, fig. 7.
 1887 Ischnodactylus esocinus (Fritsch in Fritsch and Kafka); Forir, p. 189.
1887 Ischnodactylus esocinus (Fritsch in Fritsch and Kafka); Forir, p. 189.
1893 Stenocheles esocinus Fritsch in Fritsch and Kafka; Frič, p. 106, text-fig. 137.
1894 Stenocheles esocinus Fritsch in Fritsch and Kafka; Frič, p. 102, text-fig. 137.
1929 Ischnodactylus esocinus (Fritsch in Fritsch and Kafka); Glaessner, p. 226.
1941 Oncopareia esocinus (Fritsch in Fritsch and Kafka); Mertin, p. 181, text-fig. 8b.
?1941 Oncopareia cf. esocinus (Fritsch in Fritsch and Kafka); Mertin, p. 181, text-fig. 8c; pl. 3, fig. 1.
2010 Oncopareia esocinus (Fritsch in Fritsch and Kafka); Schweitzer et al., p. 31.
2016 Oncopareia esocinus (Fritsch in Fritsch and Kafka); Feldmann et al., p. 23, fig. 20.3a.
Material examined. Part and counterpart of the holotype, NM O3467 and NM O3468 (respectively) in the palaeontological collections of the National Museum in Prague.
Diagnosis. Oncopareia with second pleonal somite tergite-pleuron boundary marked by arcuate, concave-upward, ridge. Second pleuron quadrate.
Remarks. This species is known from a single specimen representing a pleon and associated major claw, both of which are characteristic of Oncopareia. Pleonal terga are wide and convex with lateral carina at tergum-pleuron boundary on each segment. The pectinate claw with bulbous palm possesses long, slender fingers with acicular dentition. The material is flattened in fine-grained siliceous to calcareous claystones. The second pleonal somite on O. bredai differs from that of O. esocina in being shorter and less quadrate, and in having the tergite-pleuron boundary ridge straighter. Also, pleura bordered by marginal carina are narrower on O. esocina than those on O. bredai.
Range and occurrence. This species is known from the lower Coniacian (Cremnoceramus crassus inoceramid Zone) of the Teplice Formation (Rohatce Member) at Oškobrh near Poděbrady in the Czech Republic (Fritsch and Kafka, 1887). The material reported by Mertin (1941, p. 181) from the lower ‘Senonian’ (i.e., Coniacian-lower Campanian) of Salzberg near Quedlinburg, northern Germany, is certainly referable to Oncopareia, although based solely on Mertin’s photographs of an unwhitened specimen, we cannot judge whether this belongs to O. esocina, or not.
Oncopareia klintebjergensis Jakobsen and Collins, 1979
Figure 8F-G
1979 Oncopareia klintebjergensis Jakobsen and Collins, p. 61, pl. 1, figs. 1, 2.
2010 Oncopareia klintebjergensis Jakobsen and Collins; Schweitzer et al., p. 32.
2016 Oncopareia klintebjergensis Jakobsen and Collins; Feldmann et al., p. 23, fig. 20.3c-d
Material examined. Holotype MGUH 14437 (incomplete cephalothorax), paratype MGUH 14438 (right cheliped fingers).
Diagnosis. Oncopareia with long, almost straight orbital margin; cervical and combined branchial-postcervical grooves concave-forward and with two spines behind orbit.
Remarks. Oncopareia klintebjergensis is based on the anterior portion of a single cephalothorax and part of its associated right cheliped (Jakobsen and Collins, 1979). The species differs from O. bredai in several aspects, including the longer, shallower orbits, the more ventral position of prominence “ω” on the carapace, and in the claw denticles which are angled distally.
Range and occurrence. Paleocene Lellinge Greensand Limestone of Klintebjerg, eastern Sjælland, Denmark.
Oncopareia lunata (Fritsch in Fritsch and Kafka, 1887)
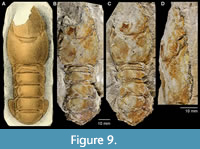
Figure 9A-D
1887 Nymphaeops? lunatus Fritsch in Fritsch and Kafka, p. 35, pl. 5, fig. 6.
 1929 Nymphaeops? lunatus; Glaessner, p. 279.
1929 Nymphaeops? lunatus; Glaessner, p. 279.
1941 Oncopareia lunatus (Fritsch in Fritsch and Kafka); Mertin, p. 180.
2000a Oncopareia? lunatus (Fritsch in Fritsch and Kafka); Tshudy and Sorhannus, pp. 481, 484-appendix 1.
2010 Oncopareia lunatus (Fritsch in Fritsch and Kafka); Schweitzer et al., p. 32.
Material examined. Holotype, NM O3473, in the palaeontological collections of the National Museum in Prague.
Diagnosis. Oncopareia with arcuate (concave-up) intercervical groove extending toward upper part of cervical groove.
Description. This species is known by a single specimen preserving part of a dorsoventrally flattened cephalothorax (without rostrum and anterior region) and the pleon in dorsal aspect. The pleon itself is connected to the cephalothorax. Long postcervical groove intersects dorsal median line slightly posteriorly to mid-length on carapace; as combined branchiocardiac-postcervical groove, extending anteroventrally to hepatic groove; then, bifurcating at third-height of carapace and as arcuate intercervical groove curving toward, but not reaching, the level of top of cervical groove. Branchiocardiac groove absent posteriorly and beginning as branchiocardiac-postcervical groove; after bifurcating extending ventrally to join semicircular hepatic groove. Cervical groove well incised, short. Anteroventral part of cephalothorax, in region of antennal and inferior grooves, not preserved. Pleura short; pleonal terga wide, convex with a lateral carina at tergum-pleuron boundary.
Remarks. Mertin (1941) referred this species to Oncopareia based on the short pleura; he did not comment on the cephalothorax grooves. Tshudy (1993, pp. 300-301), based on the published illustration, assigned the single specimen with Oncopareia-like pleon to “ Oncopareia ?”; the uncertainty was based on the urogastric groove on the cephalothorax, which is unknown in Oncopareia. Personal re-examination of the type material by one of us (MKV) confirmed the attribution of the species to Oncopareia. In this respect, it should be stressed that the original restoration of this form (Fritsch and Kafka, 1887, pl. 5, fig. 6) does not fully correspond with the original specimen. Fritsch pencil marked the specimen to highlight morphological features, and, as a result, two distinct transverse grooves crossing the dorsal midline of the cephalothorax appeared in his drawing. These grooves look to be the urogastric and postcervical grooves, the former being known in several nephropid genera, but not in Oncopareia. However, after washing the specimen, it turned out that Fritsch marked an extra line (i.e., a potential urogastric groove) on this particular specimen. In actual fact, there is only one transverse groove crossing the dorsal midline, the postcervical groove. Thus, caution must be exercised when dealing with Fritsch’s taxa on the basis of published drawings only.
Oncopareia lunata differs from O. bredai in having a considerably shorter cervical groove, a more arcuate intercervical groove oriented toward the level of the top of the cervical groove. In addition, it lacks a posteriorly defined branchiocardiac groove. In view of the fact that O. lunata is incompletely known (i.e., lacking the anterior region of the cephalothorax and chelipeds), further comparison with O. bredai is not possible.
Range and occurrence. The occurrence of O. lunata is so far limited to shallow-marine marlstones of the Bílá Hora Formation (lower–middle Turonian) in the Prague area, Czech Republic.
Oncopareia sp. 1
1993 Oncopareia sp. Tshudy, p. 303.
2000a Oncopareia bredai Bosquet; Tshudy and Sorhannus, figs. 4.5-4.8.
Material examined. Three specimens, IRScNB 90-19f, 90-19h and 90-19m (one complete and two incomplete palms).
Description. Palm bulbous, about 1.7x longer than wide; widest and thickest near distal end. Upper and lower surfaces convex. Outer margin rounded; inner margin with very prominent, broad, flat-topped keel. Keel higher, narrower, proximally; lower, broader distally. Palm upper surface with very fine (IRScNB 90-19f) or fine (IRScNB 90-19m) granules, with spine near outer-proximal corner. Lower surface and outer margin smooth. Inner marginal keel bearing a few stout, alternating spines on keel edges; otherwise with minute, densely distributed granules.
Remarks. These specimens are distinguished from O. bredai on the basis of the prominent keel on the inner margin of the claw.
Range and occurrence. Upper Maastrichtian (Kunrade Formation) of Kunrade, southern Limburg, the Netherlands.
Oncopareia sp. 2
1941 Oncopareia sp. Mertin, p. 187, text-fig. 10e, f, n.
Material examined. None.
Remarks. Known only by an incomplete major and minor claw. Solely on the basis of Mertin’s (1941) photographs of unwhitened specimens, we cannot judge whether this material belongs to O. esocina, or not. However, it certainly is referable to Oncopareia.
Range and occurrence. Upper ‘Senonian’ of Königslutter, east of Braunschweig, north-central Germany (Mertin, 1941); in current terminology this would correspond to the upper lower Campanian.
Genus Hoploparia M’Coy, 1849
Type species. Astacus longimanus Sowerby, 1826, by the subsequent designation of Rathbun (1926).
Diagnosis (after M’Coy, 1849, pp. 175-176). Carapace minutely granulose, oblong, tumid, slightly compressed, a little deeper than wide, ending in front in a strong sharp rostrum, the sides of which are strongly carinate and smooth, or with few very minute teeth; beneath the orbits the cheeks are prolonged forwards about half the length of the rostrum.
Diagnosis (after Feldmann et al., 2016, p. 16). Subcylindrical carapace smooth; rostrum long, with suprarostral spines; supraorbital spine present; scaphocerite present; subdorsal carina present; cervical groove and postcervical groove well-developed, postcervical groove extending from midline to about midheight of carapace where it joins hepatic groove to encircle adductor testis muscle attachment; branchiocardiac groove weak; strong heterochely of pereiopod 1; major chelipeds with fingers with strong, domal denticles on occlusal surfaces; minor chelipeds with finer, spinose denticles on occlusal surfaces; pereiopods 2 and 3 chelate; terga smooth or weakly ornamented; subrectangular telson; exopodite of uropods with diaeresis.
Diagnosis (after Tshudy et al., 2018, p. 174). Rostrum long, spinose. Postcervical groove well impressed over most of length; becomes subtler as it extends anteriorly toward cervical groove. Branchiocardiac groove (dorsally) usually present. Ventral extension of branchiocardiac groove typically extends to hepatic groove. Urogastric groove typically absent. Cervical groove well impressed; extends from level of orbit to junction of hepatic and antennal grooves. Median carina typically absent. Submedian carina present on a few species. Subdorsal carina present. Supraorbital spine present, typically followed by supraorbital carina. Postorbital spine typically present; gastrolateral and hepatic spine typically absent. Antennal carina absent, short, or extending to near cervical groove. Thoracic region lacks carinae. Pleonal terga unornamented, mostly unsculptured; typically with a narrow transverse furrow along posterior margin. Pleura elongate, typically cordate, ending in a point. Telson surface with a pair of submedian ridges converging posteriorly. Telson without lateral spines but with posterolateral spines. Scaphocerite present. Claws typically unequal; upper and lower surfaces lacking carinae. Exoskeleton generally granulated.
Remarks. Hoploparia was erected by M’Coy (1849) for fossils that generally resembled Homarus but had a more sculptured (grooved, locally inflated, etc.) and ornamented carapace. In fact, the name Hoploparia refers to the lobster’s “spiny cheeks”, the locally inflated and ornamented regions on the anteroventral sides of the cephalothorax. Homarus lacks these locally inflated “cheeks” and is less spiny. M’Coy’s diagnosis, reprinted here, distinguished the type species, Hoploparia longimana, from Homarus.
Hoploparia is readily distinguished from Oncopareia in the nature of the chelipeds and pleon. Claws are never pectinate in Hoploparia and pleura 3–5 in Oncopareia are characteristically quadrate, not cordate and pointed as in Hoploparia.
Hoploparia beyrichi Schlüter, 1862
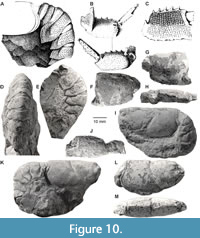
Figure 1E, Figure 2B-C, Figure 10A-M
Partim 1854 Oncopareia bredai Bosquet, p. 127, pl. 10, figs. 6, 7, 71 [not pl. 10, figs. 5, 61 ]
 1862 Hoploparia beyrichi Schlüter, p. 721, pl. 13, fig. 4.
1862 Hoploparia beyrichi Schlüter, p. 721, pl. 13, fig. 4.
1886 Homarus bredai (Bosquet); Pelseneer, p. 164, figs. 2, 3.
1929 Hoploparia bredai (Bosquet); Glaessner, p. 216.
1941 Oncopareia bredai Bosquet; Mertin, fig. 9e [pleon only].
1993 Hoploparia beyrichi Schlüter; Tshudy, figs. 17.1–17.14.
2000b Hoploparia beyrichi Schlüter; Tshudy and Sorhannus, fig. 2.3.
2010 Hoploparia beyrichi Schlüter; Schweitzer et al., p. 28.
2018 Hoploparia beyrichi Schlüter; Tshudy et al., p. 182.
Material examined. Holotype, Museum für Naturkunde Berlin, MB. A. 209; also several specimens deposited in the IRScNB collections, including those in the Bosquet (IG 4285; IRScNB 90-33a, c, g, i), De Jaer (IG 8261; IRScNB 90-24a, b) and Ubaghs (IG 6521; IRScNB 90-19j, n, 90-20, 90-22b, 90-32b, d, f and 90-39c) collections.
Diagnosis. Cephalothorax with little relief; antennal carina absent. Postorbital and supraorbital spine prominent; supraorbital followed by short carina. Pleonal terga and pleura with little relief; surfaces pitted. Claws slightly unequal in form and size.
Description. Carapace laterally compressed, twice as long as high. Postcervical groove intersecting dorsal midline at just posterior to mid-length on carapace. Postcervical groove extending transversely across dorsal surface; combined branchiocardiac and postcervical groove extending anteroventrally to level of antennal groove in slightly concave-forward arc; well impressed dorsally, becoming progressively fainter ventrally. Anterior extension of postcervical groove extending toward, but not reaching, middle of cervical groove. Hepatic groove very subtle or absent. Cervical groove subparallel to, and more vertical than, postcervical groove; originating at level of base of orbit. Cervical and antennal grooves together forming a semicircular arc. Antennal groove anteriorly curving upward, reaching carapace margin well below antennal spine. Branchiocardiac groove very short on most specimens, long on the single large specimen. Supraorbital spine followed by short carina. Postorbital spine round. Wide, flat, posteromarginal carina with elongate pits dorsally, smooth or pitted laterally. Surface of thoracic region with transversely elongate pits dorsally and laterally, granules ventrally.
Pleon laterally compressed. Terga rectangular, strongly convex, sparsely pitted. Pleura long, more densely pitted than terga. Pleuron 2 quadrate; anterior margin rounded, anteroventral corner rounded, ventral margin straight, posterior margin recurved, posteroventral corner terminating in posteriorly directed point. Pleura 3–5 cordate, terminating in point; pleuron 6 triangular. Pleura gently inflated axially, traversed by very broad, subtle, oblique ridge extending posteroventrally from anterodorsal corner of pleuron; carina most pronounced on second pleuron and becoming progressively less obvious on posterior segments. Pleura directed posteriorly relative to terga. A single, large pit present posteroventrally on pleuron 2, anteroventrally on pleura 3-5.
Claws slightly unequal in form and size. Right palm larger than left; right palm slightly longer than wide; left palm slightly more elongate. Palms widening distally; rapidly so near proximal articulation. Right palm upper surface gently convex; pitted proximally, but most of surface with flat, scale-like granules of varying size; pits along distal edges of most scales. Lower surface slightly more convex than upper surface, mostly pitted. Inner margin angular, slightly upturned, with at least four large, conical, slightly upturned spines; all but most proximal spine directed slightly distally; space between spines increasing distally. Outer margin angular, pitted.
Left palm upper surface gently convex, pitted. Lower surface convex, with very fine, very low, transversely elongate granules. Inner margin angular, with at least four large, conical, slightly upturned spines; all but most proximal spine directed distally. Outer margin broadly rounded proximally, becoming progressively narrower distally; sharply rounded at base of fixed finger.
Remarks. The holotype from the upper Maastrichtian (Kunrade Formation) of Kunrade, southern Limburg, the Netherlands, consists of an imperfectly preserved cephalothorax lacking the rostrum and a slightly better-preserved pleon lacking the telson. Schlüter (1862, p. 722) considered the specimen distinct from O. bredai, mainly on the basis of its carapace grooves and ornamentation and named it Hoploparia beyrichi. The pleons and claws that Bosquet (1854) described, illustrated and referred to Oncopareia bredai also belong to H. beyrichi; thus, some specimens from the syntype collection of O. bredai actually belong to H. beyrichi. Bosquet’s figure 6 depicts the pleon of IRScNB 90-33a, and figure 7 the major claw, IRScNB 90-33c. Figure 71a-b may correspond to IRScNB 90-24b; if not, the palms are very similar. Cephalothoraxes IRScNB 90-24a and 90-33g are referred to H. beyrichi as well.
The cephalothoraxes of H. beyrichi and O. bredai are strikingly similar considering the fundamental differences between the pleons and claws of the two species. There are, however, consistent differences between them. First, the hepatic groove is deeper in O. bredai than in H. beyrichi. Schlüter (1862, p. 722) described, in different terms, that the hepatic groove circumscribed the protuberance “χ” on O. bredai but not on his new species, H. beyrichi. Secondly, the cervical groove on O. bredai more nearly parallels the ventral portion of the branchiocardiac groove than on H. beyrichi. Finally, O. bredai is ornamented with granules, whereas H. beyrichi is covered dorsally and laterally with pits. Schlüter (1862, p. 722) noted that H. beyrichi lacked granules but was ornamented with an extremely fine sculpture.
Range and occurrence. Upper Maastrichtian of Belgium and the Netherlands.
Hoploparia biserialis Fritsch in Fritsch and Kafka, 1887
Figure 11A-C
Partim 1887 Hoploparia biserialis Fritsch in Fritsch and Kafka, p. 35, text-fig. 56, pl. 3, fig. 5; pl. 5, fig. 1 [non pl. 5, fig. 2 = Oncopareia bredai Bosquet, 1854]
 1929 Hoploparia biserialis Fritsch in Fritsch and Kafka; Glaessner, p. 216.
1929 Hoploparia biserialis Fritsch in Fritsch and Kafka; Glaessner, p. 216.
1941 Hoploparia “biserialis” Fritsch in Fritsch and Kafka; Mertin, p. 176, text-fig. 7h.
1941 ? Oncopareia biserialis (Fritsch in Fritsch and Kafka); Mertin, text-fig. 8a.
2010 Hoploparia biserialis Fritsch in Fritsch and Kafka; Schweitzer et al., p. 28.
2010 Oncopareia biseralis [sic] Fritsch in Fritsch and Kafka; Schweitzer et al., p. 31.
2018 Hoploparia biserialis Fritsch in Fritsch and Kafka; Tshudy et al., p. 182.
Material examined. Four specimens in the original Fritsch Collection from the National Museum in Prague, labelled as Hoploparia biserialis, including NM O4044 (Orig. #91; cephalothorax and pleon), NM O3470 (Orig. #86; cephalothorax), NM O3457 (Orig. #87; isolated cutter claw) and NM 06861 (cephalothorax). NM O4044 (Figure 11A) is herein designated lectotype of the species.
Diagnosis. Hoploparia with combined postcervical and branchiocardiac grooves extending to hepatic groove. Antennal carina missing. Cutter claw elongate, with fingers longer than palm.
Remarks. Two specimens, NM O3470 and NM O6861, representing isolated cephalothoraxes of O. bredai were removed from the type collection of H. biserialis (see above). The remaining specimens, NM O4044 and NM O3457, confidently confirm placement in the genus Hoploparia. The long, pointed pleura of H. biserialis NM O4044 (see Fritsch and Kafka, 1887, text-fig. 56; Figure 11A here) are clearly Hoploparia - or Homarus-like and very unlike those of Oncopareia. No claw pairs are known from this specimen. The isolated cutter claw NM O3457 (e.g., Fritsch and Kafka, 1887, pl. 3, fig. 5; Figure 11B-C here) is Hoploparia-like; with a subtriangular palm, the inner margin of which has numerous large, anteriorly directed spines in two rows; fingers are long and robust, lack dentition on occlusal margins and become narrower distally.
It is important to note, that Mertin (1941) considered these two specimens of H. biserialis (sensu Fritsch and Kafka, 1887), NM O3457 and NM O4044, to belong to different taxa. The isolated cutter claw NM O3457 (Mertin, 1941, text-fig. 7h), in Mertin’s (1941) view closely resembling that of H. longimana, was referred to as Hoploparia “biserialis”, whereas the carapace with pleon NM O4044 (Mertin, 1941, text-fig. 8a) was treated as ? Oncopareia biserialis. This pleon with cordate, pointed pleura, and cephalothorax with smooth rostrum did not deter Mertin (1941, p. 181) from placing the species in Oncopareia, which, by his definition, possesses a “stark bedornten” (strongly spiny) rostrum. Although the material comes neither from the same locality nor the same lithostratigraphic unit, we here refer both these specimens to Hoploparia biserialis, following Fritsch and Kafka (1887).
Range and occurrence. Upper Cretaceous, lower–middle Turonian of the Czech Republic; NM O3457 comes from the middle Turonian (lower part of the Jizera Formation) at Bechlín near Roudnice nad Labem and NM O4044 is from the lower–middle Turonian (Bílá Hora Formation) of Bílá Hora (‘Weissenberg’) in Prague.
Hoploparia muncki Pelseneer, 1885
1885 Hoploparia muncki Pelseneer, p. 44 [8], fig. 1.
1929 Hoploparia muncki Pelseneer; Glaessner, p. 221.
1941 Oncopareia muncki (Pelseneer); Mertin, p. 188, text-fig. 8d.
2010 Hoploparia muncki Pelseneer; Schweitzer et al., p. 30.
2010 Oncopareia muncki (Pelseneer); Schweitzer et al., p. 32.
2018 Hoploparia muncki Pelseneer; Tshudy et al., p. 183.
Material examined. None.
Remarks. Tshudy (1993, pp. 147–148) and Tshudy et al. (2018) maintained Pelseneer’s (1885) referral to Hoploparia, based on its antennal carina (albeit short), the diverging submedian carinae of the telson, and the granulation of the cephalothorax, pleon and telson. Its pleonal pleura are more like those of Homarus than those of Hoploparia.
Range and occurrence. Upper Cretaceous, lower Maastrichtian (Ciply-Malogne Phosphatic Chalk Formation; see Robaszynski et al. (2002) of the Mons Basin, southern Belgium).
ASTACIDEA INCERTAE SEDIS
“Hoploparia” macrodactyla Schlüter in von der Marck and Schlüter, 1868
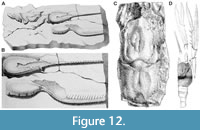
Figure 12A–B
1862 Hoploparia longimana (Sowerby); Schlüter, p. 723, pl. 11, fig. 5.
 1868 Hoploparia macrodactyla Schlüter in von der Marck and Schlüter, p. 295.
1868 Hoploparia macrodactyla Schlüter in von der Marck and Schlüter, p. 295.
1886 Ischnodactylus macrodactyla (Schlüter); Pelseneer, p. 163.
1929 Ischnodactylus macrodactylus (Schlüter); Glaessner, p. 226.
1941 Oncopareia macrodactylus (Schlüter); Mertin, p. 179.
1993 Oncopareia ? macrodactyla (Schlüter); Tshudy, p. 301.
2010 Oncopareia macrodactylus (Schlüter); Schweitzer et al., p. 32.
Material examined. None.
Remarks. Mertin (1941, p. 186) examined Schlüter’s illustrated specimen and noted that: 1) the specimen’s claws were not as well preserved as the illustration indicated; 2) the cephalothorax and shape of the claw palms were indistinguishable from those of Oncopareia coesfeldiensis; and that 3) the claw denticles were not of uniform length, as erroneously depicted in Schlüter, but of different lengths, as in O. coesfeldiensis. According to Mertin (1941), the only difference between Schlüter’s specimen and O. coesfeldiensis was that the palm of the former had two rows of large spines on the upper edge; these spines being absent in O. coesfeldiensis. Mertin (1941) declared the specimen referable to O. coesfeldiensis but did not discuss the specimen’s presumably equal claws, the latter indicated by Schlüter’s illustration and in Pelseneer’s remarks (1886, p. 163). Unfortunately, without the re-examination of the original material (which has not been traced) it is difficult to state whether the morphology of both claws as depicted is genuine. Based on the apparent lack of important characters and poor preservation of the original material we consider “Hoploparia” macrodactyla an astacidean lobster with unknown affinities (Astacidea incertae sedis), at least for the time being.
Range and occurrence. Upper lower ‘Senonian’ of Dülmen, northwest Germany (Mertin, 1941, p. 179); in current terminology this would mean the Dülmener Schichten of early Campanian age.
“Nymphaeops” belgicus Forir, 1887
Figure 12C
1887 Nymphaeops belgicus Forir, p. 157, pl. 7, fig. 1.
1929 Nymphaeops belgicus Forir; Glaessner, p. 279.
2010 Oncopareia belgicus (Forir); Schweitzer et al., p. 31.
Material studied. None.
Remarks. Tshudy (1993, pp. 369-370) concluded that the cephalothorax illustrated in dorsal view in Forir’s (1887) plate was probably not a nephropid. Mertin (1941, p. 188) wrote that the specimen was too poorly preserved to permit any comparison to Oncopareia.
Range and occurrence. Upper Cretaceous, ‘middle Senonian’, probably lower Maastrichtian, of the Liège-Limburg Basin, northwest Belgium and the southeast Netherlands (Forir, 1887, p. 160).
“Nymphaeops” sendenhorstensis Schlüter, 1862
Figure 12D
1862 Nymphaeops sendenhorstensis Schlüter, pp. 732-734, pl. 14, fig. 5.
1886 Nymphaeops sendenhorstensis Schlüter; Pelseneer, p. 173.
1929 Nymphaeops sendenhorstensis Schlüter; Glaessner, p. 279.
1941 Nymphaeops sendenhorstensis Schlüter; Mertin, p. 180.
1993 “Nymphaeops” sendenhorstensis Schlüter; Tshudy, pp. 28, 370-371.
2010 Oncopareia sendenhorstensis (Schlüter); Schweitzer et al., p. 32.
Material studied. None.
Remarks. The generic identity of N. sendenhorstensis is uncertain. Tshudy (1993, pp. 285, 370‒371) concluded that the cephalothorax, pleon (with broken pleura) and heavy, isochelous claws were unlike those of Oncopareia, and treated the specimen as “Nymphaeops” sendenhorstensis.
Range and occurrence. Upper Cretaceous, upper ‘Senonian’ (probably late Campanian) of Sendenhorst, near Münster, western Germany (Mertin, 1941, p. 180) and lower Maastrichtian of the Liège-Limburg Basin, northeast Belgium and the southeast Netherlands (Pelseneer, 1886, p. 173).
“Stenocheles” parvulus Fritsch in Fritsch and Kafka, 1887
Figure 13A-F
partim 1887 Stenocheles parvulus Fritsch in Fritsch and Kafka; p. 40, pl. 3, fig. 3 [non fig. 4 = Ctenocheles fritschi Hyžný, Kočová Veselská and Dvořák, 2014]
 partim 1929 Ischnodactylus parvulus (Fritsch in Fritsch and Kafka); Glaessner, p. 226.
partim 1929 Ischnodactylus parvulus (Fritsch in Fritsch and Kafka); Glaessner, p. 226.
1941 ? Oncopareia parvulus (Fritsch in Fritsch and Kafka); Mertin, p. 185, text-fig. 10g.
1969 ? Oncopareia parvulus (Fritsch in Fritsch and Kafka); Glaessner, p. R459.
2010 Oncopareia parvulus (Fritsch in Fritsch and Kafka); Schweitzer et al., p. 32.
2014 ? Oncopareia parvulus (Fritsch in Fritsch and Kafka), Hyžný et al., p. 246, figs. 1B, 1C, 2B.
Material studied. Lectotype (designated by Hyžný et al., 2014), NM O3455 (part), and NM O9092 (counterpart).
Remarks. Mertin (1941, p. 198) expressed uncertainty regarding the generic placement of this species, but (p. 179) referred it to “? Oncopareia ”. Tshudy (1993) concluded that the two specimens upon which S. parvulus was based (see Fritsch and Kafka, 1887, pl. 3, figs. 3, and 4) represented two different taxa but was unable to refer either specimen to a known genus and assigned both to “Stenocheles”. Hyžný et al. (2014) restudied these two specimens and recognized them as belonging to separate taxa, representing two entirely different infraorders. The heterochelous specimen NM O3456 (Fritsch and Kafka, 1887, pl. 3, fig. 4) became the holotype of Ctenocheles fritschi and the isochelous specimen NM O3455 (part) and NM O9092 (counterpart) (Fritsch and Kafka, 1887, pl. 3, fig. 3) was designated lectotype of S. parvulus (= ? Oncopareia parvulus sensu Hyžný et al., 2014). A new re-examination of the material by one of us (MKV) shows a very poorly preserved specimen with some portions being drawn onto the fossil (Figure 13A). The specimen not only does not correspond to details presented in the illustration provided by Fritsch and Kafka (1887, pl. 3, fig. 3), but also does not preserve sufficient taxonomically important characters to define the species. Moreover, its assignment to Oncopareia or Ctenocheles is highly dubious. The palms of S. parvulus are more elongate than is characteristic of either Ctenocheles or Oncopareia. The finger/palm length ratios on both specimens are smaller than on Ctenocheles, and much smaller than on Oncopareia. Importantly, the genus Stenocheles cannot be used for S. parvulus, because the type species of Stenocheles is Stenocheles esocinus (as designated by Glaessner, 1969), which is here reassigned to Oncopareia, thereby making Stenocheles a junior subjective synonym of Oncopareia (see above). Therefore, we refer S. parvulus to “Stenocheles” and consider “Stenocheles” parvulus a taxon of unknown affinities (incertae sedis). The classification of this dubious taxon at the infraordinal level is here considered only a matter of convenience rather than an assignment based on important characters.
Range and occurrence. Upper Cretaceous, lower–middle Turonian (Bílá Hora Formation) of Bílá Hora (‘Weissenberge’) in Prague, Czech Republic (Fritsch and Kafka 1887, p. 40).
Axiidea de Saint Laurent, 1979
Ctenochelidae Manning and Felder, 1991
Genus Ctenocheles Kishinouye, 1926
Type species. Ctenocheles balssi Kishinouye, 1926, by monotypy.
Ctenocheles cookei (Rathbun, 1935)
Figure 14A–B
1935 Ischnodactylus cookei Rathbun, pp. 63-64, pl. 14, figs. 1‒6.
 1935 Ischnodactylus cookei lowei Rathbun, p. 64.
1935 Ischnodactylus cookei lowei Rathbun, p. 64.
1941 Oncopareia cookei (Rathbun); Mertin, p. 179.
2010 Oncopareia cookei (Rathbun); Schweitzer et al., p. 31.
2010 Ctenocheles cookei (Rathbun); Schweitzer et al., p. 40.
2014 Ctenocheles cookei (Rathbun); Hyžný and Dulai, table 2.
2015 Ctenocheles cookei (Rathbun); Hyžný and Klompmaker, supplement 1.
Material examined. Tshudy (1993, p. 361) examined the holotype (USNM 371511) and identified the two incomplete palms of isolated claws as Ctenocheles cookei lowei.
Remarks. Mertin (1941) synonymized Ischnodactylus with Oncopareia and considered I. cookei to be a species of Oncopareia. On the basis of palm shape and the downflexed fixed finger, Tshudy (1993, pp. 360‒361) classified the isolated claw material of Ischnodactylus cookei as belonging to the axiidean ghost shrimp genus Ctenocheles. Hyžný and Dulai (2014) and Hyžný and Klompmaker (2015) also recognized this form as a representative of Ctenocheles.
Range and occurrence. Lower Eocene (Sucarnoochee beds of the Midway Formation) of Wilcox County, Alabama; and upper Eocene (Jackson Formation) of Yazoo City, Mississippi, USA (Rathbun, 1935).
Ctenocheles cultellus (Rathbun, 1935)
Figure 14C–D
1935 Ischnodactylus cultellus Rathbun, p. 64, pl. 14, figs. 7–12.
1941 Oncopareia cultellus (Rathbun); Mertin, p. 179.
1956 Ischnodactylus cultellus Rathbun; Roberts, p. 6, text-fig. 1.
2010 Oncopareia cultellus (Rathbun); Schweitzer et al., p. 31.
2010 Ctenocheles cultellus (Rathbun); Schweitzer et al., p. 40.
2014 Ctenocheles cultellus (Rathbun); Hyžný and Dulai, table 2.
2015 Ctenocheles cultellus (Rathbun); Hyžný and Klompmaker, supplement 1.
Material examined. The holotype, ANSP 30429.
Remarks. Tshudy (1993) referred the type material of the species to Ctenocheles, an assignment corroborated by Hyžný and Dulai (2014) and Hyžný and Klompmaker (2015).
Range and occurrence. Eocene: several occurrences in Alabama, Mississippi and New Jersey, USA (Rathbun, 1935).
Ctenocheles inaequidens (Pelseneer, 1886)
Figure 14E
1886 Ischnodactylus inaequidens Pelseneer, p. 162 [2], text-fig. 1.
1929 Ischnodactylus inaequidens Pelseneer; Glaessner, p. 226.
1941 Oncopareia inaequidens (Pelseneer); Mertin, p. 179, text-fig. 10o.
1993 Ctenocheles inaequidens (Pelseneer); Tshudy, pp. 361‒362.
2010 Oncopareia inaequidens (Pelseneer); Schweitzer et al., p. 32.
2010 Ctenocheles inaequidens (Pelseneer); Schweitzer et al., p. 40.
2014 Ctenocheles inaequidens (Pelseneer); Hyžný and Dulai, table 2.
2015 Ctenocheles inaequidens (Pelseneer); Hyžný and Klompmaker, supplement 1.
Material examined. Tshudy (1993) examined the holotype (IG 4285, IRScNB 90-36b), a single claw.
Remarks. Tshudy (1993) confidently referred this species to Ctenocheles; this placement has subsequently been corroborated by Hyžný and Dulai (2014) and Hyžný and Klompmaker (2015).
Range and occurrence. Upper Maastrichtian, Kunrade Formation of the Kunrade area, southern Limburg, the Netherlands (Pelseneer, 1886, p. 164).
? Ctenocheles pectiniformis (Böhm, 1891)
Figure 14F
1891 Ischnodactylus pectiniformis Böhm, p. 42, pl. 1, fig. 2, 2a.
1929 Ischnodactylus pectiniformis (Böhm); Glaessner, p. 226.
1941 Oncopareia pectiniforme [sic] (Böhm); Mertin, p. 179, text-fig. 10p.
2010 Oncopareia pectiniformis (Böhm); Schweitzer et al., p. 32.
Material studied. None.
Remarks. Although Tshudy (1993, p. 365) concluded that the generic identity of the specimen, known only by a single claw, was uncertain, the downflexed fixed finger as depicted by Böhm (1891, pl. 1, fig. 2) is very much like those in Ctenocheles. In view of the fact that this species was studied by us only through published figures, we are reluctant to assign I. pectiniformis without any reservations. Nevertheless, assignment to Oncopareia seems to be excluded.
Range and occurrence. Upper Cretaceous, ‘Senonian’ of Bayern, southern Germany.
CTENOCHELIDAE INCERTAE SEDIS
“Ischnodactylus” texanus Rathbun, 1935
1935 Ischnodactylus texanus Rathbun, p. 27, pl. 3, figs. 4-6.
1941 Oncopareia texanus (Rathbun); Mertin, p. 179.
2010 Oncopareia texanus (Rathbun); Schweitzer et al., p. 32.
Material studied. Tshudy (1993, p. 365) examined several non-type specimens, all claw fragments, contained in collections of the Texas Memorial Museum.
Remarks. Tshudy (1993, p. 365) concluded that the studied material very likely did not belong to a nephropid, and that the palm was too rectangular to fit into Ctenocheles. The material as figured by Rathbun (1935) exhibits a morphology that is typical of the minor chelae of Ctenocheles (for details see Hyžný and Dulai, 2014) and major chelae of Gourretia and Dawsonius. Confident generic assignment is pending discovery of better material.
Range and occurrence. Lower Cretaceous (Albian) at several localities in the Duck Creek Limestone, Denton Clay and Paw Paw Clay of northeast Texas (Rathbun, 1935, pp. 27‒28).
BRACHYURA Latreille, 1802
PODOTREMATA INCERTAE SEDIS
“Oncopareia” heterodon Bosquet, 1854
1854 Oncopareia ? heterodon Bosquet, p. 131, pl. 10, fig. 9a, b.
1886 Oncopareia heterodon Bosquet; Pelseneer, p. 173.
1929 Hoploparia ? heterodon (Bosquet); Glaessner, p. 219.
2010 Hoploparia heterodon (Bosquet); Schweitzer et al., p. 30.
2014 Oncopareia heterodon Bosquet; Jagt et al., p. 100, pl. 5, figs. 1, 2.
Material studied. One of us (JWMJ) has recognized several isolated fingers in the Bosquet Collection (IRScNB), embedded in matrix, which might be Bosquet’s originals (see Jagt et al., 2014, pl. 5, figs. 1, 2). There is a lot of additional material, from various levels within the upper Gulpen (Lanaye Member) and lower/middle Maastricht Formation (Valkenburg to Emael members) in the Liège-Limburg Basin (Belgium, the Netherlands). The best-preserved specimen to date is the one illustrated by Jagt et al. (2014, pl. 5, figs. 3, 4).
Remarks. Bosquet (1854) referred this form “avec doute” to Oncopareia. Known mostly by claw fingers, with only a single partial cheliped known to date; these probably belong to a brachyuran, a podotreme crab of uncertain affinities, as indicated by Jagt et al. (2014). We follow their opinion here.
Range and occurrence. Upper Gulpen (Lanaye Member) and lower/middle Maastricht Formation (Valkenburg to Emael members), all upper Maastrichtian, of southern Limburg (the Netherlands) and adjacent Belgian provinces of Liège and Limburg.
DECAPODA INCERTAE SEDIS
“Ischnodactylus” dentatus Rathbun, 1935
Figure 14G
1935 Ischnodactylus (?) dentatus Rathbun, p. 65, pl. 14, figs. 19-22.
1941 Oncopareia dentatus (Rathbun); Mertin, p. 179.
2010 Oncopareia dentatus (Rathbun); Schweitzer et al., p. 31.
2010 Ctenocheles dentatus (Rathbun); Schweitzer et al., p. 40.
2014 Ctenocheles dentatus (Rathbun); Hyžný and Dulai, table 2.
2015 Ctenocheles dentatus (Rathbun); Hyžný and Klompmaker, supplement 1.
Material examined. A single isolated finger, USNM MO495109 (holotype).
Remarks. According to Rathbun (1935, p. 65), each of the fingers studied by her was equipped with “low, triangular, thick” (i.e., not acicular) dentition, unlike that of Oncopareia or Ctenocheles. Although Hyžný and Dulai (2014) and Hyžný and Klompmaker (2015) treated Ischnodactylus dentatus as a representative of Ctenocheles, they did not discuss the species in detail. In the present taxonomic re-evaluation, we propose to remove this taxon from Ctenocheles, based on the lack of acicular dentition, typical of the genus. Unfortunately, the isolated fingers are difficult to assign to any particular group of decapod crustacean. Similar fingers can be present equally in some brachyuran crabs or axiidean ghost shrimp.
Range and occurrence. Middle Eocene of Mississippi, USA (Rathbun, 1935).
CONCLUSIONS
Oncopareia is an extinct clawed lobster genus within the clade of thaumastocheliform nephropid lobsters, characterized by short, quadrate pleura on the pleon and a major claw with a bulbous palm and acicular dentition. Oncopareia is the earliest Thaumastocheles-like lobster with the oldest example being of early to middle Turonian age. Originally, Oncopareia was erected for a single late Maastrichtian species, O. bredai, from the Netherlands. The genus has a convoluted taxonomic history that begins with a type species that is an accidental composite of two lobsters that differ at the generic level. Subsequent to Bosquet’s (1854) erection of a composite type species, the taxonomic confusion surrounding Thaumastocheles-like fossils became even more serious. Workers in the late 1800s erected new genera at a time when access to information and communication among taxonomists were not what they are today. This led to synonymous genera, even for specimens that were anatomically fairly complete. Other scholars in the late 1800s and early 1900s described isolated chelipeds with bulbous palms and slender fingers armed with acicular denticles; most of these were misidentified at the infraordinal level until recently. Mertin (1941) tried to rectify all of this by bringing all fossil lobsters with pectinate claws into one genus; a reasonable approach, but he went one step too far in synonymizing Thaumastocheles-like lobsters with Oncopareia. In fact, most of Mertin’s (1941) synonyms are not thaumastochelids at all, but instead refer to claws of other decapod crustaceans showing convergence, whereas still others are of uncertain affinity. Nonetheless, it became entrenched in the literature that Oncopareia was closely related to Thaumastocheles. Based on the redescription of the type species, Oncopareia bredai, presented herein, we have been able to revise all taxa previously attributed to the respective genus. Besides the type species, three other Oncopareia species are recognized herein. Other taxa, once attributed to Oncopareia, are reassigned to other astacidean or axiidean genera. Taxa based on poorly preserved material are considered of uncertain affinity (incertae sedis).
ACKNOWLEDGMENTS
S. Charbonnier (Muséum National d'histoire Naturelle, Paris) and J. Sklenář (National Museum, Prague) provided access to the material contained at the respective institutions. S.L. Jakobsen (Geomuseum Faxe, Denmark) is acknowledged for taking a new set of photographs of the type material of Oncopareia klintebjergensis. J.W. Stroucken (Natuurhistorisch Museum Maastricht) is thanked for taking photographs of some of the material deposited in NHMM. Constructive criticism of C.E. Schweitzer (Kent State University, Ohio) and an anonymous reviewer greatly improved the original manuscript. The research of MH was supported by Slovak Research and Development Agency (APVV 17-0555). The research of MKV was supported by the Centre for Geosphere Dynamics (UNCE/SCI/006) and by RVO 67985831 of the Institute of Geology of the Czech Academy of Sciences, v. v. i.
REFERENCES
Ahyong, S.T., Chan, T.Y., and Bouchet, P. 2010. Mighty claws: a new genus and species of lobster from the Philippine deep sea (Crustacea, Decapoda, Nephropidae). Zoosystema, 32:525-535. https://doi.org/10.5252/z2010n3a11
Beurlen, K. 1939. Neue Decapoden-Krebse aus dem ungarischen Tertiär. Paläontologische Zeitschrift, 21:135-160. https://doi.org/10.1007/BF03043252
Böhm, J. 1891. Die Kreidebildung des Fürbergs und Sulzbergs bei Siegsdorf in Oberbayern. Palaeontographica, 38:1-106.
Bosquet, J. 1854. Les crustacés fossiles du Terrain Crétacé du Limbourg. Verhandelingen uitgegeven door de Commissie belast met het Vervaardigen eener geologische Beschrijving en Kaart van Nederland, 2:1-127 [10-137]. A.C. Kruseman, Haarlem.
Bosquet, J. 1860. Versteeningen uit het Limburgsche krijt [sic]. In Staring, W.C.H. De bodem van Nederland. De zamenstelling en het ontstaan der gronden in Nederland ten behoeve van het algemeen beschreven: 362-418. A.C. Kruseman, Haarlem.
Bosquet, J. 1868. Liste des fossiles du massif crétacé du Limbourg. In Dewalque, G. : Prodrome d’une description géologique de la Belgique: 355-387. J.-G. Carmanne, Liège.
Bracken-Grissom, H.D., Ahyong, S.T., Wilkinson, R.D., Feldmann, R.M., Schweitzer, C.E., Breinholt, J.W., Bendall, M., Palero, F., Chan, T.-Y., Felder, D.L., Robles, R., Chu, K.-H., Tsang, L.-M., Kim, D., Martin, J.W., and Crandall, K.A. 2014. The emergence of lobsters: phylogenetic relationships, morphological evolution and divergence time comparisons of an ancient group (Decapoda: Achelata, Astacidea, Glypheidea, Polychelida). Systematic Biology, 63:457-479. https://doi.org/10.1093/sysbio/syu008
Bruce, A.J. 1988. Thaumastochelopsis wardi, gen. et sp. nov., a new blind deep-sea lobster from the Coral Sea (Crustacea: Decapoda: Nephropidea). Invertebrate Taxonomy, 2:903-914. https://doi.org/10.1071/IT9880903
Calman, W.T. 1911. The life of Crustacea. Methuen & Co., London.
Chang, S.-C., Tshudy, D., Sorhannus, U., Ahyong, S.T., and Chan, T.-Y. 2017. Evolution of the thaumastocheliform lobsters (Crustacea, Decapoda, Nephropidae). Zoologica Scripta, 46:372-387. https://doi.org/10.1111/zsc.12205
Dana, J.D. 1852. Macrura. Conspectus crustaceorum & conspectus of the Crustacea of the Exploring Expedition under Captain C. Wilkes, U.S.N. Proceedings of the Academy of Natural Sciences of Philadelphia, 6:10-28.
De Grave, S., Pentcheff, N.D., Ahyong, S.T., Chan, T.-Y., Crandall, K.A., Dworschak, P.C., Felder, D.L., Feldmann, R.M., Fransen, C.H.J.M., Goulding, L.Y.D., Lemaitre, R., Low, M.E.Y., Martin, J.W., Ng, P.K. L., Schweitzer, C.E., Tan, S.H., Tshudy, D., and Wetzer, R. 2009. A classification of living and fossil genera of decapod crustaceans: The Raffles Bulletin of Zoology, Suppl. 21:1-109.
de Saint Laurent, M. 1979. Vers une nouvelle classification des Crustacés Décapodes Reptantia. Bulletin de l'Office Nationale de Pêche de Tunisie, 3:15-31.
Feldmann, R.M., Jagt, J.W.M., and Tshudy, D.M. 1990. Late Maastrichtian isopod and decapod Crustacea from Haccourt (Liège), northeastern Belgium. Mededelingen Rijks Geologische Dienst, 44:23-35.
Feldmann, R.M., Schweitzer, C.E., and Karasawa, H. 2016. Part R, revised, Volume 1, Chapter 8J : Systematic descriptions: infraorder Astacidea. Treatise Online, 74:1-28.
Forir, H. 1887. Contributions à l’étude du système crétacé de la Belgique. II. Études complémentaires sur les crustacés: Annales de la Société géologique de Belgique, 14:155-175.
Frič, A. 1893. Studien im Gebiete der Böhmischen Kreideformation. Paläontologische Untersuchung der einzelnen Schichten. V. Priesener Schichten. Archiv der Naturwissenschaftlichen Landesdurchforschung von Böhmen, 9:1-134.
Frič, A. 1894. Studie v oboru křídového útvaru v Čechách. V. Březenské vrstvy. Archiv pro přírodovědné prozkoumání Čech 9. V kommissi knihkupectví Fr. Řivnáče, Prague.
Fritsch, A. and Kafka, J. 1887. Die Crustaceen der böhmischen Kreideformation. Selbstverlag in Commission von F. Rivnáč, Prague.
Glaessner, M.F. 1929. Crustacea Decapoda, Pars 41. In J.F. Pompeckj (ed.), Fossilium Catalogus, 1. Animalia. Junk, Berlin.
Glaessner, M.F. 1969. Decapoda. In Moore, R.C. (ed.), Treatise on Invertebrate Paleontology, Part R, Arthropoda 4(2):R399-R651. Geological Society of America, Boulder, and The University of Kansas Press, Lawrence.
Hyžný, M. and Dulai, A. 2014. Deep-water fossorial shrimps from the Oligocene Kiscell Clay of Hungary: taxonomy and palaeoecology. Acta Palaeontologica Polonica, 59:947-965. https://doi.org/10.4202/app.2012.0078
Hyžný, M. and Klompmaker, A.A. 2015. Systematics, phylogeny, and taphonomy of ghost shrimps (Decapoda): a perspective from the fossil record. Arthropod Systematics & Phylogeny, 73:401-437.
Hyžný, M., Kočová Veselská, M., and Dvořák, P. 2014. On the occurrence of Ctenocheles (Decapoda, Axiidea, Callianassidae) in the Bohemian Cretaceous Basin. Bulletin of Geosciences, 89:245-256. https://doi.org/10.3140/bull.geosci.1421
Jagt, J.W.M. and Bongaerts, H.L. 1986. Kreeftresten uit de Formatie van Vaals (Onder-Campanien, Boven-Krijt) in de groeve Ciments Portland Liégeois, Haccourt, Liège (B.). Natuurhistorisch Maandblad, 75:76-80.
Jagt, J.W.M., Fraaije, R.H.B., and Van Bakel, B.W.M. 2014. Decapod crustacean ‘odds and ends’ from the Maastrichtian type area (southeast Netherlands, northeast Belgium). In Fraaije, R.H.B., Hyžný, M., Jagt, J.W.M., Krobicki, M., and Van Bakel, B.W.M. (eds.), Proceedings of the 5th Symposium on Mesozoic and Cenozoic decapod crustaceans, Krakow [sic], Poland, 2013: A tribute to Pál Mihály Müller. Scripta Geologica, 147:95-115.
Jagt, J.W.M., Fraaije, R.H.B., Van Bakel, B.W.M, and Coole, Y. 2004. New data on the raninid crab Cretacoranina schloenbachi (Schlüter, 1879) (Late Cretaceous, NW Germany). Bulletin of the Mizunami Fossil Museum, 30(for 2003):51-55.
Jakobsen, S.L. and Collins, J.S.H. 1979. Decapod Crustacea from the Paleocene of Zealand, Denmark. Proceedings of the Geologists’ Association, 90:61-64. https://doi.org/10.1016/S0016-7878(79)80033-4
Karasawa, H., Schweitzer, C.E., and Feldmann, R.M. 2013. Phylogeny and systematics of extant and extinct lobsters. Journal of Crustacean Biology, 33:78-123. https://doi.org/10.1163/1937240X-00002111
Kishinouye, K. 1926. Two rare and remarkable forms of macrurous Crustacea from Japan. Annotationes Zoologicae Japonenses, 11:63-70.
Latreille, P.A. 1802. Histoire naturelle, générale et particulière, des Crustacés et des Insectes. Ouvrage faisant suite aux oeuvres de Leclerc de Buffon, et partie du Cours complet d’Histoire naturelle rédigé par C.S. Sonnini, membre de plusieurs sociétés savantes. Familles naturelles des genres, Volume 3. F. DuFart, Paris.
Linnaeus, C. 1758. Systema Naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis (Editio decima) 1. Laurentius Salvius, Holmiae.
Lőrenthey, E. and Beurlen, K. 1929. Die fossilen Dekapoden der Länder der Ungarischen Krone. Geologica Hungarica, Series Palaeontologica, 3:1-421.
Manning, R.B. and Felder, D.L. 1991. Revision of the American Callianassidae (Crustacea: Decapoda: Thalassinidea. Proceedings of the Biological Society of Washington, 104:764-792.
M’Coy, F. 1849. On the classification of some British fossil Crustacea with notices of new forms in the university collection at Cambridge. The Annals and Magazine of Natural History, 4:161-179, 330-335.
Mertin, H. 1941. Decapode Krebse aus dem Subhercynen und Braunschweiger Emscher und Untersenon sowie Bemerkungen über einige verwandte Formen in der Oberkreide. Nova Acta Leopoldina, 10:1-264.
Pelseneer, P. 1885. Notice sur un Crustacé de la Craie Brune des environs de Mons. Bulletin du Musée royal d’Histoire naturelle de Belgique, 4(2):37-46 [1-10].
Pelseneer, P. 1886. Notice sur les Crustacés décapodes du Maestrichtien du Limbourg. Bulletin du Musée royal d’Histoire naturelle de Belgique, 4(2):161-175 [1-15].
Rathbun, M.J. 1926. Phylum Arthropoda, p. 184-191, pls. 63-68. In Wade, B. (ed.), Fauna of the Ripley Formation on Coon Creek, Tennessee. United States Geological Survey Professional Paper, 137.
Rathbun, M.J. 1935. Fossil Crustacea of the Atlantic and Gulf Coastal Plain. Geological Society of America, Special Papers, 2:1-160.
Robaszynski, F., Dhondt, A.V., and Jagt, J.W.M. 2002. Cretaceous lithostratigraphic units (Belgium). In Bultynck, P. and Dejonghe, L. (eds.), Guide to a revised lithostratigraphic scale of Belgium. Geologica Belgica, 4(1-2) (for 2001):121-134.
Roberts, H.B. 1956. Early Tertiary decapod crustaceans from the Vincentown Formation in New Jersey. Bulletin of the Wagner Free Institute of Science, 31:5-12.
Schlüter, C. 1862. Die macruren Decapoden der Senon- und Cenoman-Bildungen Westphalens. Zeitschrift der deutschen geologischen Gesellschaft, 14:702-749.
Schlüter, C. 1879. Neue und weniger gekannte Kreide- und Tertiärkrebse des nördlichen Deutschlands. Zeitschrift der deutschen geologischen Gesellschaft, 31:586-615.
Schweitzer, C.E. and Feldmann, R.M. 2014. Lobster (Decapoda) diversity and evolutionary patterns through time. Journal of Crustacean Biology, 34:820-847. https://doi.org/10.1163/1937240X-00002288
Schweitzer, C.E., Feldmann, R.M., Garassino, A., Karasawa, H., and Schweigert, G. 2010. Systematic list of fossil decapod crustacean species. Crustaceana Monographs, 10:1-222.
Secretan, S. 1964. Les crustacés décapodes du Jurassique supérieur et du Crétacé de Madagascar. Mémoires du Muséum national d'Histoire naturelle Paris, C14:1-226.
Sowerby, G.B. 1826. Description of a new species of Astacus , found in a fossil state at Lyme Regis. Zoological Journal, 2:493-494.
Tshudy, D. 1993. Taxonomy and evolution of the clawed lobster families Chilenophoberidae and Nephropidae. Unpublished PhD dissertation, Kent State University, Kent, Ohio.
Tshudy, D. and Saward, J. 2012. Dinochelus steeplensis , a new species of clawed lobster (Nephropidae) from the London Clay (Eocene) of England. Journal of Crustacean Biology, 32:67-79. https://doi.org/10.1163/193724011X615343
Tshudy, D. and Sorhannus, U. 2000a. Pectinate claws in decapod crustaceans: convergence in four lineages. Journal of Paleontology, 74:474-486. https://doi.org/10.1017/S0022336000031735
Tshudy, D. and Sorhannus, U. 2000b. Jagtia kunradensis , a new genus and species of clawed lobster (Decapoda: Nephropidae) from the Upper Cretaceous (upper Maastrichtian) Maastricht Formation, the Netherlands. Journal of Paleontology, 74:224-229. https://doi.org/10.1017/S0022336000031449
Tshudy, D. and Sorhannus, U. 2003. Hoploparia , the best-known fossil clawed lobster (Family Nephropidae), is a “wastebasket” genus. Journal of Crustacean Biology, 23:700-711. https://doi.org/10.1651/C-2353
Tshudy, D., Hyžný, M., Dulai, A., and Jagt, J.W.M. 2018. Appraisal of the fossil record of Homarus (nephropid lobster), with description of a new species from the upper Oligocene of Hungary and remarks on the status of Hoploparia . Journal of Paleontology, 92:170-182. https://doi.org/10.1017/jpa.2017.65
Van Bakel, B.W.M., Fraaije, R.H.B., and Jagt, J.W.M. 2005. A new Early Campanian species of Xanthosia (Decapoda, Brachyura) from northwestern Germany. Crustaceana, 78:285-295. https://doi.org/10.1163/1568540054286547
von der Marck, W. and Schlüter, C. 1868. Neue Fische und Krebse aus der Kreide von Westphalen. Palaeontographica, 15:269-305.
Wood-Mason, J. 1874. Blind Crustacea. Proceedings of the Asiatic Society of Bengal, 1874:180-181.
